MiR-27a靶向調節PPARγ/SIRT1/NF-κB信號通路對潰瘍性結腸炎幼年小鼠腸道損傷的影響
2023-07-04 11:10:02岳萬勇農國旺潘光澤田文梅張學亮陳光健張飛喻國丹
河北醫藥 2023年10期
關鍵詞:小鼠
岳萬勇 農國旺 潘光澤 田文梅 張學亮 陳光健 張飛 喻國丹
炎癥性腸病(inflammatory bowel disease,IBD)是一種慢性非特異性腸道炎癥性疾病,如潰瘍性結腸炎(ulcerative colitis,UC)或克羅恩病。其中,與成人患者相比,UC兒科患者的疾病范圍、嚴重程度和進展情況明顯更大、更廣[1,2]。盡管目前針對成人UC的外科治療已經得到充分的研究,但并不完全適用于兒童[3]。因此,為了治愈兒童UC,探究其發病機制至關重要。miRN通過與目標mRNA中的互補序列配對并干擾其穩定性和翻譯來調節基因表達,在包括UC在內的多種自身免疫性疾病的發病機制中發揮重要作用[4]。據報道,miR-27a在腫瘤的發生發展中具有重要作用,是一種新型的生物標志物和潛在的腫瘤治療靶點[5]。此外,有研究證實,miR-27a參與UC的發生發展[6]。然而,對于miR-27a在UC中的調控、作用和靶點的理解仍然有限。在本研究中,假設miR-27a在腸道炎癥中具有重要作用,使用2,4,6-三硝基苯磺酸(2,4,6- trinitro picrylsulfonic acid,TNBS)誘導實驗小鼠UC模型[7]。本研究探索miR-27a在TNBS誘導的UC中的作用,并研究miR-27介導的腸道炎癥的潛在機制,為從實驗室到臨床識別和治療UC提供證據。
1 材料與方法
1.1 材料 (1)小鼠:SPF級幼年雄性C57BL/6小鼠(4周齡,16~20g)購自昆明醫科大學,許可證號:SCXK(滇)K2020-0006。本研究符合《實驗動物護理和使用指南》。所有涉及動物的實驗程序均經昆明醫科大學倫理委員會批準。(2)主要試劑:TNBS由Sigma-Aldrich提供,貨號:P2297;蘇木精-伊紅染色試劑盒由武漢賽維爾提供,貨號:G1001-500;TUNEL FITC凋亡檢測試劑盒(綠色熒光)由碧云天提供,貨號:C1098;TRIzol試劑由美國Invitrogen提供,貨號:15596-026;髓過氧化物酶(Myeloperoxidase,MPO)活性檢測試劑盒由Abcam提供,貨號:ab155458;Reverse Transcription試劑盒由vazyme提供,貨號:R323-02;SYBR Green PCR Master Mix由vazyme提供,貨號:Q511-03;腫瘤壞死因子α(tumor necrosis factor-α,TNF-α)、白介素1β(interleukin-1β,IL-1β)、細胞間黏附分子1(Intercellular Adhesion Moleclar-1,ICAM-1)和單核細胞趨化蛋白1(monocyte chemotactic protein-1,MCP-1)ELISA試劑盒由Abcam提供,貨號:ab46105、ab197742、ab252355、ab208979;PPARγ、SIRT1、NF-κB p65(acetyl K310)和β-actin一抗由英國Abcam提供,貨號依次為:ab207297、ab272718、ab189494、ab19870;RIPA裂解緩沖液裂由BOSTER提供,貨號:AR0102-100;BCA蛋白濃度測定試劑盒由博士德生物提供,貨號:AR0197;ECL化學發光檢測試劑盒由新賽美生物提供,貨號:P10300;Dual-Glo Luciferase Assay System由美國Promega提供,貨號:E2920。(3)主要儀器:ELX-800酶標儀由美國BIOTEK提供;DP73光學顯微鏡由日本LLYMPUS提供;ABI 7500 Fasts實時熒光定量PCR系統由杭州博日科技有限公司提供;GelDoc Go凝膠成像系統由上海天能提供。
1.2 方法
1.2.1 TBST誘導UC動物模型的建立及分組處理:小鼠經適應性喂養1周,處理前首先禁食18 h。隨后,將小鼠分為4組(每組10只):假手術對照(Control)組、模型(Model)組、miR-27a抑制劑陰性對照(miR-NC antagomir)組和miR-27a抑制劑(miR-27a antagomir)組。小鼠均使用10%水合氯醛進行腹膜內麻醉,并將由凡士林潤滑的硅膠管(直徑2 mm)通過肛門插入結腸(約8 cm)。除Control組外的其他3組小鼠注射TNBS溶液(3 mg/kg,含50%乙醇)[7],灌腸后,慢慢拔出硅膠管,用棉簽堵住小鼠肛門,將尾部提起,直至自然蘇醒;Control組注射等體積的0.9%氯化鈉溶液。隨后,miR-NC antagomir組和miR-27a antagomir組小鼠分別尾靜脈注射miR-NC antagomir、miR-27a antagomir(200 nmol/d),Control組和Model組尾部靜脈注射等量的0.9%氯化鈉溶液,連續3 d。然后,將各組小鼠10%水合氯醛麻醉處死,收集結腸組織和血液用于進一步的實驗研究。
1.2.2 結腸組織學染色與評分:結腸組織經10%甲醛固定、石蠟包埋后,制備為4 μm切片。將切片在二甲苯中脫蠟,并在梯度濃度的乙醇中水化。使用蘇木精染色5 min后進入1%酸性乙醇(70%乙醇中添加1% HCl)中3 s,后添加伊紅染色3 min。使用梯度乙醇脫水,并在二甲苯中透明。經光學顯微鏡拍照觀察。根據以下標準對結腸組織學損傷進行評分[8]:0分,黏膜、黏膜下層、固有肌層、漿膜結構正常,無炎性細胞;1分,黏膜和黏膜下層有炎性細胞;2分,透壁急性炎癥;3分,小潰瘍,透壁急性炎癥;4分,多處大潰瘍和透壁炎癥;5分,廣泛潰瘍、部分壞死和硬膜內炎癥。
1.2.3 TUNEL染色檢測結腸組織中細胞凋亡:將結腸組織切片在二甲苯中脫蠟,并在梯度濃度的酒精中水化后使用20 μg/ml蛋白酶K室溫下孵育15~30 min,后添加50 μl TUNEL溶液在37℃下避光孵育1 h。使用PBS漂洗后添加DAPI染核,再次漂洗,添加一滴防褪色固化劑孵育5 min,通過熒光顯微鏡觀察并拍照。細胞凋亡率(%)=TUNEL陽性細胞數量/DAPI陽性細胞數量×100%。
1.2.4 結腸組織中MPO活性檢測:首先使用預冷的裂解緩沖液裂解結腸組織,離心20 min(1 000 r/min),之后按照嚴格遵循試劑盒說明書檢測。
1.2.5 ELISA檢測血清中炎性因子含量:將收集到的血液,室溫靜置凝固后離心收集血清,根據ELISA試劑盒方法檢測TNF-α、IL-1β、ICAM-1和MCP-1含量。
1.2.6 qRT-PCR檢測結腸組織中miR-27a表達:使用TRIzol試劑從結腸組織中提取總RNA,并使用反轉錄試劑盒合成cDNA。接著使用SYBR Green PCR Master Mix和實時熒光定量PCR系統進行qRT-PCR測定。U6作為內部對照。通過2-ΔΔCT(ΔCTsample-ΔCTcontrol)來評估miR-27a的相對表達水平。本研究使用的引物序列如下:miR-27a:5’-TGCGCTTCACAGTGGCTAAGT-3’(正向),5’-CCAGTGCAGGGTCCGAGGTATT-3’(反向);U6:5’-CGCTTCGGCAGCACATATAC-3’(正向),5’-AAATATGGAAACGCTTCACGA-3’(反向)。
1.2.7 蛋白質印跡測定結腸組織中PPARγ/SIRT1/NF-κB信號通路相關蛋白表達:使用含PMSF蛋白酶體抑制劑的RIPA裂解緩沖液裂解結腸組織,低溫離心后收集上清液作為完整蛋白質。蛋白質樣品的濃度按照BCA蛋白濃度測定試劑盒說明書測定。每個樣品中20 μg蛋白質通過12% SDS-PAGE電泳分離并轉移到PVDF膜上,使用5%牛血清白蛋白封閉1 h,然后使用TBST洗滌膜。并在4℃下用以下一抗孵育過夜:PPARγ、SIRT1、NF-κB p65和β-actin(內參)。然后將膜與HRP標記的二抗室溫孵育1 h。使用TBST洗滌膜后添加ECL顯影液對蛋白質條帶顯影。使用凝膠成像系統和Image J軟件對蛋白印跡進行分析,計算。目的蛋白相對表達水平=目的蛋白灰度值/內參蛋白的灰度值。
1.2.8 雙熒光素酶報告基因實驗檢測miR-27a和PPARγ靶向關系:Starbase網站(https://starbase.sysu.edu.cn)預測了PPARγ的3’-UTR含有miR-27a的假定結合位點。將PPARγ 3’-UTR的野生型或突變型質粒插入到pmirGLO質粒中,構建WT-PPARγ和MUT-PPARγ質粒。將miR-NC mimics或miR-27a mimics與WT-PPARγ或MUT-PPARγ瞬時共轉染到293T細胞中,轉染48 h后收集細胞,用于熒光素酶報告基因檢測。熒光素酶活性=螢火蟲熒光值/海腎熒光值。
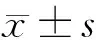
2 結果
2.1 4組小鼠結腸組織學損傷情況 (1)Control組小鼠結腸黏膜光滑完整,無炎性細胞浸潤和潰瘍,上皮細胞排列整齊;Model組和miR-NC antagomir組小鼠結腸黏膜潰瘍和糜爛,大量炎性細胞浸潤,上皮細胞排列混亂且部分損壞、脫落;miR-27a antagomir處理后小鼠結腸黏膜潰瘍和糜爛減輕,炎性細胞浸潤減少,上皮細胞排列較整齊且脫落減少。(2)對結腸組織微觀損傷進行評分,結果顯示:與Control組相比,Model組評分明顯升高(P<0.05);與miR-NC antagomir組相比,miR-27a antagomir組評分明顯降低(P<0.05);miR-NC antagomir組和Model組評分變化比較,差異無統計學意義(P>0.05)。見圖1,表1。
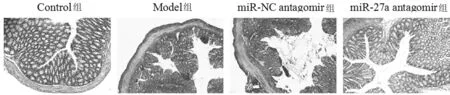
圖1 4組小鼠結腸組織學損傷(蘇木精-伊紅染色×200)
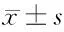
表1 4組小鼠結腸炎評分 n=10,分,
2.2 4組小鼠結腸組織細胞凋亡情況 與Control組相比,Model組小鼠結腸組織中細胞凋亡率明顯增高,差異有統計學意義(P<0.05);與miR-NC antagomir組相比,miR-27a antagomir組小鼠結腸組織中細胞凋亡率明顯降低,差異有統計學意義(P<0.05);miR-NC antagomir組和Model組細胞凋亡率變化比較,差異無統計學意義(P>0.05)。見圖2,表2。
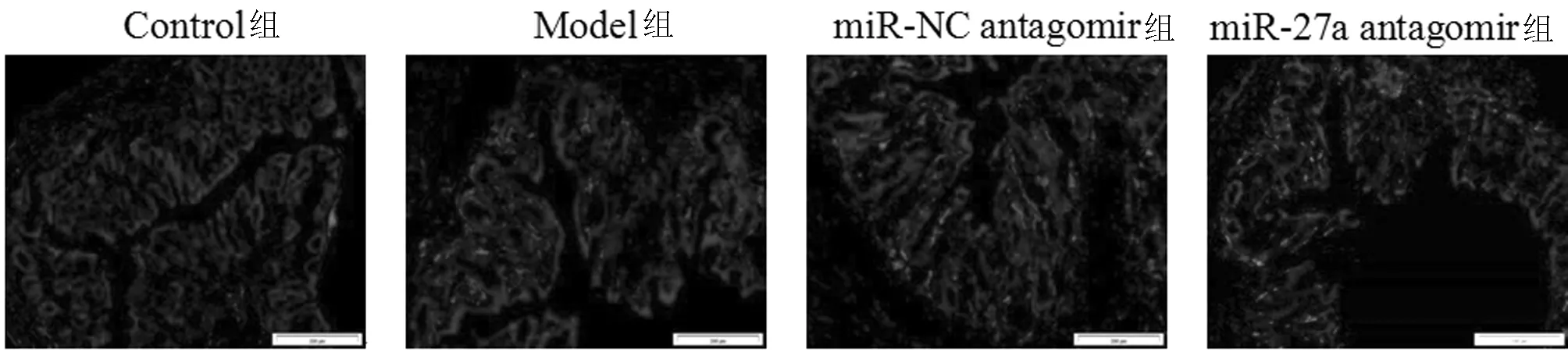
圖2 4組小鼠結腸組織中細胞凋亡情況(TUNEL染色×200)
表2 4組小鼠結腸組織中細胞凋亡率 n=10,%,
2.3 4組小鼠結腸組織中MPO活性 與Control組相比,Model組小鼠結腸組織中MPO活性明顯增高(P<0.05);與miR-NC antagomir組相比,miR-27a antagomir組小鼠結腸組織中MPO活性明顯降低(P<0.05);miR-NC antagomir組和Model組MPO活性變化比較,差異無統計學意義(P>0.05)。見表3。
2.4 4組小鼠血清中炎性因子含量 與Control組相
表3 4組小鼠結腸組織中MPO活性 n=10,pg/mg tissue,
比,Model組小鼠血清中TNF-α、IL-1β、ICAM-1和MCP-1含量明顯增高(P<0.05);與miR-NC antagomir組相比,miR-27a antagomir組小鼠血清中TNF-α、IL-1β、ICAM-1和MCP-1含量明顯降低(P<0.05);miR-NC antagomir組和Model組以上炎性因子含量變化無統計學意義(P>0.05)。見表4。
表4 4組小鼠血清中TNF-α、IL-1β、ICAM-1和MCP-1含量 n=10,pg/ml,
2.5 4組小鼠結腸組織中miR-27a和PPARγ/SIRT1/NF-κB通路相關蛋白表達水平 與Control組相比,Model組小鼠結腸組織中miR-27a表達水平和乙酰化NF-κB p65蛋白水平明顯增高(P<0.05),PPARγ和SIRT1蛋白水平明顯降低(P<0.05);與miR-NC antagomir組相比,miR-27a antagomir組小鼠結腸組織中miR-27a表達水平和乙酰化NF-κB p65蛋白水平顯著降低(P<0.05),PPARγ和SIRT1蛋白水平明顯增高(P<0.05);miR-NC antagomir組和Model組以上指標變化差異無統計學意義(P>0.05)。見圖3,表5。

圖3 Western blot檢測4組小鼠結腸組織中PPARγ、SIRT1和NF-κB p65蛋白水平
表5 4組小鼠結腸組織中miR-27a、PPARγ、SIRT1和NF-κB p65表達水平 n=10,
2.6 雙熒光素酶分析miR-27a與PPARγ靶向關系 經生物信息學分析,PPARγ和miR-27a可能的結合位點如圖所示。應用雙熒光素酶報告基因檢測以確認miR-27a和PPARγ的3’-UTR之間的結合位點,結果表明,與miR-NC mimics組相比較,miR-27a mimics組轉染WT-PPARγ細胞中熒光素酶活性顯著降低,差異有統計學意義(P<0.05),而轉染MUT-PPARγ細胞中熒光素酶活性變化比較差異無統計學意義(P>0.05)。見圖4,表6。
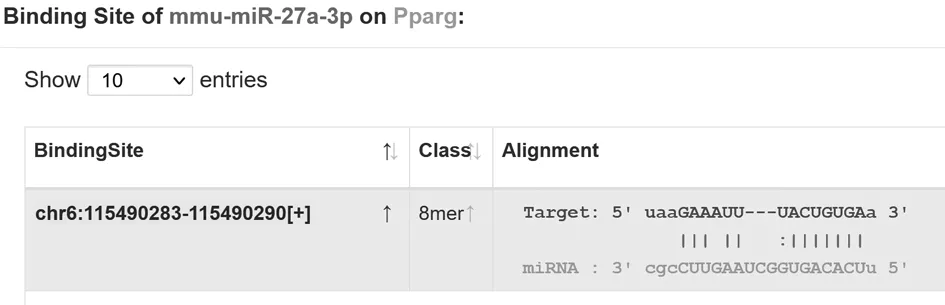
圖4 PPARγ和miR-27a結合位點圖
表6 雙熒光素酶報告基因檢測共轉染293T細胞中的熒光素酶活性n=3,
3 討論
MPO是一種溶酶體蛋白,主要在中性粒細胞中表達,其活性是中性粒細胞浸潤和急性炎癥的重要生物標志物之一[9]。本研究結果表明,TNBS誘導后小鼠結腸炎評分和MPO活性顯著上調,小鼠結腸組織有明顯的潰瘍和糜爛,且大量炎性細胞浸潤,表明TNBS誘導的UC小鼠模型成功建立。同時,本研究結果證明,在TNBS誘導的UC小鼠中,結腸組織中miR-27a的表達被顯著上調。而以往研究顯示,miR-27a在硫酸葡聚糖鈉誘導的小鼠UC模型中顯著下調[10],與本研究結果不同,這可能是因為miR-27a在不同UC模型構建方法或不同周期的小鼠中的作用機制不同。
炎性分子被認為是通過影響腸道通透性和細菌易位參與UC進展的重要因素。TNF-α觸發結腸上皮細胞中其他炎性細胞因子和黏附分子表達;IL-1β通過作用于終末細胞、巨噬細胞和中性粒細胞來調節免疫和非免疫細胞的功能,其水平與UC的疾病嚴重程度呈正相關[11];另外,ICAM-1和MCP-1等炎性細胞因子被視為結腸炎炎癥的生物標志物[12,13]。本研究結果表明,表明miR-27a抑制腸黏膜炎癥。此外,本研究還發現,TNBS誘導后,小鼠結腸組織中細胞凋亡率升高,而抑制miR-27a表達可降低細胞凋亡率。據以往研究發現,適應性的升高細胞凋亡水平可在一定程度上清除堆積的炎性因子[14]。結合本研究結果,推測抑制miR-27a表達減輕了炎性反應,而凋亡反應下調可能是為了適應炎性反應的程度,此結果也間接證實了miR-27a對炎癥的調節作用。
據報道,PPARγ/SIRT1/NF-κB信號通路參與治療硫酸葡聚糖鈉誘導的小鼠UC過程[15]。PPARγ是過氧化物酶體增殖物激活受體的一員,在UC患者結腸上皮中的表達低于對照,且可顯著抑制巨噬細胞的激活和TNF-α、IL-1β在內的多個炎性因子的產生[16]。SIRT1作為長壽蛋白SIR2的哺乳動物同源物,在UC小鼠結腸組織中可通過在賴氨酸310處脫乙酰化RelA/p65來調節細胞功能,從而減少NF-κB活化,從而減少促炎細胞因子的產生[17]。本研究結果顯示,TNBS誘導后,小鼠結腸組織中PPARγ和SIRT1蛋白水平降低,乙酰化NF-κB p65蛋白水平增高,而抑制miR-27a表達可明顯逆轉TNBS誘導對PPARγ、SIRT1、NF-κB表達的影響。另外,生物信息學分析顯示,miR-27a靶向調節PPARγ/SIRT1/NF-κB信號通路參與調控TNBS誘導的UC小鼠腸道損傷過程。
綜上所述,miR-27a靶向調控PPARγ/SIRT1/NF-κB信號通路在UC發病過程中的炎癥調節中發揮重要作用。抑制miR-27a表達可以減輕TNBS誘導的UC小鼠腸道損傷并抑制炎性因子的表達。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
東方少年·布老虎畫刊(2023年8期)2023-08-01 15:45:12
科學大眾(2021年6期)2021-07-20 07:42:44
科學(2020年3期)2020-11-26 08:18:30
學苑創造·A版(2020年9期)2020-10-13 09:41:02
娃娃樂園·綜合智能(2019年3期)2019-04-03 09:17:36
中成藥(2018年2期)2018-05-09 07:19:34
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
湖北師范大學學報(自然科學版)(2015年2期)2016-01-10 08:41:55
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
