桃慢溶質(zhì)性狀的遺傳傾向及初步定位
2023-07-27 11:09:44孟君仁牛良潘磊崔國朝孫世航段文宜曾文芳王志強
果樹學(xué)報 2023年6期
孟君仁 牛良 潘磊 崔國朝 孫世航 段文宜 曾文芳 王志強

摘要:【目的】多數(shù)桃品種果實在成熟后迅速軟化,導(dǎo)致來不及采收、采后易腐爛,是制約桃產(chǎn)業(yè)發(fā)展的關(guān)鍵問題。慢 溶質(zhì)桃具有較長的硬熟期,可以減輕因果實軟化造成的損耗,是未來桃耐貯運育種的重要方向之一。【方法】以慢溶質(zhì) 品種春雪、春瑞為試材,通過構(gòu)建群體、肉質(zhì)評價,對慢溶質(zhì)性狀進(jìn)行遺傳分析和集群分離分析(bulk segregant analysis, BSA)定位。【結(jié)果】通過對比慢溶質(zhì)品種和非慢溶質(zhì)品種成熟過程中軟化的特點,發(fā)現(xiàn)慢溶質(zhì)留樹時間較長,乙烯 延遲釋放。在慢溶質(zhì)分離群體中,留樹時間呈明顯的雙峰分布,遺傳傾向分析表明慢溶質(zhì)受到單基因或主效數(shù)量性狀 基因座(quantitative trait locus,QTL)控制,春雪慢溶質(zhì)位點為雜合,春瑞為純合。以春雪桃自交群體為試驗材料,基于 BSA分析,將慢溶質(zhì)定位到桃第4 號染色體4 個區(qū)段,總長度7.87 Mb。【結(jié)論】桃慢溶質(zhì)性狀遺傳受顯性單基因或主效 基因控制,其控制基因位于第4號染色體。
關(guān)鍵詞:桃;果實;慢溶質(zhì);軟化
中圖分類號:S662.1 文獻(xiàn)標(biāo)志碼:A 文章編號:1009-9980(2023)06-1064-08
桃(Prunus persica)原產(chǎn)中國,自1993 年起中國 桃的栽培面積和產(chǎn)量全面超過意大利和美國,成為 世界第一產(chǎn)桃大國,并一直保持至今[1]。桃因皮薄 肉厚、味道鮮美而廣受消費者喜愛[2]。與柑橘、蘋 果、梨等水果不同,大多數(shù)桃果實成熟后迅速軟化, 留樹時間和貨架期短,若提前采收又會導(dǎo)致果實品 質(zhì)下降,影響消費體驗。此外,隨著桃產(chǎn)業(yè)向集約化 方向的發(fā)展,采后遠(yuǎn)距離運輸成為常態(tài),軟溶質(zhì)桃不耐貯運的短板很大程度上限制了該種肉質(zhì)類型桃的 發(fā)展。
質(zhì)地影響桃果實留樹期和貯運性。目前桃果肉 質(zhì)地性狀主要包括:溶質(zhì)(melting flesh,MF)、不溶 質(zhì)(non- melting flesh,NMF)、硬質(zhì)(stony hard) 等[3]。不溶質(zhì)桃成熟后果實有韌性,留樹時間較長, 傳統(tǒng)上多用于加工罐頭,當(dāng)前也有部分品種進(jìn)入鮮 食市場。硬質(zhì)桃果實成熟后不釋放乙烯,果實保持 硬脆,留樹時間2 周以上,具備優(yōu)異的采后貯運特 性[4],但該種類型果實成熟后不變軟,伴隨著沒有果 實成熟期桃特征性香氣,因此也有一些局限性。此 外,國外最早于2011 年報道了油桃品種Big Top 獨 特的軟化特征,這種肉質(zhì)雖然最終會釋放乙烯使果 實變軟,但果實在成熟前期能保持較高的硬度,具有 較長的留樹期和貨架期[5-6],并將這一質(zhì)地類型分類 為慢溶質(zhì)桃(slow-melting flesh,SMF)。Chen 等[7]對 慢溶質(zhì)單株S11-8 與非慢溶質(zhì)單株M12-10 成熟過 程硬度變化規(guī)律進(jìn)行比較,發(fā)現(xiàn)S11-8 軟化速度比 M12-10 慢得多,而且S11-8 完全軟化的發(fā)生相對較 晚。Ghiani 等[6]通過比較慢溶質(zhì)型桃Big Top 和溶質(zhì) 型桃Bolero 采收后硬度和乙烯的變化,發(fā)現(xiàn)果實成 熟期的Big Top 果實硬度比Bolero 明顯更硬,采后室 溫放置5 d 后果實變軟,最終果實硬度與Bolero 類 似。在果實成熟期采收后,Bolero 桃迅速釋放乙烯, 而Big Top 桃果實采收后4 d 才開始釋放乙烯。對于 慢溶質(zhì)性狀定位的研究也有一些報道,但由于肉質(zhì) 判別標(biāo)準(zhǔn)的缺乏導(dǎo)致基因定位困難,已發(fā)表的定位 結(jié)果表現(xiàn)出較大的差異,相關(guān)分子標(biāo)記仍未能實踐 應(yīng)用[7-9]。
慢溶質(zhì)桃成熟后既有較長的留樹時間和貨架 期,又能夠保持桃果實的特殊風(fēng)味,已成為歐美國家 桃肉質(zhì)改良和發(fā)展的主要方向,但是,慢溶質(zhì)桃在中 國相對較晚才被利用,遺傳資源匱乏,相關(guān)機制研究 甚少。筆者在本研究中以春雪、春瑞等慢溶質(zhì)桃品 種為材料,通過構(gòu)建分離群體,對后代表型進(jìn)行分 析,明確慢溶質(zhì)性狀的遺傳傾向,利用BSA 分析對 該性狀進(jìn)行初步定位。研究結(jié)果初步揭示了桃果實 慢溶質(zhì)肉質(zhì)類型的生理特征、遺傳傾向及初步定位, 為今后慢溶質(zhì)候選基因的挖掘以及調(diào)控機制的研究 奠定基礎(chǔ)。
1 材料和方法
1.1 試驗材料
以SMF品種春雪、春瑞為主要親本構(gòu)建遺傳群 體,具體信息見表1。對雜交單株的肉質(zhì)鑒定于 2021 年5—8 月和2022 年5—8 月在中國農(nóng)業(yè)科學(xué)院 鄭州果樹研究所新鄉(xiāng)試驗基地的雜種苗定植圃內(nèi)進(jìn) 行,株行距為1.0 m×4.0 m,2017 年定植。慢溶質(zhì)桃軟化特點分析所用品種為春麗、春瑞,非慢溶質(zhì)分析 所用品種為春美、春蜜,均來自中國農(nóng)業(yè)科學(xué)院鄭州 果樹研究所桃育種圃。
1.2 試驗方法
1.2.1 自交選擇生長健壯的成年樹作母本樹,于 蕾期對全株進(jìn)行套袋,防止異花授粉,2 周后去除紙 袋。其他管理按常規(guī)。
1.2.2 雜交選擇生長健壯的成年樹作母本,于大 蕾期去雄,點授父本花粉,套袋,10 d 后去袋,其他管 理按常規(guī)。
1.2.3 果實生理指標(biāo)測定果實成熟階段的樣品, 果實達(dá)到八成熟開始采集,每3 d 取樣1 次,每次20 個果實。采樣時間點設(shè)為R、R+3、R+6、R+9、R+12、 R+15,代表八成熟后天數(shù)。果實采收后,立即帶回 實驗室,選取大小均勻、成熟度一致、無病蟲害和機 械損傷的果實,用于軟化生理特點分析。隨機選取9 個果實,在果實赤道處,削去縫合線兩側(cè)及對面部 位的果皮,用水果硬度計GY-4 測定果實去皮硬度, 每個果實的三個部位的平均值作為每個果實的果肉 硬度,單位為牛頓(N)。乙烯測定參考Zeng 等[10]的 方法:隨機選取9 個果實,3 個為一組,置于密封的 2.0 L保鮮罐中,密閉2 h,取1 mL氣體,用氣相色譜 儀GC-2010(島津,日本)測定乙烯釋放量,單位為 nL · g- 1 · h- 1。氣相色譜儀條件為:AI2O3/S 色譜柱 (30 m×0.53 mm,0.25 μm);柱溫50 ℃,SPL1 檢測室 溫度為200 ℃,F(xiàn)ID1 檢測室溫度為200 ℃;載氣氮 氣流速為32 mL·min-1,氫氣流速為40 mL·min-1,空 氣流速為400 mL·min-1。
1.2.4 雜交群體后代果實性狀調(diào)查通過判斷果實 著色完全、果個已膨大到商品果大小、果皮底色已完 成褪綠、品質(zhì)風(fēng)味已可食用,即果實成熟,之后每2 d 調(diào)查1 次果實硬度,參考王力榮等[11]的方法將硬度 劃分為“很硬、硬、中、軟、很軟”5 個等級,統(tǒng)計雜種 單株70%果實在成熟后變軟之前的掛樹時間,大于 等于10 d為慢溶質(zhì),小于10 d 為非慢溶質(zhì)。
1.2.5 BSA 分析在群體后代中分別選取非慢溶 質(zhì)(留樹時間<6 d)或慢溶質(zhì)(留樹時間>14 d)各 20 株,組成2 個極端性狀混池(非慢溶質(zhì)混池為NSMF pool,慢溶質(zhì)混池為SMF pool),提取基因組 DNA,委托北京百邁客生物科技有限公司進(jìn)行重測 序,全部樣品的測序深度為30 倍。參考基因組為桃 Lovell(https://www.rosaceae.org/gb/gbrowse/prunus_ persica_v2.0.a1/)。利用兩混池間基因型存在差異 的SNP、InDel 位點[12],統(tǒng)計差異位點在不同混池中 的深度,并計算每個位點ED(歐氏距離,Euclidean Distance)值。為消除背景噪音,對原始ED 值進(jìn)行 乘方處理,筆者在本試驗中取原始ED的5 次方作為 關(guān)聯(lián)值以達(dá)到消除背景噪音的功能,然后采用DISTANCE 方法[13]對ED值進(jìn)行擬合。取所有位點擬合 值的median+3SD 作為分析的關(guān)聯(lián)閾值,計算得 0.10,設(shè)定為篩選的閾值,閾值外的區(qū)域為潛在候選 調(diào)控基因定位區(qū)域。
1.2.6 數(shù)據(jù)統(tǒng)計分析與作圖數(shù)據(jù)統(tǒng)計分析與作圖 采用微軟Excel 2019。
2 結(jié)果與分析
2.1 慢溶質(zhì)品種成熟過程中硬度變化與乙烯釋放
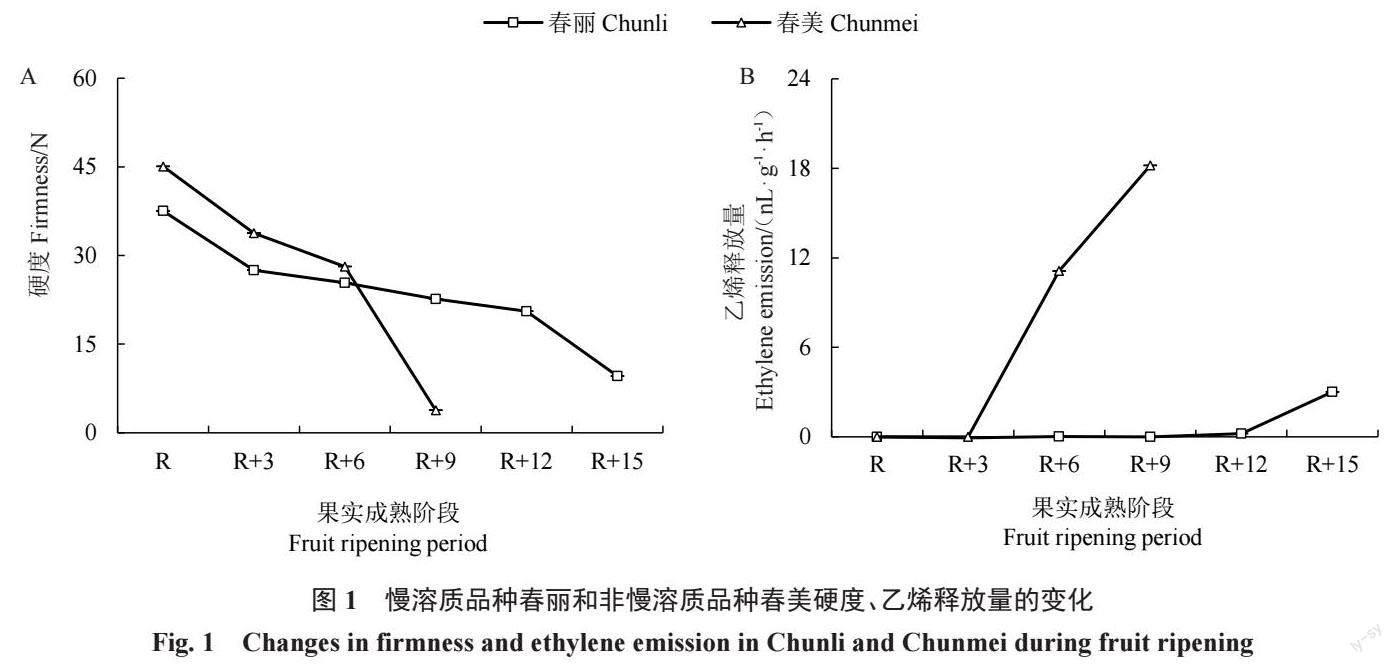
為了顯示慢溶質(zhì)桃成熟過程中的軟化特征,選 擇成熟期相近的兩組慢溶質(zhì)桃vs 非慢溶質(zhì)桃—春 麗vs 春美(圖1)和春瑞vs春蜜(圖2)進(jìn)行比較。結(jié)果 顯示,非慢溶質(zhì)品種春美、春蜜果實硬度在成熟后9 d 已降至最低,慢溶質(zhì)品種春麗、春瑞果實在成熟后15 d 才完全軟化。非慢溶質(zhì)桃春美和春蜜成熟階段硬度 保持在15 N以上的時間僅6 d;慢溶質(zhì)桃春麗和春瑞 至少12 d。因此,將果實成熟后硬度保持在15 N以 上水平的時期定為果實留樹時間,慢溶質(zhì)桃留樹時間 (天數(shù))是非慢溶質(zhì)桃的2 倍左右。慢溶質(zhì)桃品種春 麗在R+12 階段之前乙烯釋放量很低,春瑞在R+12 階段之前沒有乙烯釋放,慢溶質(zhì)桃直到最終軟化發(fā)生 時才伴隨乙烯大量釋放;而非慢溶質(zhì)桃品種在R+6階 段開始就產(chǎn)生很高的乙烯釋放量。綜上兩組數(shù)據(jù),慢 溶質(zhì)桃春瑞、春麗留樹時間至少12 d,相較非慢溶質(zhì)桃春美、春蜜長6 d,乙烯也相應(yīng)延遲釋放。
2.2 慢溶質(zhì)群體后代肉質(zhì)鑒定及遺傳傾向
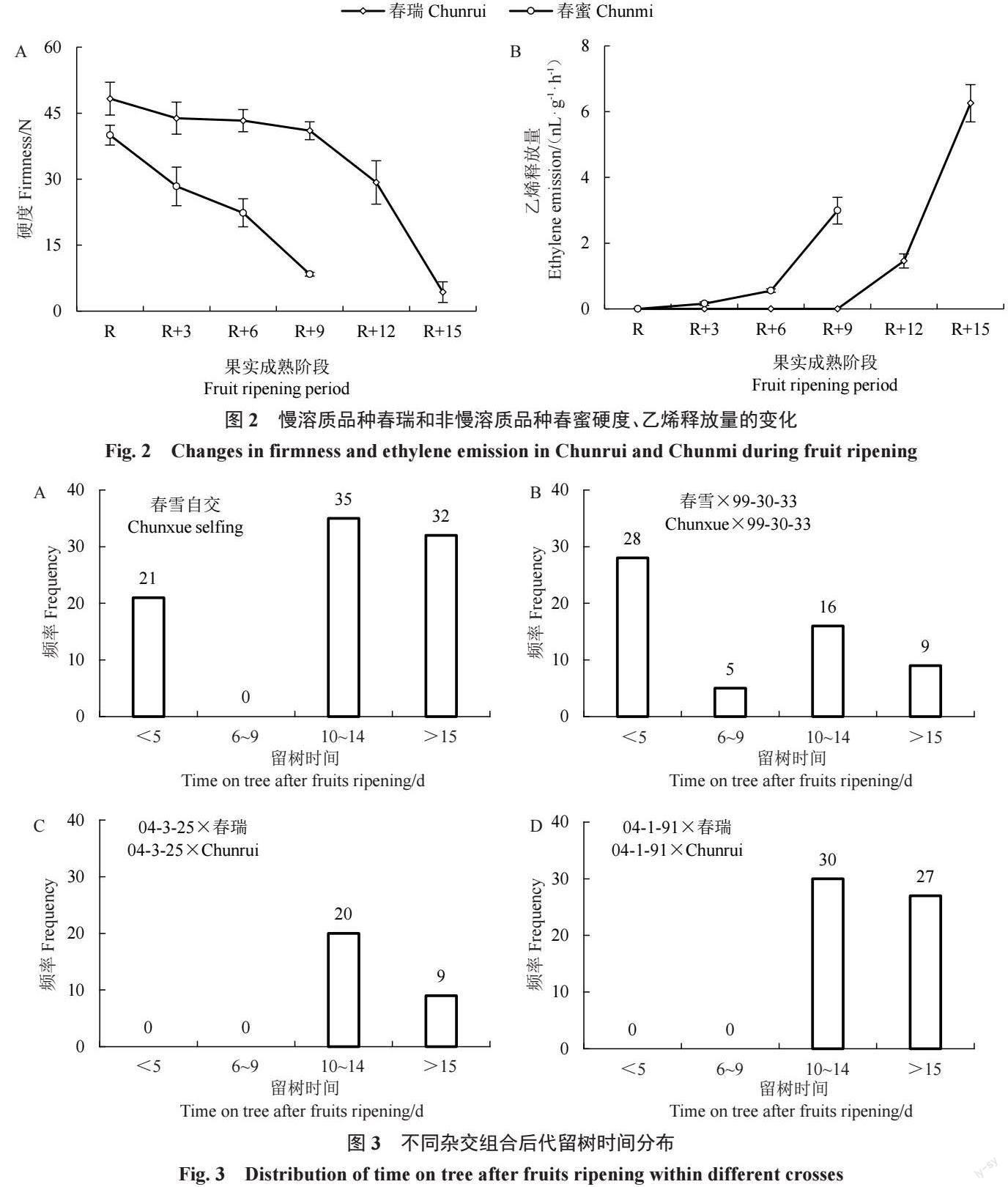
為了更高效快速鑒定群體后代表型,筆者對不 同組合后代果實留樹時間進(jìn)行評價,判別果實肉 質(zhì)。對春雪自交群體進(jìn)行表型評價,發(fā)現(xiàn)后代留樹 時間呈雙峰分布,集中在<5 d 和≥10 d,中間類型 6~9 d幾乎沒有(圖3-A),慢溶質(zhì)桃∶非慢溶質(zhì)桃=67∶ 21。對春雪×99-30-33 群體進(jìn)行表型評價,發(fā)現(xiàn)后代 留樹時間呈雙峰分布,集中在<5 d 和≥10 d,中間類 型6~9 d 只有5 個單株(圖3-B),慢溶質(zhì)桃∶非慢溶質(zhì) 桃=28∶33。對04-1-91×春瑞和04-3-25×春瑞進(jìn)行表 型評價,后代留樹時間只有≥10 d 一種類型,后代沒 有出現(xiàn)<10 d 的類型,群體后代全部為慢溶質(zhì)桃類 型(圖3-C~D)。
春雪組合后代群體中肉質(zhì)分離,春瑞組合后代 中肉質(zhì)不發(fā)生分離。因此,推測親本慢溶質(zhì)桃基因型春雪為雜合,春瑞為純合。春雪自交群體慢溶質(zhì) 桃∶非慢溶質(zhì)桃分離數(shù)據(jù)經(jīng)卡方檢驗符合3∶1 孟德 爾理論分離比例,春雪×99-30-33 群體慢溶質(zhì)桃∶非 慢溶質(zhì)桃分離數(shù)據(jù)符合1∶1 的分離比例(表2),說 明慢溶質(zhì)桃性狀的分離比例與顯性單基因或主效 基因調(diào)控性狀的分離比例一致。
2.3 慢溶質(zhì)性狀初步定位
以春雪為親本自交構(gòu)建的遺傳分離群體為試 材,根據(jù)分離單株肉質(zhì)表型的鑒定結(jié)果,選取極端非 慢溶質(zhì)單株和極端慢溶質(zhì)單株各20 株,構(gòu)建非慢溶 質(zhì)、慢溶質(zhì)基因池,進(jìn)行BSA定位分析。為了鑒定慢 溶質(zhì)桃調(diào)控位點,對兩個混池進(jìn)行高通量特異性片段測序,非慢溶質(zhì)池和慢溶質(zhì)池的測序深度分別為 33 倍和37 倍。在混池之間共獲得79 304 個SNP 和 21 715 個InDel,采用歐氏距離法(Euclidean Distance, ED算法)定位分析方法,將該性狀的調(diào)控位點 關(guān)聯(lián)到了第4號染色體(圖4)。關(guān)聯(lián)分析共得到4個 與慢溶質(zhì)桃性狀相關(guān)的候選區(qū)域,總長度為7.87 Mb (表3)。
3 討論
桃是典型的呼吸躍變型果實,在成熟后期出現(xiàn) 很高的呼吸高峰,果實迅速軟化[14]。筆者對慢溶質(zhì) 桃品種春麗、春瑞成熟軟化的生理特點進(jìn)行分析,留 樹時間長達(dá)12 d 以上,在成熟前期保持較高的硬度, 這與前人[7]對慢溶質(zhì)桃雜交單株S11-8 的生理特征觀察結(jié)果一致。慢溶質(zhì)桃與MF相比,前者果實乙 烯釋放延遲發(fā)生,果實軟化相應(yīng)延遲,這可能是慢溶 質(zhì)桃硬熟期長的原因。除此之外,對慢溶質(zhì)桃遺傳 傾向的分析也與前期春雪×春美雜交后代的研究結(jié) 果一致[15]。此外,Chen等[7]通過對春雪和紅不軟的F1 群體分析,發(fā)現(xiàn)后代慢溶質(zhì)桃∶非慢溶質(zhì)桃接近1∶1, 推測春雪慢溶質(zhì)桃位點也為雜合,可能受顯性單基 因控制。
慢溶質(zhì)性狀的基因定位研究相對匱乏,并且沒 有獲得準(zhǔn)確的定位區(qū)間。Serra 等[8]以慢溶質(zhì)型桃品 種Big Top 為父本,兩種溶質(zhì)型(Armking 和Nectaross) 分別為母本,構(gòu)建雜交群體,通過高密度SNP芯 片構(gòu)建遺傳圖譜,定位了與慢溶質(zhì)桃性狀相關(guān)聯(lián)的 3 個區(qū)段,分別位于第4、5、6 三條染色體上。Ciacciulli 等[9]通過建立力學(xué)模型,對慢溶質(zhì)型BRebus028 和MF型BMax10 雜交后代進(jìn)行表型評價,進(jìn)行連鎖 作圖,將慢溶質(zhì)桃定位到第8 號染色體上的一個 QTL(qSwS8.1),并利用全基因組關(guān)聯(lián)分析(genomewide association study,GWAS)進(jìn)行了驗證。Chen 等[7]通過BSA-seq 鑒定到了第4 號染色體兩個QTL, 并結(jié)合轉(zhuǎn)錄組分析,在5.6 Mb定位區(qū)間內(nèi)鑒定到了 29 個可能的慢溶質(zhì)基因。筆者在本試驗中通過對 春雪自交后代的連續(xù)調(diào)查,結(jié)合遺傳定位分析將慢 溶質(zhì)桃定位在了桃第4 號染色體上,與Serra 等[8]和 Chen 等[7]的定位結(jié)果一致,且有一定的重疊,未來需 要進(jìn)一步研究縮小慢溶質(zhì)桃定位區(qū)間,挖掘慢溶質(zhì) 桃性狀的候選基因。
此外,桃果實成熟期也可能對肉質(zhì)有影響,在對 春雪分離群體的調(diào)查發(fā)現(xiàn),親本春雪含有極早熟的 基因,5 月底之前成熟的極早熟單株中沒有發(fā)現(xiàn)慢 溶質(zhì)桃,之后全部為慢溶質(zhì)桃。由于控制成熟期的 遺傳位點被定位在第4 號染色體[15-17],與慢溶質(zhì)桃在 同一連鎖群上,后代極早熟單株中沒有慢溶質(zhì)型,可 能是慢溶質(zhì)桃與熟期性狀存在不完全連鎖。筆者在 本試驗中進(jìn)行BSA 定位的兩個混池之間除了肉質(zhì) 的顯著差別外,也無法規(guī)避熟期的差異,熟期會影響 對肉質(zhì)的判定。因此,定位結(jié)果中包括了與熟期相 關(guān)的基因,這也為慢溶質(zhì)候選基因的篩選增加了一 定難度。后續(xù)筆者將利用基因組重測序數(shù)據(jù),在定 位區(qū)間開發(fā)更多標(biāo)記,通過更多群體進(jìn)行驗證,更精 細(xì)地對慢溶質(zhì)性狀進(jìn)行定位,為慢溶質(zhì)分子標(biāo)記輔 助育種提供幫助。
4 結(jié)論
通過對慢溶質(zhì)和非慢溶質(zhì)桃成熟軟化的生理差 異進(jìn)行比較,發(fā)現(xiàn)慢溶質(zhì)桃春麗、春瑞成熟期留樹時 間長達(dá)12 d,是非慢溶質(zhì)桃春美、春蜜的2 倍左右, 乙烯延遲釋放,果實最終變軟。遺傳傾向分析表明, 慢溶質(zhì)性狀為顯性單基因或主效基因控制遺傳,且 調(diào)控基因定位在第4號染色體上。
參考文獻(xiàn)References:
[1] 俞明亮,王力榮,王志強,彭福田,張帆,葉正文. 新中國果樹科 學(xué)研究70 年:桃[J]. 果樹學(xué)報,2019,36(10):1283-1291.
YU Mingliang,WANG Lirong,WANG Zhiqiang,PENG Futian, ZHANG Fan,YE Zhengwen. Fruit scientific research in New China in the past 70 years:Peach[J]. Journal of Fruit Science, 2019,36(10):1283-1291.
[2] 徐磊,陳超. 中國桃產(chǎn)業(yè)經(jīng)濟分析與發(fā)展趨勢[J]. 果樹學(xué)報, 2023,40(1):133-143.
XU Lei,CHEN Chao. Economic situation and development countermeasures of Chinese peach industry[J]. Journal of Fruit Science,2023,40(1):133-143.
[3] 曾文芳,王志強,牛良,潘磊,丁義峰,魯振華,崔國朝. 桃果實 肉質(zhì)研究進(jìn)展[J]. 果樹學(xué)報,2017,34(11):1475-1482.
ZENG Wenfang,WANG Zhiqiang,NIU Liang,PAN Lei,DING Yifeng,LU Zhenhua,CUI Guochao. Research process on peach fruit flesh texture[J]. Journal of Fruit Science,2017,34(11): 1475-1482.
[4] 牛良,曾文芳,潘磊,孟君仁,魯振華,崔國朝,王志強. 硬質(zhì)桃 研究現(xiàn)狀及展望[J]. 果樹學(xué)報,2020,37(8):1227-1235.
NIU Liang,ZENG Wenfang,PAN Lei,MENG Junren,LU Zhenhua, CUI Guochao,WANG Zhiqiang. Research status and perspective for stony-hard peach[J]. Journal of Fruit Science,2020, 37(8):1227-1235.
[5] LAYNE D R,BASSI D. The peach:Botany,production and uses[ M]. UK:CABI,2008:1-36.
[6] GHIANI A,NEGRINI N,MORGUTTI S,BALDIN F,NOCITO F F,SPINARDI A,MIGNANI I,BASSI D,COCUCCI M. Melting of‘Big Topnectarine fruit:Some physiological,biochemical, and molecular aspects[J]. Journal of the American Society for Horticultural Science,2011,136(1):61-68.
[7] CHEN C W,GUO J,CAO K,ZHU G R,F(xiàn)ANG W C,WANG X W,LI Y,WU J L,XU Q,WANG L R. Identification of candidate genes associated with slow-melting flesh trait in peach using bulked segregant analysis and RNA-seq[J]. Scientia Horticulturae, 2021,286:110208.
[8] SERRA O,GIN?-BORDONABA J,EDUARDO I,BONANY J, ECHEVERRIA G,LARRIGAUDI?RE C,AR?S P. Genetic analysis of the slow-melting flesh character in peach[J]. Tree Genetics & Genomes,2017,13(4):77.
[9] CIACCIULLI A,CIRILLI M,CHIOZZOTTO R,ATTANASIO G,DA SILVA LINGE C,PACHECO I,ROSSINI L,BASSI D. Linkage and association mapping for the slow softening (SwS) trait in peach [P. persica (L.) Batsch] fruit[J]. Tree Genetics & Genomes,2018,14(6):93.
[10] ZENG W F,PAN L,LIU H,NIU L,LU Z H,CUI G C,WANG Z Q. Characterization of 1-aminocyclopropane-1-carboxylic acid synthase (ACS) genes during nectarine fruit development and ripening[J]. Tree Genetics & Genomes,2015,11(2):18.
[11] 王力榮,朱更瑞,方偉超. 桃(Prunus persica L. )種質(zhì)資源果實 數(shù)量性狀評價指標(biāo)探討[J]. 園藝學(xué)報,2005,32(1):1-5.
WANG Lirong,ZHU Gengrui,F(xiàn)ANG Weichao. The evaluating criteria of some fruit quantitative characters of peach (Prunus persica L.) genetic resources[J]. Acta Horticulturae Sinica, 2005,32(1):1-5.
[12] CINGOLANI P,PLATTS A,WANG L L,COON M,NGUYEN T,WANG L,LAND S J,LU X Y,RUDEN D M. A program for annotating and predicting the effects of single nucleotide polymorphisms, SnpEff[J]. Fly,2012,6(2):80-92.
[13] HILL J T,DEMAREST B L,BISGROVE B W,GORSI B,SU Y C,YOST H J. MMAPPR:Mutation mapping analysis pipeline for pooled RNA-seq[J]. Genome Research,2013,23(4):687-697.
[14] TONUTTI P,CASSON P,RAMINA A. Ethylene biosynthesis during peach fruit development[J]. Journal of the American Society for Horticultural Science,1991,116(2):274-279.
[15] 孟君仁. 桃慢溶質(zhì)性狀調(diào)控機制初步研究[D]. 北京:中國農(nóng)業(yè) 科學(xué)院,2021.
MENG Junren. Preliminary study on regulation mechanism of slow melting flesh characteristics in peach[D]. Beijing:Chinese Academy of Agricultural Sciences,2021.
[16] EDUARDO I,PACHECO I,CHIETERA G,BASSI D,POZZI C,VECCHIETTI A,ROSSINI L. QTL analysis of fruit quality traits in two peach intraspecific populations and importance of maturity date pleiotropic effect[J]. Tree Genetics & Genomes, 2011,7(2):323-335.
[17] PIRONA R,EDUARDO I,PACHECO I,DA SILVA LINGE C, MICULAN M,VERDE I,TARTARINI S,DONDINI L,PEA G,BASSI D,ROSSINI L. Fine mapping and identification of a candidate gene for a major locus controlling maturity date in peach[J]. BMC Plant Biology,2013,13:166.
