轉錄因子SREBP1通過促進SLC16A8表達調控腫瘤酸性微環境參與結直腸癌發生發展
2023-06-27 08:43:02彭明沙周翼劉剛馮雪雅彭洪
川北醫學院學報 2023年6期
關鍵詞:數據庫
彭明沙,周翼,劉剛,馮雪雅,彭洪
(川北醫學院第二臨床醫學院·南充市中心醫院,1.肛腸外科;2.超聲科,四川 南充 637000)
結直腸癌(colorectal cancer,CRC)是世界范圍內最常見的胃腸道惡性腫瘤。2020年全球新增190萬例CRC病例,有93.5萬例死亡,占所有癌癥死亡的1/10[1]。大多數CRC患者在早期沒有明顯癥狀,大約40%~50%的患者在診斷時已經處于晚期并有遠處轉移[2-3]。因此深入研究CRC的發病機制有利于該病的準確診斷、精準治療和早期預防,也是降低CRC死亡率的有效途徑。SLC16基因家族由14個成員組成,也稱為單羧酸轉運蛋白(monocarboxylate transporter,MCT)家族[4]。SLC16家族成員參與多種代謝途徑,包括能量代謝、糖異生、T淋巴細胞活化、腸道代謝、甲狀腺激素代謝和腦、骨骼肌、心臟和腫瘤細胞中的藥物轉運[5-6]。SLC16A8作為該基因家族成員之一,主要負責單羧酸代謝產物的運輸,如丙酮酸鹽、L-乳酸鹽和酮體[7]。然而尚不清楚SLC16A8與CRC發生發展的關系。甾醇調節元件結合蛋白(SREBPs)是具有基本螺旋-環-螺旋亮氨酸拉鏈結構域的轉錄因子家族,可調節參與脂質合成和攝取途徑的基因[8]。SREBP1由SREBF1基因編碼,是控制細胞脂質代謝的調節器,可通過調控相關基因表達調節膽固醇和脂肪酸的合成及攝入[9]。越來越多的證據表明SREBP1參與了不同癌癥的代謝重編程,如前列腺癌、乳腺癌和膠質母細胞瘤,是癌癥治療的潛在靶點[10]。既往研究[11]發現,肝癌微環境中乳酸濃度的增加可負反饋促進MCT1介導的乳酸攝取并下調SREBP1的表達,然而CRC中SREBP1是否參與SLC16A8介導的乳酸轉運尚無報道。
本研究擬探討SLC16A8通過調控腫瘤酸性微環境促進CRC細胞增殖和侵襲,以及轉錄因子SREBP1上調SLC16A8表達促進乳酸轉運參與CRC發生發展的作用機制。
1 材料與方法
1.1 生物信息學分析
從TCGA數據庫下載619名CRC患者的病例資料,分析SLC16A基因家族14個成員與CRC患者預后的關系。通過UALCAN (http://ualcan.path.uab.edu/analysis-prot.html)網站分析SLC16A8在結腸癌和直腸癌中的表達。通過STRING(https://cn.string-db.org/cgi/input?sessionId=bg39Tea7hbww)數據庫篩選出38個SLC16A8結合蛋白,通過GEPIA2數據庫(http://gepia2.cancer-pku.cn/#index)獲得與SLC16A8基因表達相關的前100個基因,結合這兩個數據集共138個基因。通過Metascape數據庫(https://metascape.org/gp/index.html#/main/step1)對SLC16A8的功能富集進行分析。通過LinkedOmics數據庫(http://www.linkedomics.org/)查詢SLC16A8潛在的轉錄因子,再通過JASPAR(http://jaspar.genereg.net/)網站分析SLC16A8啟動子序列與轉錄因子SREBP1可能的結合位點。通過UALCAN數據庫分析SREBP1 mRNA的表達。使用Kaplan-Meier Plotter網站(http://kmplot.com/analysis/index.php?p=background)分析SREBP1表達水平與CRC預后的關系,再通過GEPIA2數據庫分析SREBF1與SLC16A8表達的相關性。
1.2 細胞培養及轉染
FHC(正常結直腸粘膜上皮細胞)、SW620、SW480、HCT116及HT29細胞株均購自中國科學院細胞庫。FHC和HT29使用1640培養基(Gibco)培養;SW620和SW480使用L-15培養基(Gibco)培養;HCT116細胞使用10%胎牛血清DMEM培養基(HyClone;Cytiva)培養;由GenePharma合成并構建SREBP1過表達載體(OE-SREBP1)、SLC16A8過表達載體(OE-SLC16A8)及SLC16A8 siRNA。根據制造商的說明,使用Lipofectamine 3000(Invitrogen)將其轉染到HCT116細胞中。陰性對照組轉染陰性對照序列到HCT116細胞。SLC16A8 siRNA序列如下:5′-GCCUGUUGUUGCAGAGGAA-3′。通過拯救實驗檢測SREBP1是否通過正調控SLC16A8表達促進乳酸轉運,分組如下:空白對照組,陰性對照組,SREBP1過表達組和SREBP1過表達+SLC16A8 siRNA組。
1.3 RT-qPCR檢測細胞中SLC16A8的表達水平
吸掉培養基,PBS洗1次,吸掉PBS,加入1 mL Trizol(Invitrogen),將底部細胞吹打起來,轉入1.5 mL LEP管裂解細胞,分離各細胞總RNA,在37 ℃下用RNaseR(2 U/μg)處理提取的RNA 10 min,DNA marker以ddH2O為模板作為PCR陰性對照。逆轉錄后,采用2-ΔΔCt法計算基因的相對表達量。SLC16A8序列如下:5- AGCCGTGAGCGTCTTCTTC-3(sense),5- GTCAGGTAGAGCTCCAGGAG-3 (antisense)。
1.4 CCK-8細胞增殖實驗
設立空白對照組(不作處理),陰性對照組(添加無義序列)和SLC16A8過表達組并使用CCK-8檢測細胞增殖。在96孔板中培養HCT116細胞(4×105個/mL),并根據實驗目的轉染48 h后向每孔添加10 μL CCK-8(Dojindo)。2 h后,測試450 nm下的OD值。
1.5 Transwell細胞體外侵襲實驗
設立空白對照組(不作處理),陰性對照組(添加無義序列)和SLC16A8 siRNA組并通過Transwell檢測細胞侵襲能力,使用涂有Matrigel基質的Transwell腔室測試細胞侵襲能力。將處理好的細胞懸浮在無血清培養基中,然后將無血清細胞懸液(500 μL)和添加FBS的培養基(200 μL)分別添加到上腔和下腔。進入下腔的細胞在37 ℃培養48 h后用多聚甲醛固定,然后用0.1%結晶紫染色30 min后在顯微鏡下拍照及計數。
1.6 細胞上清液乳酸測定
設立空白對照組,陰性對照組和SLC16A8過表達組并分析HCT116細胞上清液中乳酸表達水平,將1×104個細胞在含有10%胎牛血清和6 mmol/L葡萄糖的無糖DMEM培養基中培養,然后將細胞轉移至含有5% CO2溫度為37 ℃的培養箱中培養48 h。收集上清液,按照制造商的說明使用熒光乳酸測定試劑盒檢測上清液中乳酸水平。
1.7Westernblot
總蛋白通過RIPA裂解物(Beyotime)收獲,并通過BCA試劑盒(Invitrogen)檢測。在SDS-PAGE凝膠中分離蛋白質(40 μg)后轉移至PVDF膜(Millipore)。封閉后,將膜在4 ℃下暴露于一級抗體(Abcam)過夜,隨后使用二級抗體(Abcam)處理1 h,并通過滴加ECL顯色溶液進行顯影。使用ECL化學發光后,蛋白質在凝膠成像儀(Bio-Rad)上進行顯影。
1.8 雙熒光素酶報告實驗
分別合成熒光素酶報告基因載體pGL3-SLC16A8與熒光素酶報告基因缺失突變載體pGL3-SLC16A8-DM。將pGL3-SLC16A8或pGL3-SLC16A8-DM與pCMV-Myc-SREBP1過表達載體及內參質粒pRL-TK共轉染至HCT116細胞48 h后測定熒光素酶活性。將CRC HCT116細胞接種至24孔板,將熒光素酶報告基因載體pGL3-SLC16A8或熒光素酶報告基因缺失突變載體pGL3-SLC16A8-DM與pCMV-Myc-SREBF1過表達載體或pCMV-Myc質粒及內參質粒pRL-TK共轉染至細胞,設定熒光素酶報告基因載體、pCMV-Myc-SREBF1過表達載體與內參質粒pRL-TK轉染比例為50∶50∶1。轉染48 h后,按照Dual-Luciferase Reporter Assay System(Promega)說明書測定熒光素酶活性。
1.9 統計學分析
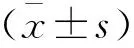
2 結果
2.1 SLC16A家族與CRC預后的關系
TCGA數據結果表明高表達SLC16A8基因的CRC患者提示預后較差(P<0.05),其余13個基因表達水平與CRC患者預后均無相關性(P>0.05)。見圖1。

2.2 SLC16A8在CRC組織中高表達
UALCAN數據庫結果表明SLC16A8在結腸癌和直腸癌中均高于癌旁組織(P<0.05)。見圖2。

2.3 SLC16A8在CRC細胞系中的表達
RT-qPCR結果表明,SLC16A8在CRC細胞株(SW620、SW480、HCT116和HT29)中的表達水平均高于FHC細胞(P<0.05)。既往研究[12]證實SLC16A8在HCT116細胞中穩定表達,因此后續實驗選擇HCT116細胞。見圖3。
2.4 SLC16A8對HCT116細胞增殖的影響
結果表明SLC16A8過表達組中HCT116細胞增殖能力提高(P<0.05)。見圖4。

2.5 SLC16A8對HCT116細胞侵襲的影響
空白對照組(64.00±1.00)和陰性對照組(62.67±1.53)相比,SLC16A8 siRNA組(34.67±1.53)中HCT116細胞侵襲能力降低(t=27.828,22.450,P<0.05)。見圖5。

2.6 SLC16A8功能富集
使用Metascape數據庫對SLC16A8的功能富集進行分析,發現其主要功能聚集在乳酸轉運及血管生成。見圖6。

2.7 SLC16A8對細胞上清液中乳酸水平的影響
結果表明SLC16A8過表達組細胞上清液中乳酸相對表達水平增高(P<0.05)。見圖7。

2.8 SREBP1可能是SLC16A8的轉錄因子
通過LinkedOmics數據庫發現SREBP1可能是SLC16A8的轉錄因子,且SREBP1蛋白與SLC16A8有多個結合位點。因為評分愈高兩者結合的可能性愈大,所以后續實驗選擇評分最高的結合位點“CTCACCCCAT”進行研究。見圖8。

2.9 SREBP1蛋白可與SLC16A8靶向結合
結果顯示與共轉染pCMV-Myc空載體對照組(1.00±0.05)相比,過表達SREBP1可上調SLC16A8熒光素酶報告基因活性(1.79±0.03,t=-24.099,P<0.05)。將SREBP1結合位點“CTCACCCCAT”缺失后,SLC16A8熒光素酶報告基因活性降低(1.19±0.07,t=13.704,P<0.05),說明SREBP1能夠通過結合“CTCACCCCAT”促進SLC16A8轉錄。見圖9。

2.10 SREBF1基因在CRC中高表達
通過UALCAN分析SREBF1在CRC中的表達,結果表明SREBF1在CRC中的表達水平高于癌旁組織(P<0.05)。見圖10。

2.11 SREBF1與患者預后相關
Kaplan-Meier Plotter網站分析結果表明高表達SREBF1的CRC患者預后較差(P<0.05)。見圖11。

2.12 SREBP1正調控SLC16A8表達
GEPIA2數據庫分析結果表明SREBP1與SLC16A8正相關(r=0.24,P<0.05)。接下來分析SREBP1對SLC16A8編碼蛋白MCT3的影響,結果表明與陰性對照組(0.174±0.001)相比,SREBP1過表達組中MCT3蛋白相對表達水平增高(0.592±0.003,t=255.972,P<0.05)。見圖12。

2.13 SREBP1通過正調控SLC16A8表達促進乳酸轉運
拯救實驗檢測SREBP1是否通過正調控SLC16A8表達促進乳酸轉運。結果顯示與空白對照組(1.000±0.032)和陰性對照組(0.979±0.070)相比,SREBP1過表達組細胞上清液中乳酸表達水平增高(1.707±0.144,t=8.290,7.875,P<0.05)。但相比SREBP1過表達組,SREBP1過表達+SLC16A8 siRNA組中乳酸水平降低(1.313±0.057,t=4.403,P<0.05),說明抑制SLC16A8表達可逆轉SREBP1對乳酸轉運的促進作用,證實SREBP1通過正調控SLC16A8促進乳酸轉運。見圖13。

3 討論
CRC是胃腸道常見的惡性腫瘤,其發病率逐年上升。大多數CRC患者確診時已處于晚期,對于這些患者來說手術和輔助治療效果有限且不良反應多,患者預后較差。因此深入了解CRC發生和發展的分子機制至關重要。
SLC16A8屬于SLC16A基因家族,可編碼單羧酸轉運蛋白3(MCT3)。既往研究發現,MCT1-4蛋白主要功能為介導質子耦連的乳酸等單羧酸類物質的跨膜轉運,通過參與調控腫瘤細胞能量代謝和腫瘤微環境酸化影響腫瘤細胞的生物學行為[5-6]。關于SLC16A8(MCT3)的研究較少,研究[13]發現SLC16A8在乳腺癌中低表達,但SLC16A8與CRC發生發展的關系尚無報道。TCGA數據庫的分析結果表明SLC16A8與患者預后相關,高表達SLC16A8的CRC患者預后明顯較差(P<0.05)。與癌旁組織相比,SLC16A8在結腸癌和直腸癌組織及CRC細胞中的表達水平均增高,表明SLC16A8可能與CRC的發生發展密切相關。CCK-8及Transwell實驗結果發現,SLC16A8可促進CRCHCT116細胞的增殖和遷移,證實SLC16A8在CRC發生發展中扮演著促癌因子的角色。為明確SLC16A8發揮促癌作用的具體機制,本研究對SLC16A8的功能富集進行分析并發現其主要功能聚集在乳酸轉運及血管生成。
在實體腫瘤中,由于腫瘤組織迅速增長及腫瘤組織內部血管系統不完備會導致腫瘤組織內氧氣供應不足。由于氧氣供給不足,腫瘤細胞只能夠通過無氧酵解的方式進行能量代謝,其會造成乳酸的積累并反饋抑制糖酵解,同時導致細胞內環境酸化使細胞生長受到抑制;而SLC16A8編碼的MCT3蛋白可特異性的將乳酸轉運到細胞外,這對于調控糖酵解具有重要生理意義[13]。且轉運至細胞外的乳酸可造成腫瘤微環境pH值降低,整體呈現酸性環境,而腫瘤酸性微環境可促進腫瘤細胞的增殖,重塑細胞外基質使腫瘤細胞侵入并轉移到周圍和遠處的器官,還有助于建立利于免疫逃逸的免疫抑制微環境等[14]。本研究發現SLC16A8過表達組HCT116細胞上清液中乳酸表達水平明顯增高,證實SLC16A8可將乳酸轉運到CRC細胞外形成腫瘤酸性微環境,從而促進CRC細胞的增殖和遷移。
轉錄因子是指能夠與真核基因的順式作用元件發生特異性相互作用,并對基因的轉錄有激活或抑制作用的DNA結合蛋白。使用LinkedOmics數據庫對SLC16A8的轉錄因子進行預測,發現其可與SREBP1結合,通過JASPAR網站發現SLC16A8啟動子區與轉錄因子SREBP1有多個結合位點,且熒光素酶實驗也證實SLC16A8可與SREBP1靶向結合。SREBP1蛋白由SREBF1基因編碼,是控制細胞脂質代謝的調節器,但其與腫瘤酸性微環境的關系尚無報道[15]。通過生物信息學數據庫發現SREBF1在CRC中高表達,且高表達SREBF1的CRC患者預后較差,說明SREBF1參與CRC的發生發展。研究結果表明SREBF1基因與SLC16A8基因在CRC中的表達正相關,且高表達SREBP1可促進MCT3蛋白表達,證實轉錄因子SREBP1可通過促進SLC16A8基因轉錄上調MCT3蛋白表達。通過拯救實驗驗證SREBP1通過正調控SLC16A8表達促進腫瘤酸性微環境的形成,結果表明SREBP1過表達組中的乳酸表達水平較空白對照組和陰性對照組增高,但與SREBP1過表達組相比,SREBP1過表達+ SLC16A8 siRNA組中的乳酸水平有所降低,說明抑制SLC16A8表達可逆轉SREBP1對乳酸轉運的促進作用,證實SREBP1通過正調控SLC16A8增加乳酸轉運以形成腫瘤酸性微環境,并最終促進CRC細胞的增殖和遷移。
綜上,本研究結果表明轉錄因子SREBP1可對SLC16A8的轉錄起到正調控作用,導致SLC16A8編碼的MCT3蛋白在結直癌中高表達并促進乳酸外排以形成腫瘤酸性微環境,最終參與CRC的發生發展。SLC16A8在CRC中的研究尚少,本研究為完善CRC的發生發展機制和CRC的防治提供了新的依據和作用靶點,后續還需要更多的體外實驗和體內實驗對其進行驗證。
猜你喜歡
財經(2017年15期)2017-07-03 22:40:49
財經(2017年2期)2017-03-10 14:35:35
華東師范大學學報(自然科學版)(2017年1期)2017-02-27 13:41:08
財經(2016年15期)2016-06-03 07:38:02
財經(2016年3期)2016-03-07 07:44:46
財經(2016年6期)2016-02-24 07:41:51
財經(2015年3期)2015-06-09 17:41:31
財經(2014年21期)2014-08-18 01:50:18
財經(2014年6期)2014-03-12 08:28:19
財經(2013年6期)2013-04-29 17:59:30
