兩種池塘養殖模式下異育銀鯽(Carassius auratus gibelio)養成品質及臟器相關生理特征的差異*
2023-08-03 07:32:18李有志戎華南申屠基康王志錚
海洋與湖沼 2023年4期
李有志 戎華南, 2 陳 雨 申屠基康 王志錚
兩種池塘養殖模式下異育銀鯽()養成品質及臟器相關生理特征的差異*
李有志1戎華南1, 2陳 雨1申屠基康3王志錚1①
(1. 浙江海洋大學水產學院 浙江舟山 316022; 2. 慈溪市水產技術推廣中心 浙江慈溪 315300; 3. 寧波市海洋與漁業研究院 浙江寧波 315048)
水產養殖動物的生存策略往往會因養殖模式的變更而發生改變, 并導致其養成品體色、肌肉品質和臟器生理特征等均出現一定程度的差異。以池塘生態主養模式(M1)和池塘生態套養模式(M2)下養殖7個月的異育銀鯽夏花苗種為研究對象, 以體表色差、背肌物性、全魚營養成分和排氨率、耗氧率、窒息點以及臟器消化酶和抗氧化酶活力為測定指標, 較系統開展了兩者間養成品質及臟器相關生理特征的差異研究。結果表明: (1) 兩者間在體表色差上具較好的區分度, 其中M2實驗個體的體色更接近于池底背景色, 具更好的擬境隱蔽性; (2) 在所測7項背肌物性指標中, 兩者間具有顯著差異的為硬度、彈性、膠黏性和耐咀性, 且均呈M1>M2(<0.05); (3) 全魚一般營養成分中, 除灰分和粗蛋白含量均呈M1≈M2(>0.05)外, 水分和粗脂肪含量分別呈M1>M2(<0.05)和M2>M1(<0.05); (4) 在所測17種氨基酸中, 在含量上具顯著差異的僅為組氨酸, 呈M1>M2(<0.05), 在各類氨基酸含量上具顯著差異的僅為∑SEAA、∑NEAA和∑FAA, 且均呈M1>M2(<0.05); (5) 共檢到4種飽和脂肪酸、5種單不飽和脂肪酸和10種多不飽和脂肪酸, 其中ΣSFA和ΣMUFA均呈M1
養殖模式; 異育銀鯽; 體表色差; 背肌質構; 營養成分組成; 生理特征
體色和食用價值既是評價水產養殖動物養殖品質的重要內容, 也是反映其生存福利狀況的具體表現。消化、呼吸、排泄和抗氧化生理作為水產動物機體代謝互為關聯的方面, 均與其所處生存環境條件息息相關(黃溢明, 1982; 劉松巖, 2006)。Wagner等(2001)指出, 同種生物的不同群體為了經常適應不同的生活環境而形成了各自特有的生物學特性, 包括生長、發育、繁殖以及對環境因子的適應力等。王志錚等(2012, 2013a, 2013b)的研究也表明, 水產養殖動物的生存策略會因養殖模式的變更而發生改變, 并導致其養成品形質、體色、肌肉品質、血清生化和臟器消化酶與抗氧化酶活力均出現一定程度的差異。因此, 比較不同養殖模式下目標水產養殖動物機體生理代謝水平的差異, 進而深入全面探究其體色和食用價值由此發生改變的邏輯關聯, 對于優化目標水產養殖動物的養殖模式與工藝參數, 進而推進其健康高質養殖具有重要現實意義。
異育銀鯽系我國七大大宗淡水魚類之一(戈賢平, 2010), 廣泛養殖于我國沿海和內陸省份, 池塘主養和池塘套養為其最為常見的傳統養殖模式(鄭勁松, 2004; 楊興麗等, 2022)。為探析不同養殖模式下異育銀鯽的生存對策差異, 本研究團隊以生態主養模式(M1)和生態套養模式(M2)下養殖7月齡的異育銀鯽夏花魚苗為研究對象, 從形質特征和生物學性狀對體質量影響效果兩個維度, 揭示了M1和M2實驗群體為分別貫徹并實施積極取食和伏擊取食策略, 依次采取的運動和攝食并重的高能耗“添加模式”(additivity model)和更專注攝食的低能耗“優先模式” (prioritization model), 是導致兩者在增重機制和-生存對策選擇軸上均發生明顯乖離的本質原因(陳雨等, 2022; 徐英杰等, 2023)。為進一步聚焦M1和M2實驗群體間的生存對策差異, 并印證上述研究結果的可靠性, 本研究于2020年12月中旬較系統開展了M1和M2實驗群體間體表色差、背肌質構、全魚營養成分以及臟器消化、呼吸、排泄、抗氧化生理的差異研究, 以期為銀鯽品質鑒定系統的構建以及生態高值養成技術研究與開發提供基礎資料。
1 材料與方法
1.1 實驗材料
1.1.1 供試鯽 實驗用異育銀鯽()的來源及選取要求均完全同陳雨等(2022)。
1.1.2 實驗用水 為經自然曝氣48 h的自來水, pH (7.56±0.02)、DO (7.49±0.05) mg/L, 水質符合《NY 5051-2001無公害食品淡水養殖用水水質》要求。
1.2 實驗方法
1.2.1 體表色差的測定 隨機選取M1和M2實驗個體各3尾, 以鰓蓋部、尾柄部、體背部和體腹部的中央區域以及側線部(按等間距法沿側線走向設置5個測定位點, 取其均值)為測定部位, 采用CR-400色差儀(柯尼卡美能達控股株式會社), 逐尾測定體表Hunter LAB值(值由黑至白的取值范圍為0~100,值由綠至紅的取值范圍為–60~60,值由黃到藍的取值范圍為–60~60)。
1.2.2 背肌物性的測定 任取M1和M2實驗個體各5尾, 逐尾刮除背部鱗片并用手術刀和手術剪割取背肌組織塊制成規格為2 cm×2 cm×1 cm的鮮樣后, 即刻用TA.XT Plus型食品物性測試儀測定硬度、黏性、彈性、內聚性、膠黏性、耐咀性和回復性等7項物性指標。物性測試儀采用TPA模式, 測試探頭為P/5, 測試前、后移動速度均為5 mm/s, 測試移動速度為1 mm/s, 測距為3 mm。
1.2.3 營養成分的測定 隨機選取停食暫養2 d后的M1和M2實驗個體各30尾進行全魚營養成分測定。其中, 水分、灰分、粗脂肪、粗蛋白含量以及脂肪酸組成的樣品處理及檢測方法分別按GB 5009.3-2016 (直接干燥法)、GB 5009.4-2016、GB 5009.6-2016、GB 5009.5-2016和GB/T 5009.124-2003 (酸堿水解法),氨基酸組成采用日立L-8900高速氨基酸分析儀進行測定。
1.2.4 耗氧率、排氨率和窒息點的測定 在室溫26℃條件下, 以容量為10 L為透明塑料水桶(實驗實際容積為5 L)為呼吸室, 各呼吸室均放入同一養殖模式實驗魚4尾, 按楊程等(2016)的方法分別測定M1和M2實驗個體的耗氧率、排氨率和窒息點。其中, M1和M2實驗個體的排氨率和耗氧率測定均各設3個重復, 窒息點測定均各設4個重復。
1.2.5 臟器消化酶和抗氧化酶活力的測定 任取M1和M2實驗魚各3尾, 于冰盤上逐尾解剖摘取心、肝、胃、腸和鰓, 并去除其內容物及附于其上的脂肪和結締組織, 4 °C雙蒸水沖凈, 濾紙吸干表面水分后, 將各臟器分別放入規格為5 mL的離心管內并作好標記保存于–80 °C超低溫冰箱備測。其中, 消化酶測定指標為胃蛋白酶、腸淀粉酶和肝脂肪酶, 測定抗氧化酶的靶器官為心、鰓、肝, 測定指標為SOD (超氧化物歧化酶)、CAT (過氧化氫酶)和POD (過氧化物酶)。測定上述酶活的試劑盒均購自南京建成生物工程研究所, 測定步驟及計算方法均按所附說明書。
1.3 數據處理
借助SPSS17.0對實驗所得各項數據進行統計分析, 并比較組內、組間差異顯著性(<0.05為差異顯著)。
2 結果
2.1 體表色差
由表1可見, M1和M2實驗個體間在體表色差上的異同主要表現為: (1)值除鰓蓋部中央區和側線部均呈M1>M2(<0.05)外, 其余測定部位均呈M1≈M2(>0.05), 即M1實驗個體的側線部和鰓蓋部中央區均較M2更顯亮白; (2)值僅鰓蓋部中央區呈M1>M2(<0.05), 其余測定部位均呈M1
表1 兩種池塘養殖模式下實驗鯽的體表色澤Hunter LAB值
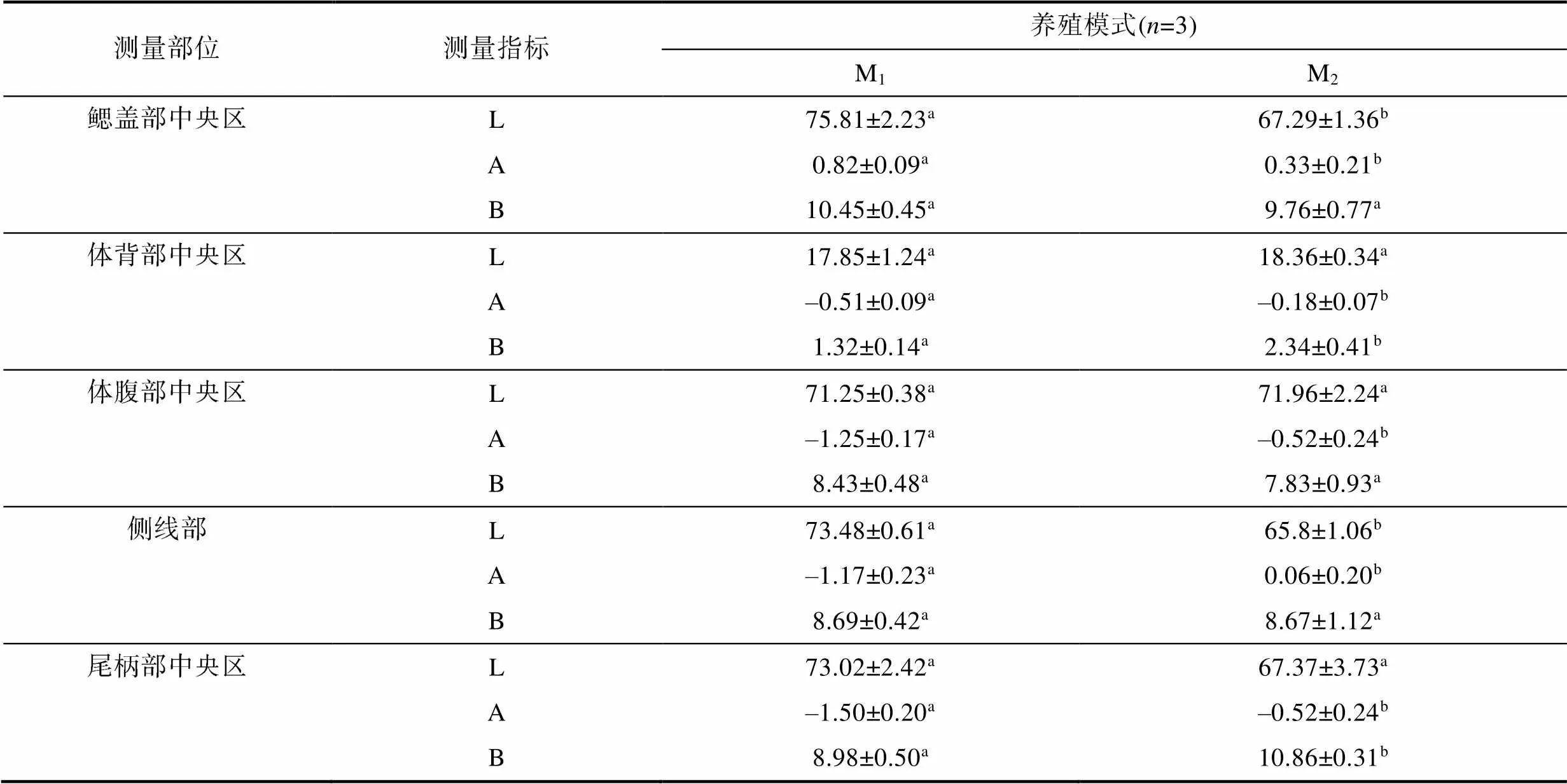
Tab.1 Hunter LAB values of body surface color of experimental C. auratus gibelio in two pond aquaculture modes
注: a、b標注組間差異(<0.05), 字母相同表示無差異, 下同
2.2 背肌物性
由表2可見, 在所測7項背肌物性指標中, 除黏性、內聚性和回復性等3項指標均呈M1≈M2(>0.05)外, 余下的硬度、彈性、膠黏性和耐咀性等4項指標均呈M1>M2(<0.05)。由此可知, M1和M2實驗個體間背肌物性的相似性僅為42.8%, 具有較好的區分度,且M1實驗個體的背肌較M2更顯緊實且富彈性, 具更佳的食用口感。
2.3 一般營養成分
由表3可見, 全魚一般營養成分中除灰分和粗蛋白含量均呈M1≈M2(>0.05)外, 水分和粗脂肪含量分別呈M1>M2(<0.05)和M2>M1(<0.05), 表明M1實驗個體較M2具更高的脂肪氧化代謝水平。
2.4 脂肪酸組成
由表4可見, M1與M2實驗個體在脂肪酸組成上的異同主要表現為: (1) 所檢4種飽和脂肪酸的總含量ΣSFA呈M1

圖1 兩種池塘養殖模式實驗鯽的側視圖
表2 兩種池塘養殖模式下實驗鯽的背肌物性差異
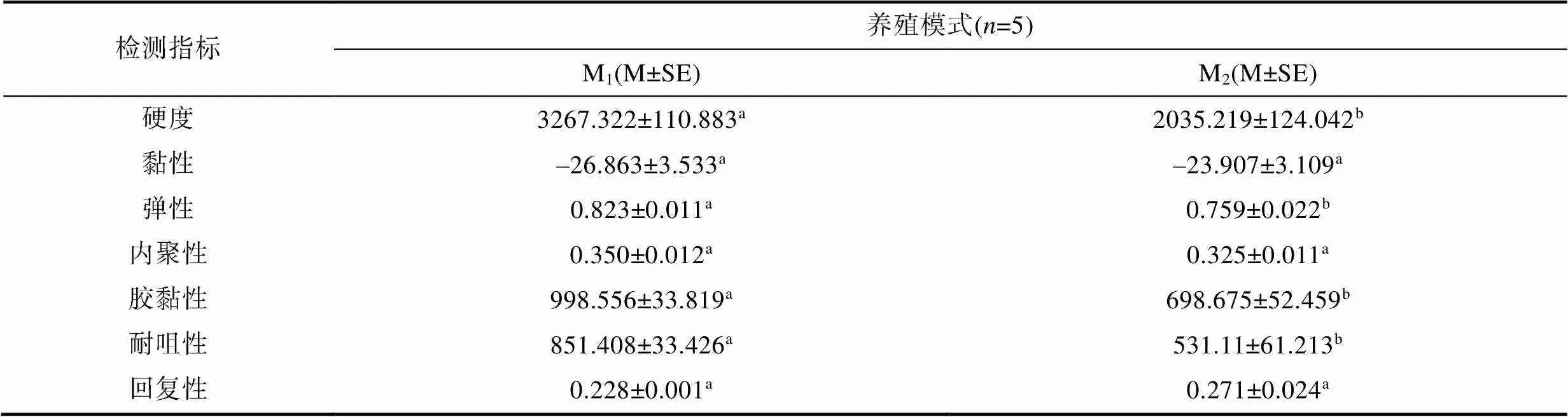
Tab.2 Physical differences in dorsal muscle properties of experimental C. auratus gibelio in two pond aquaculture modes
表3 兩種池塘養殖模式下實驗鯽的全魚常規營養成分(單位: %)
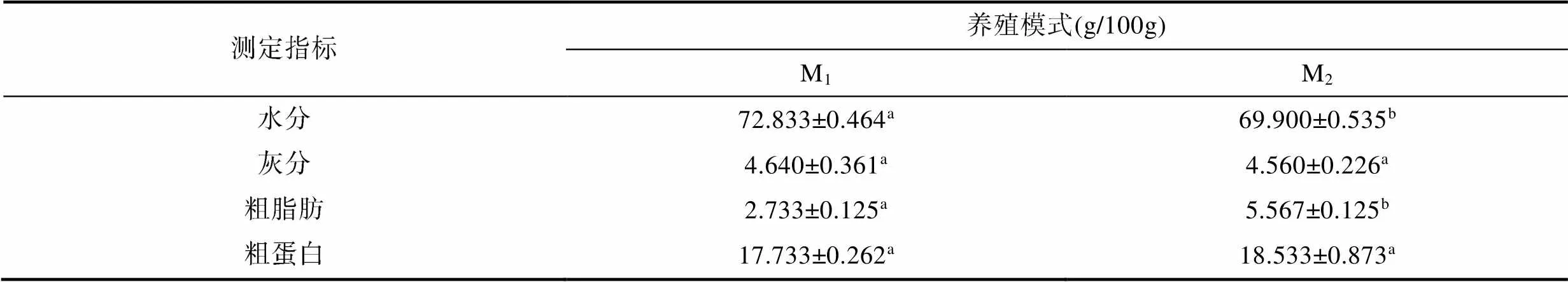
Tab.3 Whole-body nutrient composition of experimental C. auratus gibelio in two pond aquaculture modes
表4 兩種池塘養殖模式下異實驗鯽脂肪酸組成的差異
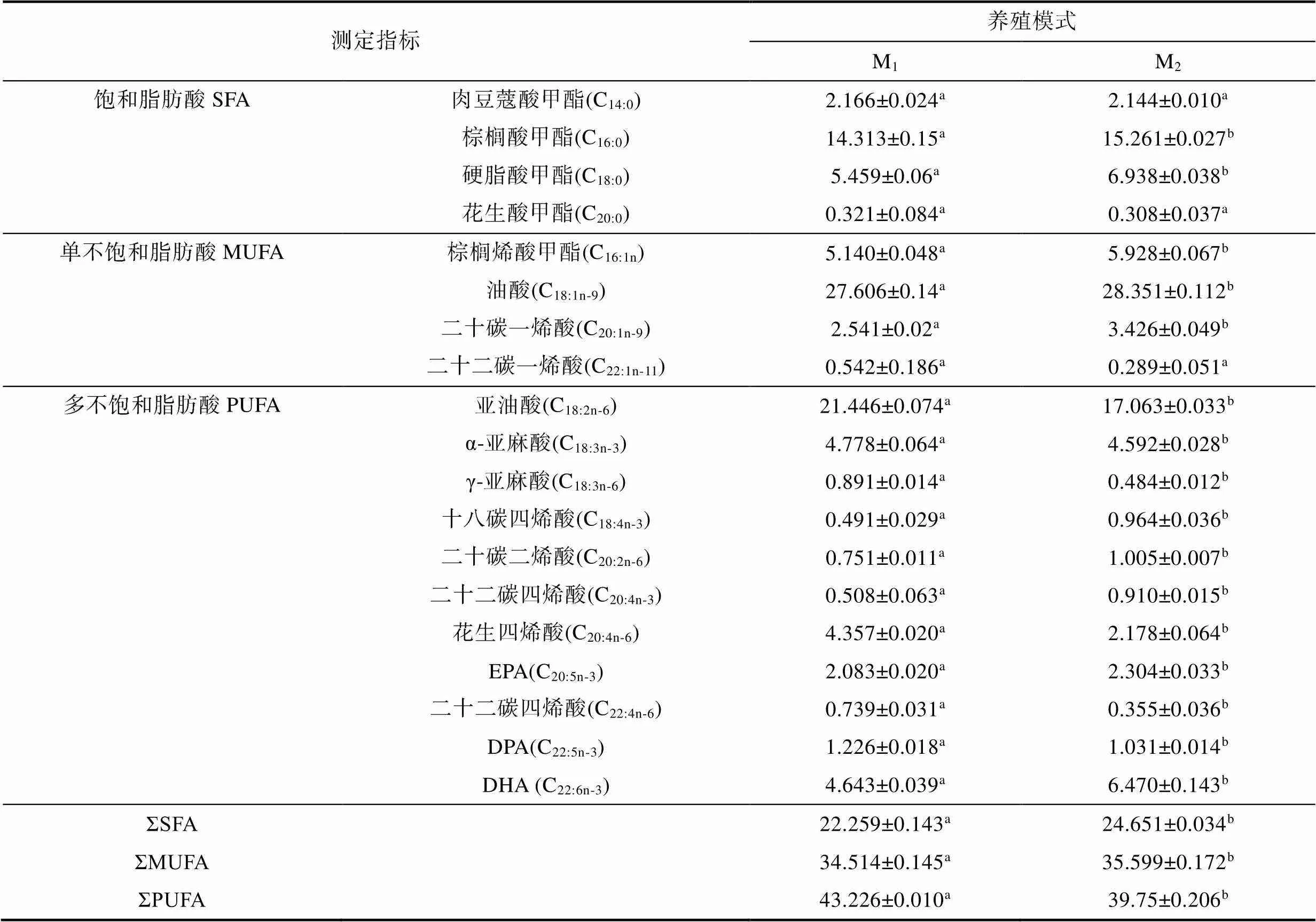
Tab.4 Differences in fatty acid composition of experimental C. auratus gibelio in two pond aquaculture modes
2.5 氨基酸組成
由表5可見, 在所測17種氨基酸含量上, M1與M2實驗個體間具顯著差異的僅為組氨酸, 呈M1>M2(<0.05); 在各類氨基酸含量上, 除∑EAA和∑EAA/∑NEAA均呈M1≈M2(>0.05)外, ∑SEAA、∑NEAA和∑FAA均呈M1>M2(<0.05)。由此可知, M1實驗個體在氨基酸水平上較M2具更高的食用營養價值。
表5 兩種池塘養殖模式下異育銀鯽的氨基酸組成差異(單位: g/100g)
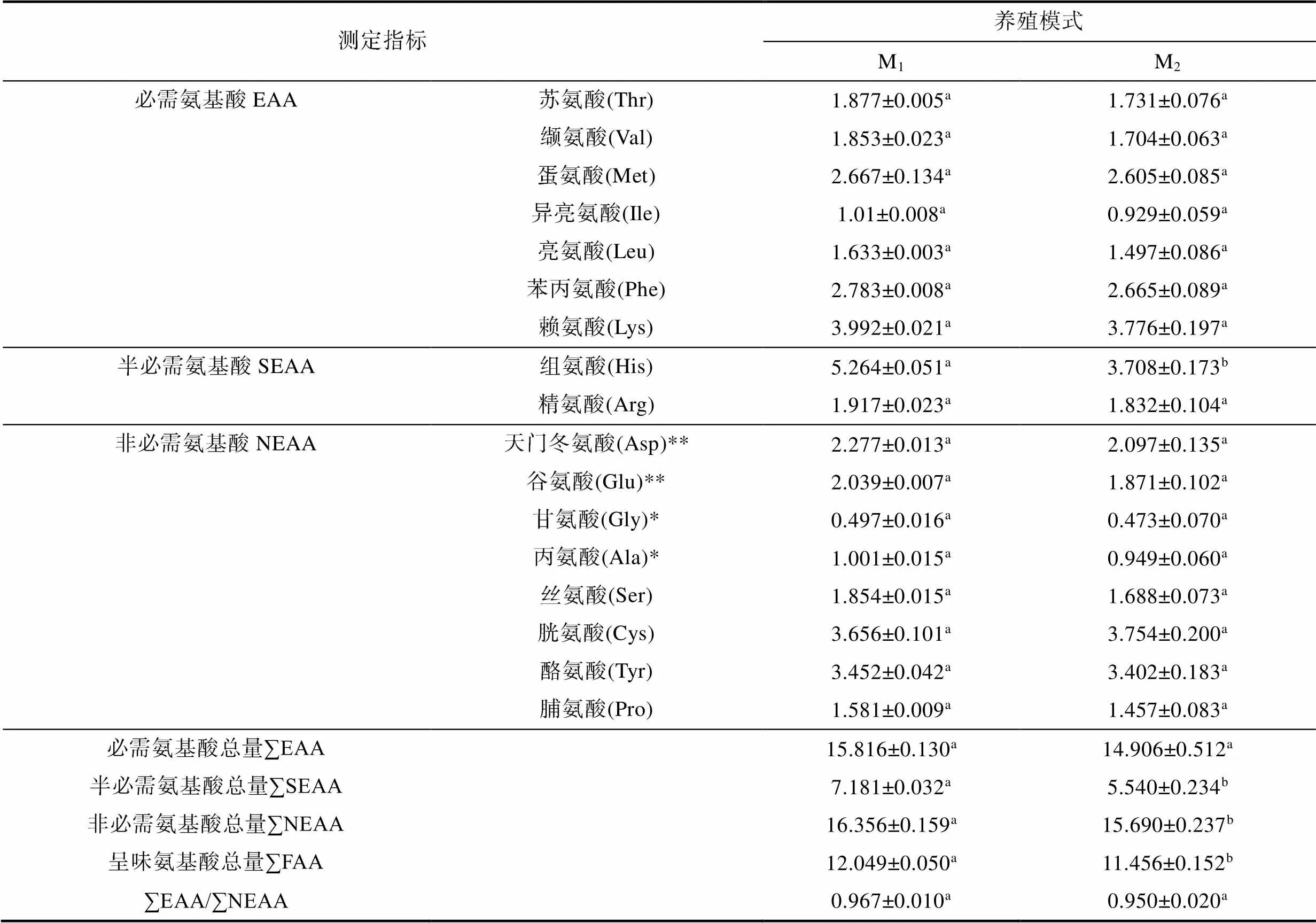
Tab.5 Differences in amino acid composition of experimental C. auratus gibelio in two pond aquaculture modes
注: 上標**、*分別示甘味氨基酸和鮮味氨基酸
2.6 耗氧率、排氨率與窒息點
由圖2和圖3可見, M1與M2實驗個體間在耗氧率、排氨率與窒息點上的異同主要表現為: (1) M1實驗個體的晝均、夜均和日均耗氧率均與M2實驗個體無顯著差異(>0.05), 且兩者的耗氧晝夜節律也均呈晝均>日均>夜均(<0.05), 表明異育銀鯽屬晝行性魚類, 其晝均、夜均和日均耗氧率以及晝夜耗氧節律均未因池塘養殖模式的變更而發生實質性改變; (2) M1實驗個體的晝均、夜均和日均排氨率均與M2實驗個體無顯著差異(>0.05), 但兩者的晝夜排氨節律則分別呈晝均≈夜均≈日均(>0.05)和晝均>日均>夜均(<0.05), 表明池塘養殖模式的變更可引起實驗鯽晝夜排氨晝夜節律的顯著改變; (3) M1實驗個體的窒息點水中氧含量顯著低于M2實驗個體(<0.05), 表明M1較M2實驗個體具更強的耐低氧能力。

圖2 兩種池塘養殖模式下實驗鯽的耗氧率和排氨率
注: 上標a、b示組間差異, 上標x、y、z示組內差異, 上標相同字母示無顯著差異
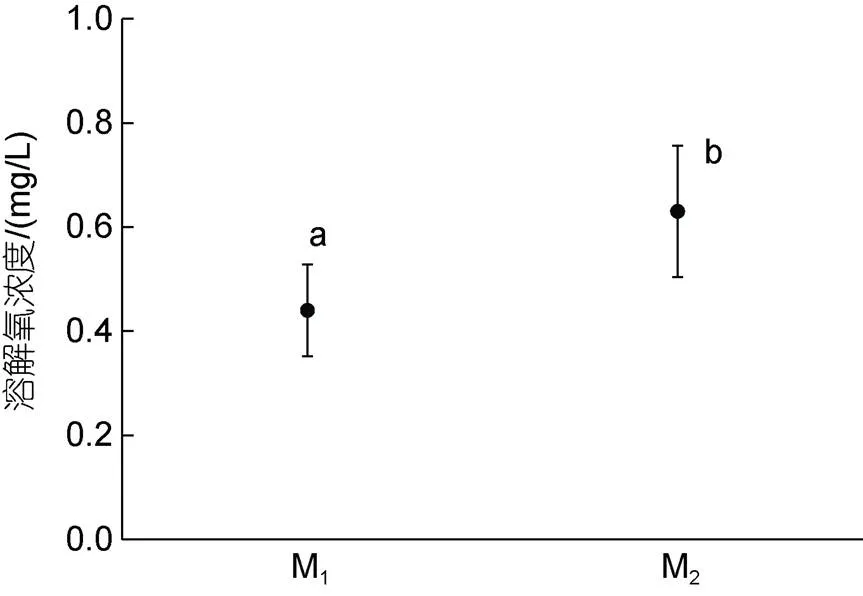
圖3 兩種池塘養殖模式下實驗鯽窒息點的水中含氧量
2.7 臟器消化酶和抗氧化酶活力
由圖4和圖5可見, M1與M2實驗個體間在臟器消化酶和抗氧化酶活力上的異同主要表現為: (1) 從臟器消化酶活力看, 實驗鯽的胃蛋白酶和肝脂肪酶活力均呈M1≈M2(>0.05), 而腸淀粉酶活力則呈M1
3 討論
3.1 實驗鯽取食策略與養殖品質間的相關性
動物的運動代謝和攝食代謝既是兩個互為關聯的重要生理過程(Hicks, 2004; Fu, 2007), 也是影響進而決定魚類養成品質的重要代謝途徑(王志錚等, 2012, 2013a)。因此, 本研究所涉M1和M2實驗群體間的養成品質差異, 無疑與兩者分別所采取的積極取食為主和伏擊取食為主的取食策略密切相關(徐英杰等, 2023)。即(1)鯽屬側扁型底層魚類, M2實驗個體鰓蓋中央區和側線部體色較M1均顯著偏黑(<0.05), 具更好的擬境隱蔽性的結果(表1, 圖1), 更有助于其貫徹以伏擊取食為主的取食策略; (2) 從背肌質構和全魚一般營養成分組成看, M1實驗個體的背肌硬度、彈性、膠黏性和耐咀性均顯著大于M2(<0.05), 更顯緊實且富彈性(表2), 以及僅體脂含量顯著小于M2(<0.05)和水分含量顯著大于M2(< 0.05), 具更高體脂氧化代謝水平(表3)的結果, 既與大湖養殖模式團頭魴背肌亮度、白度、彈性、咀嚼性、回復性和水分含量均顯著大于普通池塘養殖模式, 而粗脂肪含量則顯著低于池塘養殖模式的結果(李溫蓉等, 2022)相近, 也與種草養殖模式草魚肌肉白度、硬度、咀嚼性、回復性以及水分含量、膠原蛋白含量和膠原纖維致密度均顯著高于普通養殖模式的結果(張曦等, 2021; 溫利等, 2022)相似, 在反映M1實驗個體以積極取食為主的取食策略的同時, 也表明改善養殖環境可顯著增進養殖魚類的運動能力并增強其攝食代謝強度; (3) 一般而言, 脂類的營養價值主要取決于PUFA的類型和含量。從全魚脂肪酸和氨基酸組成看, 本研究中M1實驗個體全魚ΣSFA與ΣMUFA均顯著小于M2(<0.05), ΣPUFA顯著大于M2(<0.05) (表4), 以及∑SEAA、∑NEAA、∑FAA和組氨酸含量均顯著大于M2(<0.05) (表5)的結果, 在揭示實驗鯽體脂氧化代謝的主要底物為SFA與MUFA的同時, 也反映了M2實驗個體因采取以殘餌和腐屑為主要食源的伏擊取食策略, 致使其食用營養價值明顯劣于M1。綜上可知, 造成M1和M2實驗個體間養殖品質差異的本質系兩者為更好地適應各自所處生存環境和食源質量而采取不同生存對策所致。另, 鑒于M1養殖模式實驗鯽養殖品質明顯優于M2的研究結果, 與適量種植水生植物可顯著改善池塘養殖環境(Pokorny, 1990; 周遺品等, 2011)并有效提高水產養殖對象養成品質(劉鑫等, 2003; 徐增洪等, 2016)的結論相吻合, 建議在具體養殖生產實踐中應以優化養殖水質和提高食源質量為導向, 通過提高目標養殖對象的生存福利以切實提升其養殖品質。
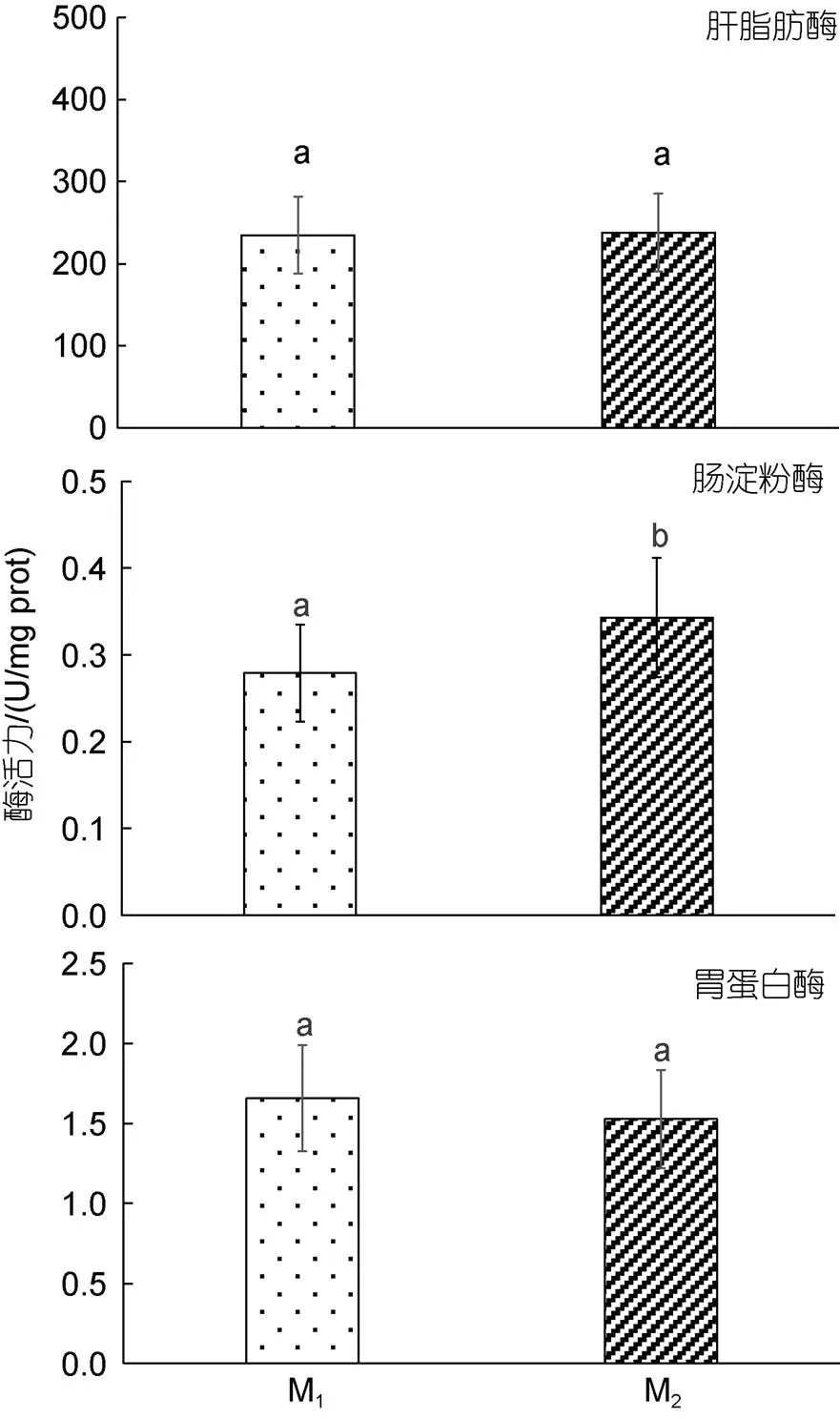
圖4 兩種池塘養殖模式下實驗鯽的臟器消化酶活力
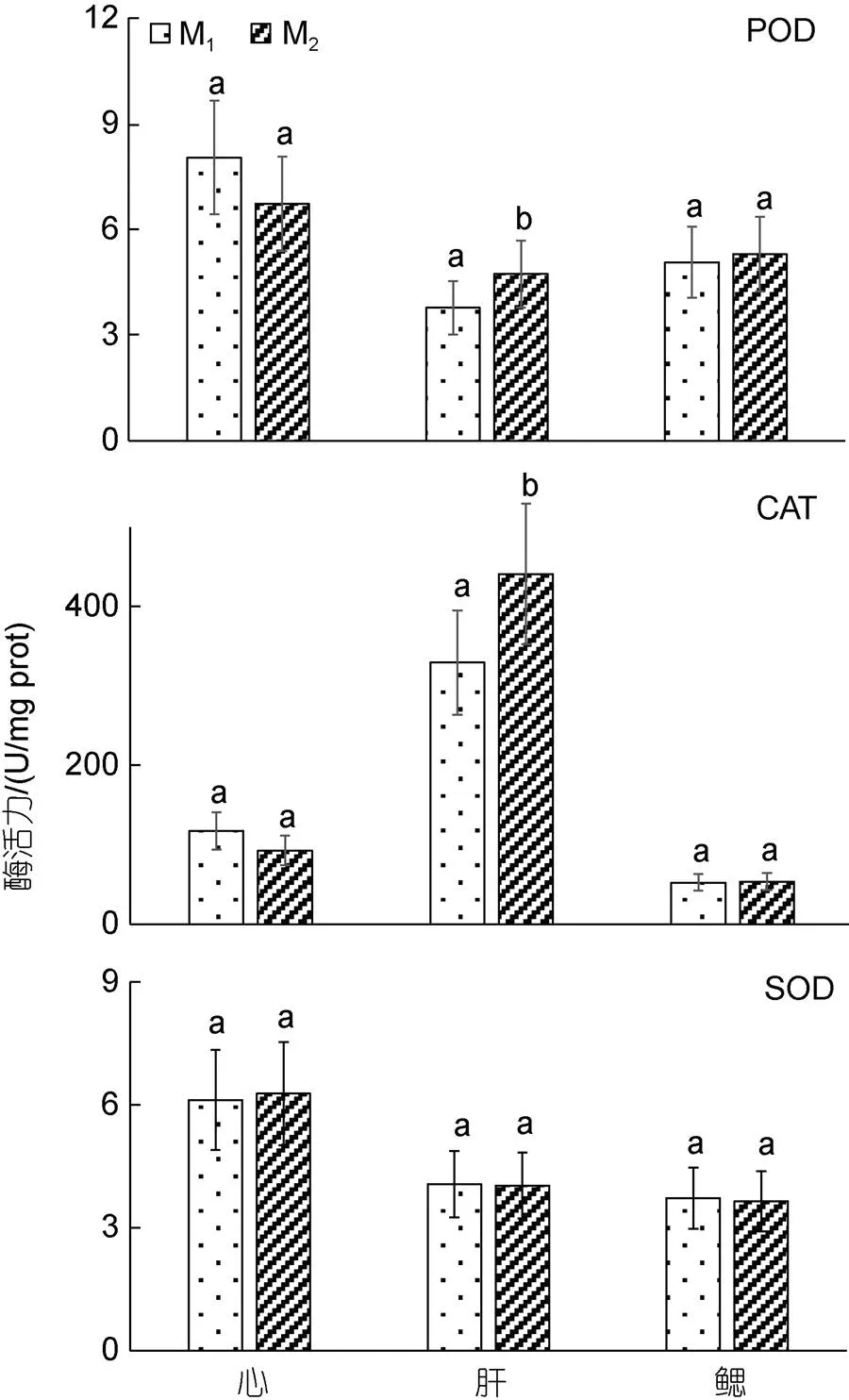
圖5 兩種池塘養殖模式下實驗鯽臟器的抗氧化酶活力
3.2 實驗鯽臟器生理代謝機能與取食對策間的相關性
生命代謝特征既是表征水生生物所處生存環境優劣程度的重要依據, 也是反映水生生物健康程度和養成品質的重要指標。目標水產養殖對象往往會應養殖模式的改變而采取相應的生存對策, 并表露出不同的生命代謝特征(王志錚等, 2013b)。陳雨等(2022)由M1、M2實驗群體生物學性狀對體質量影響效應差異, 得出兩者在-生存對策選擇軸上表露出明顯偏離傾向的推論, 為我們從機體生理代謝角度闡析兩者間取食對策的差異提供了重要啟示。研究表明, 活性器官的臟器系數與機體代謝水平呈正相關(Itazawa, 1983; Oikawa, 1992)。經稱量和計算, M1實驗個體心、鰓、胃、肝、腸等活性器官的質量及其臟器系數均顯著大于M2(<0.05) (陳雨等, 2022), 因而M1實驗個體的機體代謝水平應遠高于M2, 這就為M1實驗個體貫徹并實施以積極取食為主的取食策略奠定了扎實的生理基礎。胃、肝、腸作為魚類的重要消化臟器和各種營養物質消化吸收與代謝的主要場所, 其發達程度直接影響魚類的生長速度(馬細蘭等, 2009)。本研究中, 腸淀粉酶活力呈M1 (1) M1、M2實驗個體在體表色差、背肌物性和全魚一般營養成分上均具較好的區分度。其中, M1實驗個體的背肌更顯緊實且富彈性, 具更高的機體脂肪氧化代謝水平, 而M2實驗個體的體色則具更好的擬境隱蔽性。 (2) 實驗鯽體脂氧化代謝的主要底物為SFA和MUFA。因M2實驗個體的主要食源為殘餌和腐屑, 劣于M1, 致使其腸淀粉酶活力顯著大于M1, 而全魚ΣPUFA、組氨酸和各類氨基酸含量均顯著低于M1。 (3) M1、M2實驗個體的養成品質差異與它們分別所采取的積極取食為主和伏擊取食為主的取食策略密切相關。造成實驗鯽實施不同取食策略的內在機制主源于其機體內各類活性臟器對所處生存環境和食源質量的綜合生理反應, 尤以肝臟抗氧化生理為甚。即M1實驗個體依仗更強的機體耐低氧能力和肝臟抗氧化能力, 通過權衡“消耗—獲利”, 在運動強度上采取了以積極取食為主的高能耗代謝策略, 為抵制脂肪過度消耗, 在蛋白質利用上采取了晝夜更顯均衡的排氨機制; M2實驗個體面對高企的肝臟抗氧化壓力和相對較弱的耐低氧能力, 在運動強度上采取了以伏擊取食為主的低能耗代謝策略, 在蛋白質利用上采取了順應晝夜耗氧節律的節能型排氨機制。 (4) 鑒于M1實驗個體的養成品質明顯優于M2, 建議在具體養殖生產實踐中應以優化養殖水質和提高食源質量為導向, 通過提高目標養殖對象的生存福利以切實提升其養殖品質。 馬細蘭, 張勇, 周立斌, 等, 2009. 脊椎動物雌雄生長差異的研究進展[J]. 動物學雜志, 44(2): 141-146. 王志錚, 楊磊, 朱衛東, 2012. 三種養殖模式下日本鰻鱺養成品的形質差異[J]. 應用生態學報, 23(5): 1385-1392. 王志錚, 付英杰, 楊磊, 等, 2013a. 三種養殖模式下日本鰻鱺()養成品體色和肌肉品質的差異[J]. 海洋與湖沼, 44(4): 1042-1049. 王志錚, 趙晶, 楊磊, 等, 2013b. 三種養殖模式下日本鰻鱺()養成品血清生化指標和臟器消化酶、抗氧化酶活力的差異[J]. 海洋與湖沼, 44(2): 403-408. 戈賢平, 2010. 我國大宗淡水魚養殖現狀及產業技術體系建設[J]. 中國水產(5): 5-9. 劉松巖, 2006. 環境脅迫對中華鱘體內自由基水平和抗氧化酶活力的影響[D]. 武漢: 華中農業大學. 劉鑫, 王哲, 張一, 等, 2003. 兩種沉水植物對黃顙魚()夏花培育水體主要水質因子的影響[J]. 湖泊科學, 15(2): 184-188. 李治, 謝小軍, 曹振東, 2005. 攝食對南方鲇耗氧和氨氮排泄的影響[J]. 水生生物學報, 29(3): 247-252. 李溫蓉, 田明禮, 安玥琦, 等, 2022. 池塘養殖和大湖養殖對“華海1號”團頭魴魚肉品質的影響[J]. 水產學報, 46(7): 1220-1234. 楊興麗, 張芹, 李治勛, 等, 2022. 異育銀鯽“中科5號”池塘生態增效養殖模式總結[J]. 科學養魚(10): 79-80. 楊程, 劉哲宇, 朱衛東, 等, 2016. 水溫對水產經濟動物棘胸蛙()蝌蚪耗氧率、窒息點和排氨率的影響[J]. 海洋與湖沼, 47(6): 1233-1240. 張曦, 萬瑩, 席汝娟, 等, 2021. 種青養殖模式提升草魚肌肉中膠原蛋白的相關研究[J]. 淡水漁業, 51(5): 70-75. 陳雨, 戎華南, 劉哲宇, 等, 2022. 兩種池養模式下異育銀鯽()生物學表型對體質量影響效果的差異分析[J]. 海洋與湖沼, 53(5): 1161-1169. 周遺品, 劉雯, 雷澤湘, 等, 2011. 銅錢草對城市生活污水的凈化效果[J]. 仲愷農業工程學院學報, 24(2): 9-12. 鄭勁松, 2004. 池塘主養異育銀鯽放養模式的研究[J]. 安徽農業科學, 32(2): 353-354. 徐英杰, 戎華南, 陳雨, 等, 2023. 兩種池塘養殖模式下異育銀鯽()的形質特征差異分析[J]. 海洋與湖沼, 54(1): 204-213. 徐增洪, 劉國鋒, 水燕, 等, 2016. 人工栽培水草對池塘養殖生態環境和蝦蟹生長的影響[J]. 江蘇農業科學, 44(6): 328-330, 331. 黃溢明, 1982. 環境因素對魚類某些生理機能的影響[J]. 生理科學進展, 13(2): 162-167. 溫利, 田明禮, 安玥琦, 2022. 不同養殖模式對草魚的營養與食用品質的影響[J]. 華中農業大學學報, 41(3): 244-251. BUSHNELL P G, JONES D R, FARRELL A P, 1992. The arterial system [J]. Fish Physiology, 12: 89-139. EVANS D H, PIERMARINI P M, CHOE K P, 2005. The multifunctional fish gill: Dominant site of gas exchange, osmoregulation, acid-base regulation, and excretion of nitrogenous waste [J]. Physiological Reviews, 85(1): 97-177. FU S J, CAO Z D, PENG J L, 2007. Effect of feeding and fasting on excess post-exercise oxygen consumption in juvenile southern catfish (Chen) [J]. Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology, 146(3): 435-439. HICKS J W, BENNETT A F, 2004. Eat and run: prioritization of oxygen delivery during elevated metabolic states [J]. Respiratory Physiology & Neurobiology, 144(2/3): 215-224. ITAZAWA Y, OIKAWA S, 1983. Metabolic rates in excised tissues of carp [J]. Experientia, 39(2): 160-161. OIKAWA S, TAKEMORI M, ITAZAWA Y, 1992. Relative growth of organs and parts of a marine teleost, the porgy,, with special reference to metabolism-size relationships [J]. Japanese Journal of Ichthyology, 39(3): 243-249. POKORNY J, KVET J, ONDOK J P, 1990. Functioning of the plant component in densely stoked fish ponds [J]. Bulletin of Ecology, 21(3): 44-48. WAGNER E J, ARNDT R E, BROUGH M, 2001. Comparative tolerance of four stocks of cutthroat trout to extremes in temperature, salinity, and hypoxia [J]. Western North American Naturalist, 61(4): 434-444. DIFFERENCES OF QUALITY AND VISCERAL-RELATED PHYSIOLOGICAL CHARACTERISTICS OFIN TWO POND AQUACULTURE MODES LI You-Zhi1, RONG Hua-Nan1, 2, CHEN Yu1, SHENTU Ji-Kang3, WANG Zhi-Zheng1 (1. Fisheries College, Zhejiang Ocean University, Zhoushan 316022, China; 2. Cixi Fisheries Technology Extension Center, Cixi 315300, China; 3. Ningbo Ocean and Fisheries Research Institute, Ningbo 315048, China) Thelarvae were cultured in common-culture pond mode (M1) and the poly-culture pond mode (M2) for 7 months. The body surface color difference, back muscle physical properties, nutrient composition and ammonia excretion rate of whole fish, oxygen consumption rate, asphyxia point, and the activities of digestive enzymes and antioxidant enzymes in organs were measured. In order to systematically study the differences in cultivation quality and visceral-related physiological characteristics between the two groups. The results showed that: (1) there was a good discrimination degree in the body surface color difference between the two groups. Among them, the body color of M2experiment was closer to the background color at the bottom of the pond, which had better simulated environment camouflage;(2) Among the seven physical properties of the back muscles measured, the hardness, elasticity, adhesiveness and chew ability showed significant differences between the two groups (M1>M2) (<0.05);(3) In the general nutritional composition of whole fish,the contents of ash and crude protein were M1≈ M2(>0.05), while the contents of water and crude fat were M1>M2(<0.05) and M2>M1(<0.05), respectively;(4) Among the 17 amino acids tested, only histidine showed significant difference in content, showing M1>M2(<0.05), only ∑SEAA, ∑NEAA and ∑FAA had significant differences in the contents of various amino acids, and they were M1>M2(<0.05);(5) A total of four saturated fatty acids, five monounsaturated fatty acids and ten polyunsaturated fatty acids were detected, among which ΣMUFA and ΣSFA showed M1 aquaculture mode;; body surface color difference; dorsal muscle texture; nutrient composition; physiological characteristics *國家星火計劃項目, 2013GA701022號; 中央財政重大農業技術推廣項目, YY2017003號; 慈溪市農業科技重點項目, CN201901號; 寧波市重大科技任務攻關項目, 2022Z183號。李有志, 碩士研究生, E-mail: 1156604931@qq.com; 同等貢獻第一作者: 戎華南, 高級工程師, 碩士生導師, E-mail: 13706742262@139.com 王志錚, 教授, E-mail: wzz_1225@163.com 2022-12-28, 2023-02-22 Q953; S96 10.11693/hyhz202212003414 結論
猜你喜歡
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42音樂探索(2022年2期)2022-05-30 21:01:37中學生數理化·中考版(2022年11期)2022-02-16 07:01:20小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00中國特種設備安全(2018年11期)2019-01-08 02:08:32小學科學(學生版)(2018年7期)2018-08-13 09:33:04發明與創新(2016年38期)2016-08-22 03:02:52太空探索(2016年5期)2016-07-12 15:17:55鄭州大學學報(醫學版)(2015年2期)2015-02-27 14:50:46
