不同大熊貓擴散地景觀格局和植被覆蓋變化的比較研究
2023-08-01 04:14:10姜波宇田關勝王翰霖李明明許路冉江洪
四川動物 2023年4期
姜波宇 , 田關勝, 王翰霖 , 李明明 , 許路 , 冉江洪
(1. 四川大學生命科學學院,生物資源與生態環境教育部重點實驗室,成都 610065; 2. 四川大學生命科學學院,四川省瀕危野生動物保護生物學重點實驗室,成都 610065; 3. 四川申果莊自然保護區管理處,四川 越西 616650)
擴散是動物的基本生活史特征(Walters,2000)。動物通過擴散來達到種群內和種群間個體交換的目的,對防止近親繁殖、提高種群環境適應能力、擴大種群的分布區具有重要作用(趙洪峰等,2013;陳儷心等,2019)。目前,擴散已成為生態學家關注的重點問題之一(Walters,2000;Clob?ertet al.,2009)。
動物擴散到新的生境,面臨著繼續遷移或定居的選擇。生境選擇理論通常認為,個體通過做出最優的定居決策,在異質性景觀中選擇最高質量的棲息地,最大化其適合度(Fretwell,1969)。而影響棲息地質量的因子很多,景觀格局和植被覆蓋均是影響棲息地質量的重要因子(徐文茜,2016)。景觀格局是從宏觀尺度反映棲息地質量的主要方法指標(李向陽等,2009;劉艷萍等,2012),很大程度上影響著動物種群的分布(范隆慶等,2010)。此外,植被覆蓋也會影響棲息地質量,一般植被覆蓋越好,區域生態環境質量也越好(王琦等,2019)。同時,植被覆蓋及其動態變化在一定程度上會潛在影響動物棲息地的分布與變化(盧學理等,2008;趙海迪等,2014)。
大熊貓Ailuropoda melanoleuca是世界生物多樣性保護的旗艦物種。目前孤立地分布在我國四川、甘肅、陜西的6 大山系中,其中,涼山山系是大熊貓分布的最南端(和梅香等,2018)。有關大熊貓微生境和宏生境選擇的研究較多(Qiet al.,2009,2011;和梅香等,2018;李楊倩等,2018),但對大熊貓擴散的研究卻鮮有報道,目前僅有通過群體遺傳學數據分析得出大熊貓是偏雌性擴散(Zhanet al.,2007;Huet al.,2010),而對擴散大熊貓的宏生境選擇還未知。
四川省第四次大熊貓調查結果顯示,涼山山系越西縣東北部的梅花鄉-白果鄉區域和雷波縣西部的拉咪鄉-長河鄉區域是2012 年調查發現的大熊貓新分布區,其在2001 年第三次大熊貓調查時均無大熊貓分布(四川省林業廳,2015)。通過對這2 個區域的調查和長期監測,發現截至2021 年越西縣梅花鄉-白果鄉區域一直有大熊貓活動痕跡,而雷波縣拉咪鄉-長河鄉區域在2014—2021 年未發現大熊貓痕跡,這為比較不同大熊貓擴散地景觀格局和植被覆蓋變化,以及大熊貓擴散的宏生境選擇提供了機會。
本文利用3S技術與景觀生態學的數量分析方法定量對比分析了2 個擴散區域的景觀格局和植被覆蓋在近10 年間的時空變化特征,探究景觀格局和植被覆蓋變化對大熊貓宏生境選擇的潛在影響,以期了解可能影響擴散大熊貓宏生境利用的主要因子,識別可能影響大熊貓擴散利用的關鍵環境參數,為大熊貓棲息地的保護與恢復提供科學依據。
1 研究區域概況
研究區域位于四川省涼山山系,根據2012 年全國第四次大熊貓調查對大熊貓棲息地的范圍劃分,以山脊或道路為界(李云飛等,2015),越西區域:越西縣東北部的梅花鄉-白果鄉(102.61°~102.70°E,28.73°~28.87°N),海拔1 517~3 579 m,面積80.22 km2,第四次大熊貓調查發現大熊貓痕跡點18 個,2021 年和2022 年仍然發現有大熊貓活動痕跡;雷波區域:雷波縣南部的拉咪鄉-長河鄉(103.27°~103.42°E,28.17°~28.44°N),海拔1 437~3 471 m,面積173.74 km2,第四次大熊貓調查發現大熊貓痕跡點5個,2014—2021年未發現大熊貓活動痕跡(圖1)。
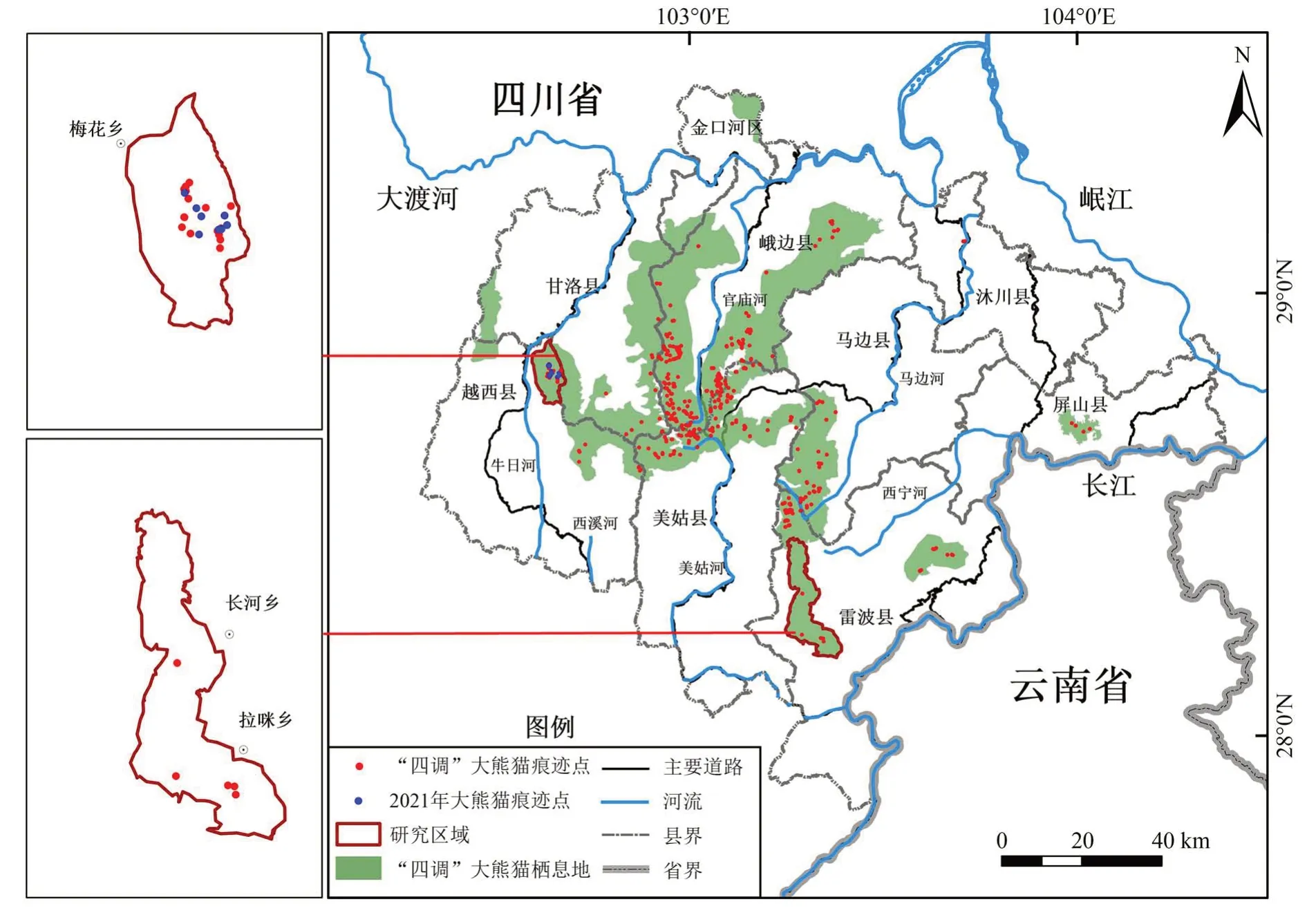
圖1 研究區域Fig. 1 Study area
2 研究方法
2.1 野外調查方法
2.1.1 植被調查 調查樣線按照全國第四次大熊貓調查標準布設,在樣線上合適位置設置植被調查樣方,大小為20 m×20 m,記錄植被類型和地理坐標(饒佳等,2018),用于遙感影像的解譯及之后的精度評價。2021 年6 月在越西區域和雷波區域分別設置了22 條和34 條樣線,70 個和105 個植被樣方。
2.1.2 動物調查 在進行植被調查的同時進行大熊貓痕跡調查,即發現大熊貓痕跡點時,記錄痕跡點類型及其經緯度、海拔等信息。在2021年6月、8月、12月和2022年4月進行了4次調查。
2.2 遙感影像數據獲取
遙感影像數據選擇Landsat8衛星影像,數據來自地理空間數據云(http://www.gscloud.cn)和美國地質勘探局USGS(https://earthexplorer.usgs.gov)。選取研究區域2011 年和2021 年植被生長季且云量低于5%的TM 影像。數據波段空間分辨率均為30 m。
在ENVI5.3 中對2 個研究區域2011年和2021年的Landsat8 遙感影像進行解譯。首先對影片進行預處理,具體過程包括大氣校正、幾何精校正、輻射定標、拼接與裁剪、監督分類與非監督分類相結合等。監督分類主要使用最大似然分類方法,最終將研究區域內土地覆蓋類型劃分為針葉林、闊葉林、灌叢、草甸、農業用地和建筑用地6類。
選擇Cohen 提出的Kappa 系數法(Cantor,1996),并結合實地植被樣方數據和奧維衛星影像圖,由于研究區域面積較小,每一類別分別選取盡可能多的點用于精度評價,以減少由于影像分辨率差異和幾何糾正引起的誤差,若Kappa系數大于0.8,總體精度均達到85%以上,表明解譯精準度較高,解譯結果與真實情況有較強的一致性(Manandharet al.,2009)。2011 年解譯結果通過全國第四次大熊貓調查數據庫中記錄的植被樣方數據進行精度評價,2021 年解譯數據通過2021 年野外調查記錄的樣方數據進行精度評價。
2.3 景觀指數分析
用Fragstats 4.2.1 分別計算研究區域的景觀格局指數,包括斑塊面積、平均斑塊面積、相似鄰近比度、聚集度指數、景觀形狀指數、香農多樣性指數和破碎化指數。采用Mann-WhitneyU檢驗分別對2 個區域各景觀類型2011年和2021年的斑塊面積進行差異分析,統計分析在SPSS 25中進行。
2.4 植被覆蓋分析
歸一化植被指數(normalized differential veg?etation index,NDVI)是反映區域植被動態變化最敏感的指標之一(王小霞等,2020),對植被覆蓋度檢測幅度較寬,具有良好的相位和空間適應性,被廣泛應用于反映區域尺度的植被覆蓋度狀況(蓋永芹等,2009)。
為降低其他干擾因素的影響,選擇植被生長物候期(4—10 月)的數據,以2011 年和2021 年生長期內森林的NDVI 均值代表當年的森林覆蓋度。NDVI 數據來源于美國國家航空航天局NASA(https://ladsweb. modaps. eosdis. nasa. gov/search/)MODIS 數據產品的MOD13Q1 數據集,空間分辨率為250 m×250 m,時間分辨率為16 d。運用MODIS Reprojection Tools 對原始數據進行格式和投影轉換,然后在ArcGIS 10.2 用最大值合成法合成生長期年NDVI數據(Holben,1986)。
根據四川省第四次大熊貓棲息地評價圖層,將適宜棲息地作為高質量棲息地(國家林業和草原局,2021),分別計算2011 年和2021 年研究區域和高質量棲息地的NDVI,柵格大小為250 m×250 m。NDVI年際變化率(K)通過2011年和2021年各像元的NDVI 值計算得到:K=(NDVI2021-NDVI2011)/10。其中,K0.005 表示森林覆蓋度呈增加趨勢(馬曉勇等,2019)。采用Mann-WhitneyU檢驗分別對2個區域及其高質量棲息地各柵格10 年間的NDVI 值進行顯著性差異分析,統計分析在SPSS 25中進行。
3 研究結果
3.1 分類及精度評價結果
分類結果顯示,越西區域2011 年和2021 年的解譯總體精度分別為89.26%和87.59%,Kappa 系數分別為0.823和0.839;雷波區域2011年和2021年的解譯總體精度分別為86.23%和87.77%,Kappa系數分別為0.817和0.847(圖2)。

圖2 2011年和2021年越西區域、雷波區域的景觀格局Fig. 2 Landscape patterns of Yuexi area and Leibo area in 2011 and 2021
3.2 景觀斑塊類型尺度的變化
3.2.1 景觀斑塊類型面積變化 研究區域的主要景觀類型均為森林,包括針葉林和闊葉林,是整個景觀格局的基質;灌叢和草甸面積均不大;農業用地和建筑用地占比很少。10 年間,越西區域針葉林面積減少7.18%,闊葉林面積增加2.97%,森林面積總體減少;雷波區域針葉林面積增加5.72%,闊葉林面積增加5.96%,森林面積總體增加(表1)。
3.2.2 景觀斑塊類型破碎化變化 2011—2021年,越西區域針葉林和闊葉林的平均斑塊面積、相似鄰近比度、聚集度指數均增加,且針葉林的平均斑塊面積增加明顯;森林整體破碎化情況減弱。雷波區域針葉林的平均斑塊面積增加,但相似鄰近比度和聚集度指數均減小;闊葉林的平均斑塊面積減小,相似鄰近比度和聚集度指數也有一定程度減小;森林整體破碎化情況增強。
Mann-WhitneyU檢驗結果表明,2011—2021年,越西區域除草甸和建設用地的平均斑塊面積無顯著變化(P>0.05)外,針葉林極顯著增加(P<0.01),其他景觀斑塊類型顯著增加(P<0.05);雷波區域的闊葉林、灌叢和草甸的平均斑塊面積極顯著減少(P<0.01),針葉林和農業用地顯著增加(P<0.05),建筑用地顯著減少(P<0.05)(表2)。

表2 2011年和2021年研究區域不同景觀類型的平均斑塊面積、相似鄰近比度和聚集度指數Table 2 Mean patch size, percentage of like adjacencies and aggregation index of different landscape types in the two study areas between 2011 and 2021
3.3 景觀尺度的變化
2011—2021 年,越西區域的景觀破碎化指數和景觀形狀指數均減小,香農多樣性指數增加;雷波區域的景觀破碎化指數和景觀形狀指數均增加,香農多樣性指數降低(表3)。

表3 2011年和2021年研究區域不同景觀類型的景觀形狀指數、香農多樣性指數和破碎化指數Table 3 Landscape shape index, Shannon’s diversity index and fragmentation index of different landscape types in the two study areas between 2011 and 2021
3.4 區域整體和高質量棲息地的植被覆蓋度對比
2011—2021 年,2 個區域絕大部分柵格的NDVI 值>0.6,部分柵格的NDVI 值>0.8。10 年間,79.85%越西區域的NDVI值基本保持不變,17.33%增加,高質量棲息地的NDVI 值平均增加0.025;91.31%雷波區域的NDVI 值基本保持不變,7.53%增加,高質量棲息地的NDVI 值平均增加0.019。2011—2021 年,2 個區域高質量棲息地的NDVI 值均顯著增加,但NDVI 值的增加量沒有顯著差異(圖3)。

圖3 2011—2021年越西區域和雷波區域植被覆蓋(NDVI值)的變化Fig. 3 Vegetation coverage (NDVI) changes in Yuexi area and Leibo area from 2011 to 2021
4 討論
本研究結果顯示,2 個區域森林面積的變化幅度都相對較小。2011—2021 年,越西區域針葉林面積有一定減少,主要是由于針葉樹種被當地社區居民作為木材和薪材采伐所致;其他景觀類型的轉換主要是由放牧、耕地開墾、新農村基礎設施建設等活動所致。雷波區域的變化主要源自植樹造林導致的灌叢和草地面積減少,森林面積增加,新農村和基礎設施建設導致了建設用地增加。
2011—2021 年,雷波區域整體森林破碎化程度增加,主要是由于人工新造林及植被自然恢復更替等導致針葉林和闊葉林斑塊鑲嵌較多;而2001 年前森林采伐和農墾活動頻繁,致使林相退化,以次生林為主(戴波等,2014)。而次生林是由多個先鋒樹種形成不同的斑塊,導致區域異質性增加,形成的斑塊面積也較小,森林群落的演替會使區域破碎化程度加劇(張會儒等,2009)。越西區域則主要是部分針葉林轉化為闊葉林,導致區域內針葉林、闊葉林斑塊分布更規則、聚集,整體森林破碎化程度降低。
2011—2021 年,越西區域景觀受人類活動干擾程度減少,景觀整體破碎化程度減弱;而雷波區域受人類活動干擾程度增加,景觀整體破碎化程度增加。對于以森林為主要景觀類型的區域,植被的次生演替可以使景觀結構趨于復雜,景觀豐富度提高,多樣性指數提高;而諸如采伐、開荒等人類活動往往使景觀類型變得單一、豐富度下降、多樣性指數降低(張爽等,2004)。越西區域植被次生演替情況更好,植被恢復情況更好,而雷波區域植被次生演替情況較差,且人類活動更頻繁,這與2 個區域景觀形狀指數得出的結論一致。產生這種結果的原因可能是,雷波區域村莊較多且分散,居民的農牧活動對整體景觀影響較大;且過去10 年間人工林種植較多,種植類型較為單一,也使其植被次生演替情況較差,多樣性指數較低。
2 個區域在2011 年和2021 年均為植被覆蓋良好區域(NDVI>0.6)(王曉利等,2013)。2011—2021年,2 個區域中只有部分區域的NDVI 增加,而大部分區域維持不變。越西區域的增加部分大多屬于天然林,減少區域主要位于居民區附近,由開墾農田、放牧、林木砍伐等所致。雷波縣的增加區域主要位于人工造林區域,減少區域主要是居民點周邊及草地區域,由林木砍伐及放牧等活動所致。
棲息地破碎化和植被覆蓋變化是宏觀尺度上對動物棲息地選擇的兩大重要影響因素(楊少文,2015)。對于大熊貓這樣的大型哺乳動物,棲息地破碎化和植被覆蓋變化影響其分布的現象尤為明顯(白文科等,2017;朱若檸等,2019)。森林和整體景觀破碎化使大熊貓的活動范圍減小,生存環境孤立,影響大熊貓的擴散行為(李國春,2017;陳文德等,2021)。本研究發現,擴散大熊貓對宏生境的選擇也與森林和景觀破碎化有關,偏向于選擇森林(尤其是針葉林)和整體景觀破碎化較低的生境,這與張爽等(2004)的研究結果類似。而植被覆蓋度與大熊貓的隱蔽選擇、棲息地選擇緊密聯系,大熊貓對隱蔽條件的要求可理解為對植被(尤其是喬木)覆蓋度的選擇及對各種干擾的趨避(孫亮等,2022),其選擇具有較高喬木郁閉度的發育成熟的濃密林冠下的棲息地(Wanget al.,2022),較大的喬木郁閉度可為其提供良好的隱蔽條件(孫飛翔等,2013)。但同時,林下竹子的生長也需要一定的光照條件,較低的郁閉度和喬木密度(植被覆蓋度)可以滿足林下竹子生長對光照的需求(李爽等,2017)。本研究并沒有發現植被覆蓋的提升對大熊貓的宏生境選擇產生積極影響,可能是因為在2 個區域植被覆蓋度自身基數較高的情況下,植被覆蓋度的增加并不會使生境的適合度產生明顯的提高。
在宏觀尺度上,大熊貓擴散后居留的越西區域相較其短期停留后又離開的雷波區域,森林和整體景觀破碎化程度均相對較低,景觀受人為活動影響較小,大熊貓主要活動的針葉林平均斑塊面積較大,同時區域的香農多樣性指數也較高。這可能預示著大熊貓對新擴散區域的森林和整體景觀破碎化、針葉林的平均斑塊面積具有一定的選擇性,同時可能偏好森林管護好、人為活動影響小、植被快速演替恢復的區域。這提示在大熊貓棲息地保護與恢復時,需要注重棲息地景觀結構的恢復,減少森林景觀破碎化程度,擴大其主要活動棲息地的斑塊面積,同時加強管控措施減少干擾,促進植被的健康恢復。
影響大熊貓宏生境選擇的主要因素包括地理環境、森林群落結構、主食竹和干擾等(何敏等,2018),本研究僅涉及到影響大熊貓擴散的地理環境因素和森林群落結構因素中的一部分。影響大熊貓擴散居留的還有其生物學特征,如繁殖等生理因素(魏輔文,2018),并不完全由外界條件決定。因此,后續研究還需要結合更多類型數據,從多個方面進行綜合分析,以進一步了解擴散大熊貓對宏生境的選擇,科學系統地指導大熊貓的保護與棲息地恢復工作。本結果可為認識擴散大熊貓的宏生境選擇提供有益的參考,對大熊貓棲息地恢復具有積極作用。
致謝:感謝四川省林業和草原局提供的四川省第四次大熊貓調查資料;感謝越西縣林業和草原局、雷波縣林業和草原局和越西申果莊自然保護區、雷波嘛咪澤自然保護區等單位與相關人員給予的幫助。感謝李雨航師兄、劉雨欣師姐、夏林超等人對數據收集與分析、制圖、論文修改的幫助。
猜你喜歡
現代裝飾(2021年6期)2021-12-31 05:27:54
今日農業(2021年9期)2021-11-26 07:41:24
發明與創新·小學生(2021年3期)2021-03-25 11:48:49
小學科學(學生版)(2020年12期)2021-01-08 09:28:10
小學科學(學生版)(2020年7期)2020-07-28 08:00:52
少年漫畫(藝術創想)(2020年12期)2020-06-09 05:50:08
照明工程學報(2016年3期)2016-06-01 12:17:56
中國科技博覽(2016年2期)2016-04-25 20:32:39
小學生導刊(2016年34期)2016-04-11 00:49:44
電測與儀表(2015年5期)2015-04-09 11:30:52
