樟芝深層發(fā)酵多糖對(duì)抗生素相關(guān)性腹瀉小鼠腸道菌群的調(diào)節(jié)作用
2023-08-12 00:41:18李華祥陸春雷葉青雅趙靈惠高亞軍楊振泉
食品科學(xué) 2023年13期
關(guān)鍵詞:小鼠
李華祥,吉 丹,陸春雷,葉青雅,趙靈惠,高亞軍,高 璐,楊振泉
(揚(yáng)州大學(xué)食品科學(xué)與工程學(xué)院,江蘇 揚(yáng)州 225127)
人類的腸道菌群是一個(gè)由數(shù)萬(wàn)億細(xì)菌組成的微生物生態(tài)系統(tǒng),其中絕大多數(shù)屬于厚壁菌門(Firmicutes)和擬桿菌門(Bacteroidetes),包括雙歧桿菌(Bifidobacterium)、乳酸桿菌(Lactobacillus)、腸球菌(Enterococcus)、產(chǎn)氣莢膜梭菌(Clostridium perfringens)和假單胞菌(Pseudomonas)等[1]。腸道菌群的組成、代謝、生理、營(yíng)養(yǎng)和免疫活動(dòng)通常高度穩(wěn)定,但也可能受到年齡、飲食、生活方式、抗生素的使用等多種因素的影響[2]。目前,抗生素參與疾病治療的現(xiàn)象越來(lái)越普遍,但廣譜抗生素的大量使用不僅會(huì)影響目標(biāo)病原體,還會(huì)干擾腸道中其他共生微生物,降低其定植能力,導(dǎo)致腸道細(xì)菌的多樣性、均勻度和分類豐富度迅速下降[3]。在長(zhǎng)期使用抗生素的過(guò)程中,有益菌大量流失[4],同時(shí)耐藥的有害微生物數(shù)量不斷增加,導(dǎo)致有害菌逐漸占據(jù)主導(dǎo)地位,最終導(dǎo)致患抗生素相關(guān)性腹瀉及其他慢性疾病的風(fēng)險(xiǎn)急劇增加[3,5-6]。有研究指出,即使是短期服用抗生素也會(huì)導(dǎo)致耐藥菌群在人類腸道中穩(wěn)定存在數(shù)年[7]。可見(jiàn),調(diào)節(jié)腸道菌群穩(wěn)態(tài)和恢復(fù)腸道菌群結(jié)構(gòu)對(duì)防治部分慢性疾病至關(guān)重要。
益生元是一種可促進(jìn)胃腸道中有益菌生長(zhǎng)并對(duì)致病菌發(fā)揮拮抗作用的活性物質(zhì),而一些不易被消化的寡糖和多糖常常具有與益生元相似的功能[8]。自然界中的多糖按來(lái)源可分為植物多糖、動(dòng)物多糖及微生物多糖[9]。其中,微生物多糖中的真菌多糖能夠通過(guò)選擇性刺激有益于宿主健康的一種或有限數(shù)量細(xì)菌的生長(zhǎng)活性來(lái)調(diào)節(jié)腸道菌群并發(fā)揮益生元作用[10-11]。此外,真菌多糖還具有改善腸道完整性及減輕腸黏膜損傷等功效,有利于調(diào)節(jié)腸道菌群平衡[12]。
樟芝(Antrodia cinnamomea)又名牛樟芝、牛樟菇等,是一種珍稀的藥食兩用蕈菌,具有解酒保肝、抗炎、抗癌、抗腫瘤、抗病毒、降血脂及調(diào)劑免疫等多種生物活性[13]。近年來(lái),研究者從樟芝子實(shí)體和發(fā)酵菌絲體中分離出200余種活性物質(zhì)[14],其中,多糖是最主要的活性物質(zhì)之一,具有較高的研發(fā)價(jià)值。但目前關(guān)于樟芝多糖的研究主要集中于活性及藥理方面[15-16],對(duì)其調(diào)節(jié)小鼠腸道菌群的相關(guān)研究較少。因此,本實(shí)驗(yàn)首先提取和純化樟芝深層發(fā)酵所產(chǎn)胞內(nèi)多糖(Antrodia cinnamomeaintracellular polysaccharides,AIPS)及胞外多糖(Antrodia cinnamomeaexopolysaccharides,AEPS),再對(duì)其進(jìn)行組成分析及結(jié)構(gòu)表征,最后探究其對(duì)抗生素相關(guān)性腹瀉小鼠腸道菌群的調(diào)節(jié)作用。
1 材料與方法
1.1 動(dòng)物、材料與試劑
小鼠為無(wú)特定病原體(SPF)級(jí)ICR小鼠(6 周齡、體質(zhì)量(20±2)g,使用許可證號(hào):SYXK(蘇)2016-0019),均由揚(yáng)州大學(xué)比較醫(yī)學(xué)中心提供。動(dòng)物實(shí)驗(yàn)操作均按揚(yáng)州大學(xué)實(shí)驗(yàn)動(dòng)物倫理委員會(huì)的相關(guān)規(guī)定執(zhí)行。
樟芝菌株(ATCC 200183)購(gòu)自美國(guó)菌種保存中心,目前保存于揚(yáng)州大學(xué)食品科學(xué)與工程學(xué)院食品質(zhì)量與安全控制實(shí)驗(yàn)室。
鹽酸林可霉素(lincomycin hydrochloride,LIH)上海生工生物工程有限公司;低聚果糖 上海麥克林生化科技股份有限公司;胃蛋白酶 福州飛凈生物科技有限公司;BCA蛋白試劑盒 上海碧云天生物技術(shù)有限公司;α-淀粉酶(4 000 U/g) 上海源葉生物科技有限公司;無(wú)水乙醇、正丁醇、三氯甲烷、苯酚、硫酸、多糖分子質(zhì)量標(biāo)準(zhǔn)品及單糖標(biāo)準(zhǔn)品 國(guó)藥集團(tuán)化學(xué)試劑有限公司;所有試劑均為分析純。
1.2 儀器與設(shè)備
Infinite F50酶標(biāo)儀 上海勒菲生物科技有限公司;RE-52B旋轉(zhuǎn)蒸發(fā)儀 上海雅蓉生化儀器設(shè)備有限公司;AlpHA1-2LD冷凍干燥機(jī) 德國(guó)Martin Christ公司;670-IR+610-IR紅外光譜儀 美國(guó)Varian公司;5 L發(fā)酵罐 匯森生物設(shè)備鎮(zhèn)江有限公司;1525高效液相色譜儀 美國(guó)Waters公司;ICS-5000離子色譜儀美國(guó)戴安公司。
1.3 方法
1.3.1 樟芝5 L罐深層發(fā)酵
將5 L玻璃發(fā)酵罐空消(121 ℃、30 min)后,向其中裝入3 L發(fā)酵培養(yǎng)基(葡萄糖 20 g/L、酵母浸出粉2 g/L、MgSO4·7H2O 3 g/L、KH2PO43 g/L,pH 4.5)并進(jìn)行實(shí)消(121 ℃、20 min)。冷卻后,按1×106個(gè)/mL的接種量進(jìn)行接種,在26 ℃、150 r/min及0.3 m3/h通氣量條件下發(fā)酵10 d。
1.3.2 樟芝多糖的提取及純化
1.3.2.1 樟芝胞內(nèi)多糖的提取
將5 L罐樟芝發(fā)酵液用4 層紗布過(guò)濾,收集濾渣(菌絲體)并75 ℃烘干。稱取10 g干燥并粉碎的樟芝菌絲體粉末,加入100 mL去離子水,95 ℃水浴提取2 h,重復(fù)提取2 次,合并上清液并蒸發(fā)濃縮至20 mL,加入3 倍體積的無(wú)水乙醇,4 ℃靜置過(guò)夜,離心(8 000 r/min、10 min、4 ℃)、棄上清液,沉淀用超純水復(fù)溶后真空冷凍干燥,即得樟芝胞內(nèi)粗多糖粉末。為獲得足夠量的AIPS,上述操作可重復(fù)多次。
1.3.2.2 樟芝胞外多糖的提取
將5 L罐樟芝發(fā)酵液用4 層紗布過(guò)濾,收集濾液,離心(6 000 r/min、10 min、4 ℃)去沉淀,收集上清并蒸發(fā)濃縮至20 mL,再加入3 倍體積的無(wú)水乙醇,4 ℃靜置過(guò)夜,離心(8 000 r/min、10 min、4 ℃)棄上清液,沉淀用超純水復(fù)溶后真空冷凍干燥,即得樟芝胞外粗多糖粉末。為獲得足夠量的AEPS,上述操作可重復(fù)多次。
1.3.2.3 樟芝多糖的純化
采用Sevag法[17]除去樟芝多糖(AEPS及AIPS)中的蛋白質(zhì)。具體步驟:將正丁醇和三氯甲烷按照體積比1∶4混合,將混合液加入至5 倍體積的樟芝多糖水溶液中,劇烈振蕩1 min,4 000 r/min離心10 min,取上層水相,重復(fù)多次后,蒸發(fā)濃縮去除有機(jī)溶劑即可。
1.3.3 樟芝多糖組成分析
1.3.3.1 總糖及蛋白質(zhì)量分?jǐn)?shù)測(cè)定
總糖質(zhì)量分?jǐn)?shù)測(cè)定:以葡萄糖為標(biāo)準(zhǔn)品,用苯酚-硫酸法測(cè)定樟芝多糖中的總中性糖質(zhì)量分?jǐn)?shù);蛋白質(zhì)量分?jǐn)?shù)測(cè)定:采用BCA蛋白試劑盒測(cè)定樟芝多糖中的總蛋白質(zhì)量分?jǐn)?shù),操作步驟嚴(yán)格按照說(shuō)明書進(jìn)行。
1.3.3.2 單糖組成測(cè)定
采用HPLC法檢測(cè)單糖組成。樣品前處理:用2 mol/L的HCl溶液于96 ℃水解多糖樣品4 h,再用2 mol/L的NaOH溶液中和;檢測(cè)條件:儀器為ICS-5000離子色譜儀,色譜柱為CarboPac PA20,柱溫為45 ℃,流動(dòng)相為98.2%的水和1.8%的250 mmol/L NaOH溶液,流速為0.5 mL/min,洗脫30 min;單糖標(biāo)準(zhǔn)品:阿拉伯糖、半乳糖、葡萄糖、木糖、甘露糖、半乳糖醛酸及葡萄糖醛酸,純度均為色譜純。
1.3.4 樟芝多糖分子質(zhì)量分布的測(cè)定
采用高效凝膠過(guò)濾色譜法(high performance gel filtration chromatography,HPGFC)測(cè)定樟芝多糖的分子質(zhì)量分布情況。檢測(cè)條件:儀器為1525高效液相色譜儀,色譜柱為Ultrahydrogel?水溶性凝膠柱(300 mm×7.8 mm),柱溫為45 ℃,流動(dòng)相為0.1 mol/L的NaNO3溶液,流速為0.9 mL/min;右旋糖酐(Dextran)多糖標(biāo)準(zhǔn)品:相對(duì)分子質(zhì)量分別為135 350、3 800、9 750及2 700,純度均為色譜純。
1.3.5 樟芝多糖的紅外光譜分析
在室溫下,稱取干燥的樟芝多糖粉末與光譜級(jí)KBr粉末一起充分研磨,壓片,以純KBr粉末薄片作為空白背景,在4 000~400 cm-1的波數(shù)范圍內(nèi)進(jìn)行掃描分析。
1.3.6 樟芝多糖體外抗消化能力的測(cè)定
1.3.6.1 抗α-淀粉酶消化能力的測(cè)定
抗α-淀粉酶消化能力的測(cè)定參照文獻(xiàn)[18],取10 mL磷酸鹽緩沖液,用1 mol/L的HCl溶液調(diào)節(jié)pH值至6.8,加入100 mg樟芝多糖粉末,再加入20 mgα-淀粉酶使其終濃度為8 U/mL。37 ℃水浴反應(yīng)150 min,每30 min取樣測(cè)定釋放的還原糖含量,并計(jì)算水解度。還原糖含量采用DNS法測(cè)定,初始總糖含量(即最初還原糖含量)采用苯酚-硫酸法測(cè)定,水解度按照下式計(jì)算。
1.3.6.2 抗模擬胃液消化能力的測(cè)定
抗模擬胃液消化能力的測(cè)定參考文獻(xiàn)[19]。首先,配制人工模擬胃液。稱取胃蛋白酶3.0 g,用300 mL生理鹽水溶解,等體積分為3 份,用鹽酸溶液(2 mol/L)分別調(diào)節(jié)pH值至2.0、3.0和4.0,再用微孔濾膜(0.22 μm)過(guò)濾除菌備用。隨后,各取10 mL上述不同pH值的模擬胃液,加入100 mg樟芝多糖粉末,37 ℃水浴反應(yīng)180 min,每30 min取樣測(cè)定釋放的還原糖含量,按照上式計(jì)算水解度。
1.3.7 小鼠實(shí)驗(yàn)
1.3.7.1 小鼠造模
設(shè)置3 個(gè)LIH灌胃劑量,分別為2.7、3.7、4.2 g/kgmb,進(jìn)行連續(xù)3 日灌胃造模,每日灌胃兩次(上午9點(diǎn)和下午7點(diǎn)各一次)。停止給藥后,觀察小鼠在隨后1~4 d的毛發(fā)、飲食、活動(dòng)及糞便形態(tài)等指標(biāo),以確定小鼠造模所用LIH的最適給藥劑量。
1.3.7.2 小鼠分組及處理
將30 只實(shí)驗(yàn)小鼠隨機(jī)分為5 組(n=6):正常組(NC組)、模型組(RG組)、樟芝胞外多糖處理組(AEPS組)、樟芝胞內(nèi)多糖處理組(AIPS組)及低聚果糖陽(yáng)性對(duì)照組(FOS組)。隨后,在1~3 d,NC組按3.7 g/kgmb的劑量灌胃無(wú)菌生理鹽水;RG組、AEPS組、AIPS組及FOS組按3.7 g/kgmb的劑量灌胃LIH。各組每日均灌胃兩次(上午9點(diǎn)和下午7點(diǎn)各一次);接下來(lái)的4~6 d,NC組和RG組每日下午7點(diǎn)按0.03 g/kgmb的劑量灌胃無(wú)菌生理鹽水,AEPS組按0.03 g/kgmb的劑量灌胃AEPS,AIPS組按0.03 g/kgmb的劑量灌胃AIPS,F(xiàn)OS組按0.03 g/kgmb的劑量灌胃低聚果糖。
1.3.7.3 小鼠糞便16S rDNA測(cè)序
第7天處死小鼠并采集小鼠糞便樣本,液氮速凍后置于干冰中寄送至北京諾禾致源科技股份有限公司進(jìn)行16S rDNA測(cè)序。采用Illumina NovaSeq6000測(cè)序平臺(tái)。使用正向引物(5’-CCTACGGGNGGCWGCAG-3’)和反向引物(5’-GGACTACHVGGGTATCTAAT-3’)通過(guò)聚合酶鏈?zhǔn)椒磻?yīng)(polymerase chain reaction,PCR)擴(kuò)增16S rDNA基因的V3~V4區(qū)域。初始DNA片段用Flash軟件進(jìn)行讀取,使用QIIM軟件對(duì)拼接數(shù)據(jù)進(jìn)行質(zhì)量控制,然后對(duì)所有樣本的有效Tags以97%的一致性(Identity)進(jìn)行操作分類單元(operational taxonomic units,OTU)聚類和物種分類分析。
1.4 數(shù)據(jù)處理與分析
每組實(shí)驗(yàn)及檢測(cè)均設(shè)置至少3 個(gè)重復(fù)。所有數(shù)據(jù)均以平均值±標(biāo)準(zhǔn)差的形式表示,采用Origin 2021軟件繪圖。
2 結(jié)果與分析
2.1 樟芝多糖的結(jié)構(gòu)分析
用Sevag法經(jīng)8 次脫蛋白處理后,檢測(cè)樟芝多糖組成情況。由表1可知,AEPS和AIPS的總中性糖質(zhì)量分?jǐn)?shù)分別為82%和88%,蛋白質(zhì)量分?jǐn)?shù)均小于1.5%,純度已足以用于小鼠實(shí)驗(yàn)[20]。此外,AEPS和AIPS的主要組成單糖均為葡萄糖、半乳糖及甘露糖,但含量差異較大。其中,在葡萄糖相對(duì)含量方面,AEPS為84.73%,而AIPS為55.31%;在半乳糖相對(duì)含量方面,AEPS為7.84%,AIPS卻高達(dá)28.52%;在甘露糖相對(duì)含量方面,AEPS為5.27%,AIPS高達(dá)14.34%。此外,AEPS中還含有葡萄糖醛酸和半乳糖醛酸,相對(duì)含量分別為0.76%和1.4%,而AIPS中則含有1.83%的氨基半乳糖。
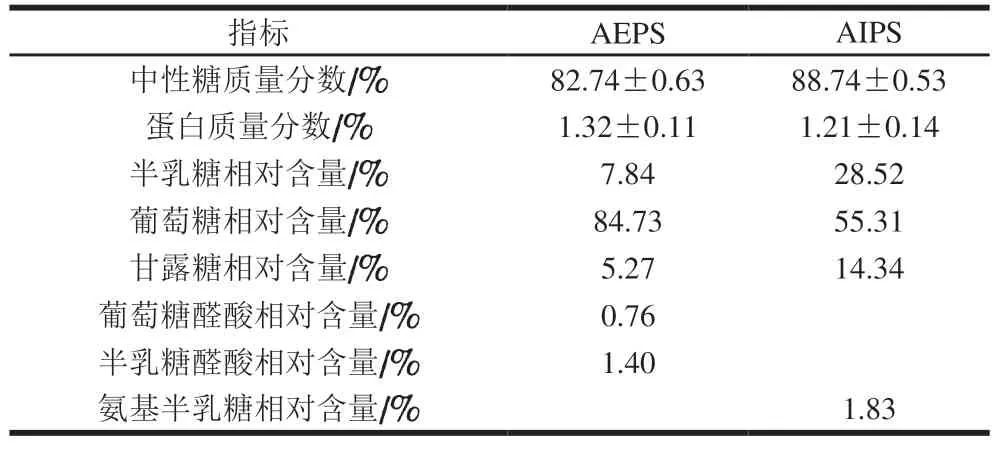
表1 AEPS及AIPS的組成測(cè)定結(jié)果Table 1 Monosaccharide composition of AEPS and AIPS
2.2 樟芝多糖的分子質(zhì)量分布
樟芝多糖的分子質(zhì)量分布結(jié)果如圖1所示,AEPS主要由3 種不同分子質(zhì)量的多糖組成,其中分子質(zhì)量為1 013 kDa的多糖占比為77.96%,分子質(zhì)量為233 kDa的多糖占比為18.04%,分子質(zhì)量為28 743 kDa的多糖占比為3.99%。AEPS的平均分子質(zhì)量為4.16×105Da;AIPS主要由2 種不同分子質(zhì)量的多糖組成,其中分子質(zhì)量為12 452 kDa的多糖占比為49.54%,分子質(zhì)量為1 623 kDa的多糖占比為50.46%。AIPS的平均分子質(zhì)量為3.52×106Da。可見(jiàn),AEPS和AIPS在多糖組成上亦有著明顯差異。
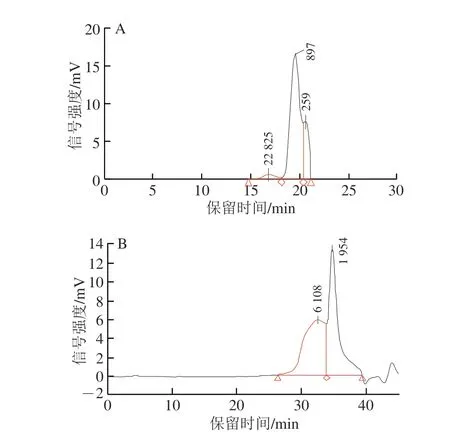
圖1 AEPS(A)和AIPS(B)的分子質(zhì)量分布Fig.1 Molecular mass distribution of AEPS (A) and AIPS (B)
2.3 樟芝多糖的紅外光譜分析
樟芝多糖紅外光譜分析結(jié)果如圖2 所示,在4 000~400 cm-1范圍內(nèi),AEPS在3 400 cm-1左右有強(qiáng)而寬的吸收峰,這主要是由多糖中羥基的伸縮振動(dòng)引起的;在1 673 cm-1左右有強(qiáng)而寬的吸收峰,這主要是由羰基(C=O)的伸縮引起的,表明AEPS具有酰胺結(jié)構(gòu);在1 100 cm-1處有強(qiáng)而寬的吸收峰,這主要是由吡喃環(huán)結(jié)構(gòu)的C—O鍵引起的,說(shuō)明AEPS具有吡喃糖環(huán)[21];在944 cm-1處有強(qiáng)而寬的吸收峰,說(shuō)明AEPS存在β-型糖苷鍵(圖2A)。AIPS在3 276 cm-1左右有強(qiáng)而寬的吸收峰,這主要是由多糖羥基中分子間的氫鍵伸縮振動(dòng)引起的[22];在3 000 cm-1左右有吸收峰,這主要是由—CH—鍵引起的,表明AIPS具有—C≡C—H結(jié)構(gòu);在1 642 cm-1左右存在強(qiáng)吸收峰,表明A I P S 存在酰胺結(jié)構(gòu);在1 074 cm-1和1 047 cm-1處存在強(qiáng)吸收峰,表明AIPS存在C—O結(jié)構(gòu)(圖2B)。
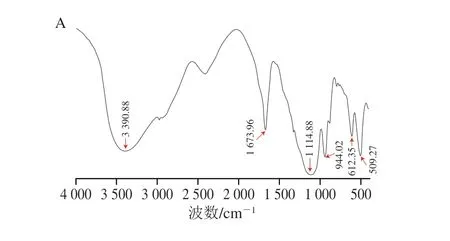
圖2 AEPS(A)和AIPS(B)傅里葉變換紅外光譜Fig.2 Fourier transform infrared spectra of AEPS (A) and AIPS (B)
2.4 樟芝多糖的體外抗消化能力
建立體外消化模型,對(duì)樟芝多糖的抗消化能力進(jìn)行測(cè)定,結(jié)果如圖3所示。AEPS在α-淀粉酶的作用下,水解度隨著時(shí)間的延長(zhǎng)而增加,在150 min后水解度變化不明顯,約為0.02%(圖3A)。AEPS在不同pH值的胃模擬液中水解度也較低,在不同pH值條件下的水解度都會(huì)隨著時(shí)間的延長(zhǎng)而增加(圖3B),其中在pH 2.0條件下水解度最高,210 min時(shí)約為0.05%,AEPS在pH 3.0胃模擬液中的水解度高于pH 4.0胃模擬液中的水解度。此外,AIPS在α-淀粉酶的作用下,180 min時(shí)的水解度為0.028%(圖3C)。AIPS在不同pH值的胃模擬液中水解度較AEPS高,AIPS同樣也在pH 2.0的胃模擬液中水解度最高,210 min時(shí)為0.064%(圖3D)。上述結(jié)果表明,AEPS及AIPS均具有較強(qiáng)的抗消化能力,即若口服樟芝多糖,其可順利進(jìn)入腸道并發(fā)揮作用。
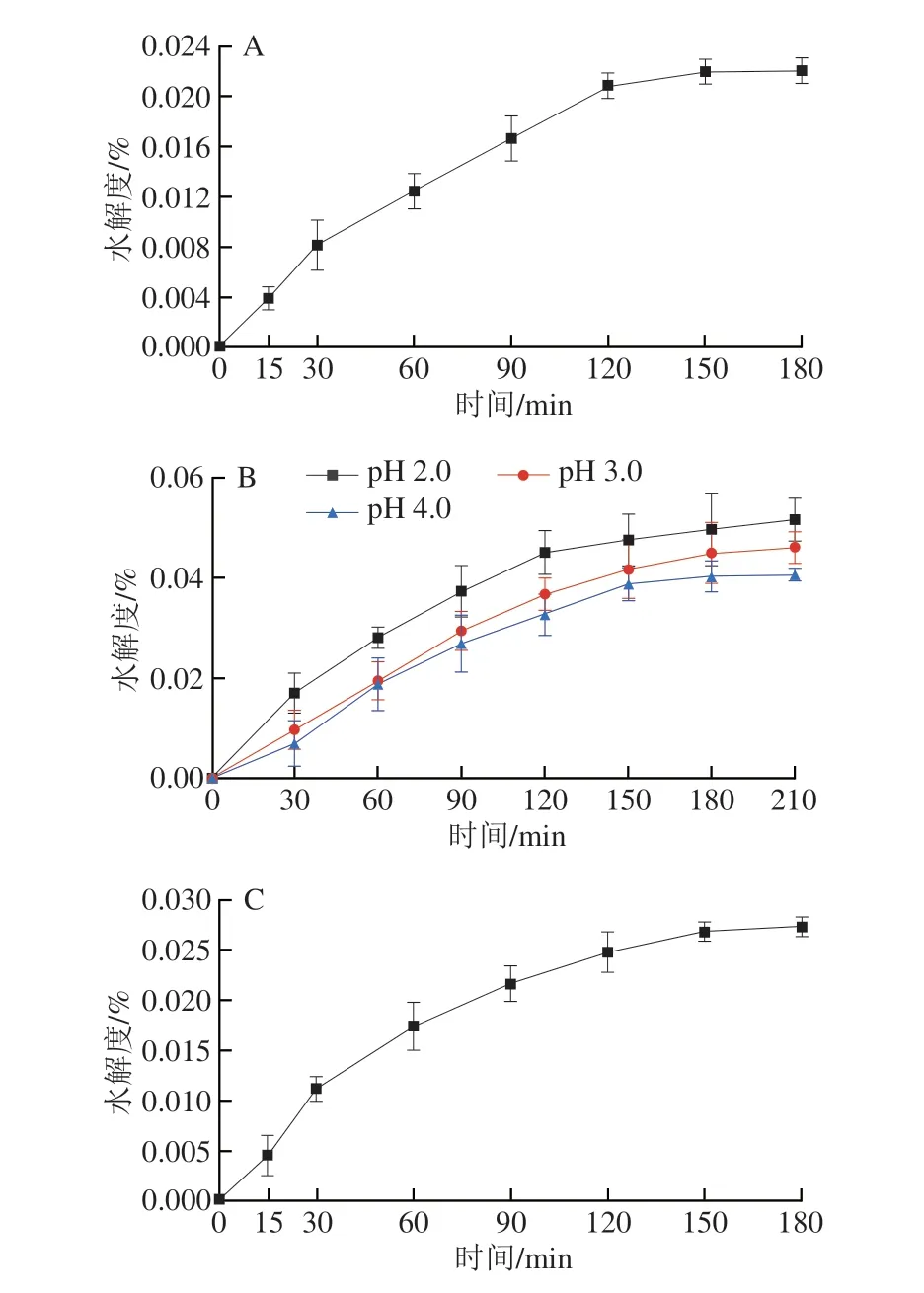
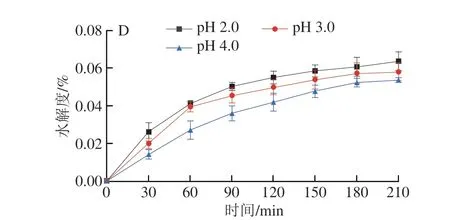
圖3 樟芝多糖的體外抗消化能力Fig.3 In vitro digestive resistance of polysaccharides from Antrodia camphorata
2.5 樟芝多糖對(duì)抗生素相關(guān)性腹瀉小鼠腸道菌群的調(diào)節(jié)作用
2.5.1 不同劑量LIH對(duì)小鼠狀態(tài)的影響
如圖4所示,不同LIH劑量組小鼠停藥后4 d內(nèi)的狀態(tài)和表現(xiàn)差異較為明顯。灌胃2.7 g/kgmbLIH的小鼠毛發(fā)狀態(tài)與正常對(duì)照組相比沒(méi)有明顯的變化,均較為平滑;此外,小鼠活動(dòng)正常,攝食量與正常對(duì)照組相比無(wú)明顯差異。灌胃3.7 g/kgmbLIH的小鼠出現(xiàn)輕微的脫毛現(xiàn)象,并且活動(dòng)遲緩、毛發(fā)粗糙、食欲減退,攝食量與正常對(duì)照組相比明顯減少;灌胃4.2 g/kgmbLIH的小鼠出現(xiàn)多處大面積的脫毛現(xiàn)象,行為活動(dòng)遲緩、煩躁少動(dòng)、食欲不振,幾乎不攝食。此外,從小鼠糞便形態(tài)來(lái)看,灌胃2.7 g/kgmbLIH的小鼠出現(xiàn)輕微腹瀉,并在停藥1 d后即完全恢復(fù)正常;灌胃3.7 g/kgmbLIH的小鼠糞便稀軟、不成形,呈爛泥狀,表現(xiàn)為中度腹瀉,并在停藥4 d后恢復(fù)正常;灌胃4.2 g/kgmbLIH的小鼠糞便稀溏,稀薄如水,呈現(xiàn)水樣便,表現(xiàn)為嚴(yán)重腹瀉,并在停藥4 d后仍持續(xù)腹瀉。綜上所述,最終選擇3.7 g/kgmb作為后續(xù)小鼠建模的LIH給藥劑量。

圖4 不同劑量LIH下小鼠停藥2~4 d的狀態(tài)Fig.4 Conditions of mice at 2–4 days after LIH withdrawal
2.5.2 AEPS及AIPS對(duì)小鼠腸道菌群的影響
2.5.2.1 腸道菌群多樣性分析
為了解各組小鼠體內(nèi)腸道菌群的物種豐富度和均一性,首先對(duì)小鼠腸道菌群測(cè)序結(jié)果進(jìn)行α-多樣性分析,結(jié)果如圖5所示。RG組小鼠腸道菌群的多樣性明顯低于NC組,說(shuō)明LIH嚴(yán)重破壞了小鼠體內(nèi)的腸道菌群平衡,并抑制或殺死了大量微生物;同時(shí),灌胃AEPS、AIPS及FOS(低聚果糖)均有助于調(diào)節(jié)和恢復(fù)小鼠腸道菌群的多樣性,且AEPS的效果優(yōu)于AIPS和FOS。
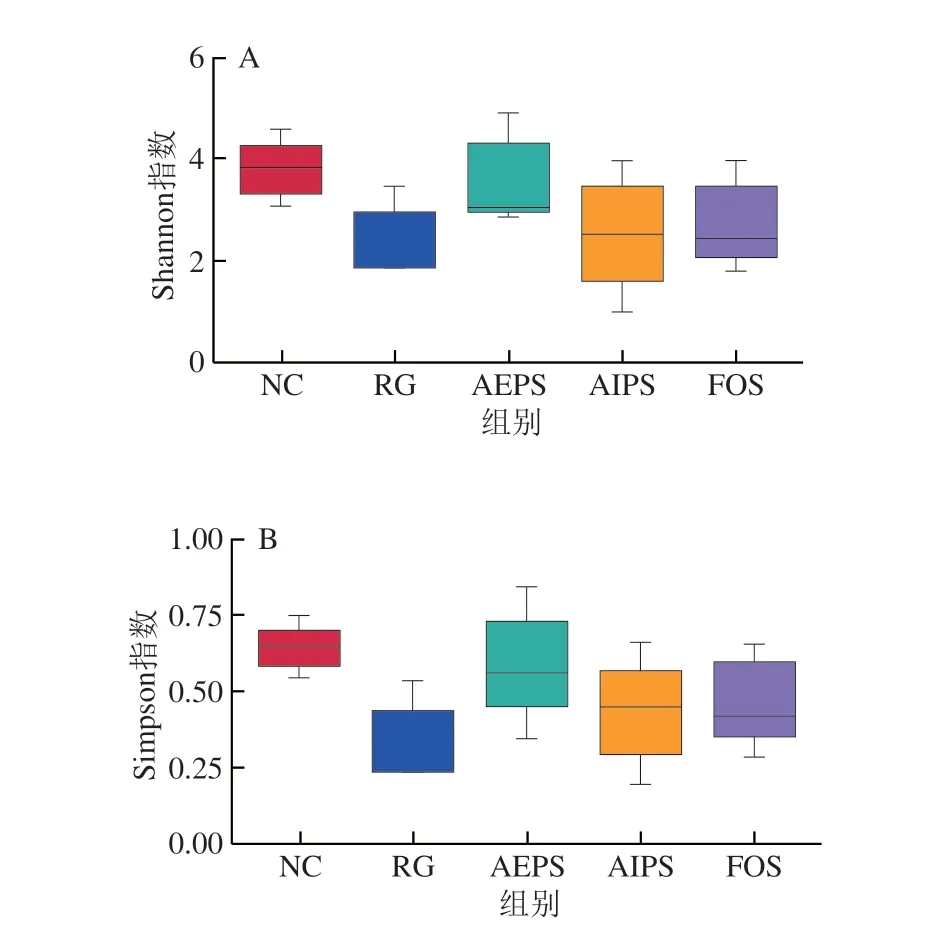
圖5 腸道菌群α-多樣性分析的Shannon指數(shù)(A)和Simpson指數(shù)(B)Fig.5 Shannon (A) and Simpson (B) indexes of α-diversity analysis of intestinal microbiota
為了比較不同處理組間小鼠腸道菌群結(jié)構(gòu)的差異,對(duì)測(cè)序結(jié)果進(jìn)行了β-多樣性分析(主坐標(biāo)分析(principal co-ordinates analysis,PCoA)和非度量多維尺度(non-metric multidimensional scaling,NMDS)分析)(圖6)。結(jié)果表明,不同處理組之間的小鼠腸道菌群結(jié)構(gòu)均具有明顯差異,說(shuō)明灌胃LIH、AEPS、AIPS及FOS均對(duì)小鼠腸道菌群結(jié)構(gòu)產(chǎn)生了較為明顯的影響,也說(shuō)明樟芝多糖對(duì)小鼠腸道菌群多樣性具有明顯的調(diào)節(jié)作用。此外,從各組中重復(fù)樣本間的距離來(lái)看,各組重復(fù)樣本均較為集中,說(shuō)明各重復(fù)樣本的差異較小,重復(fù)性較好。
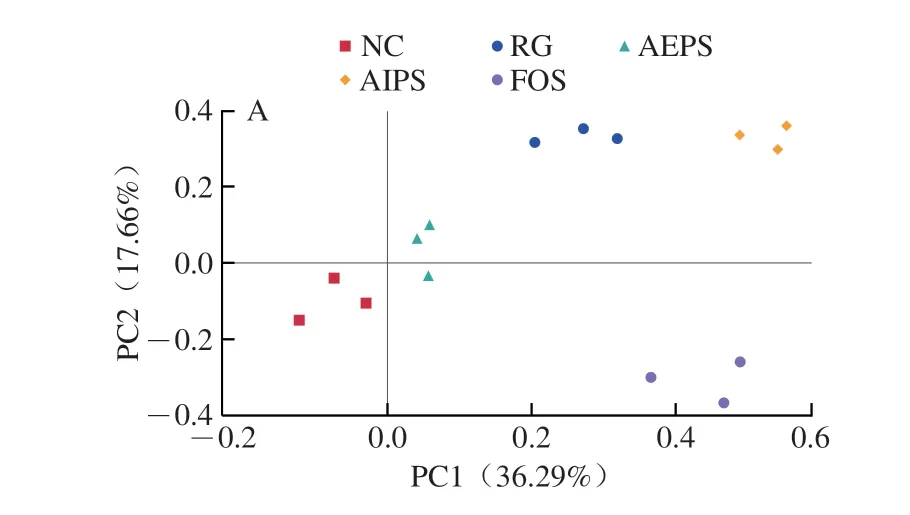
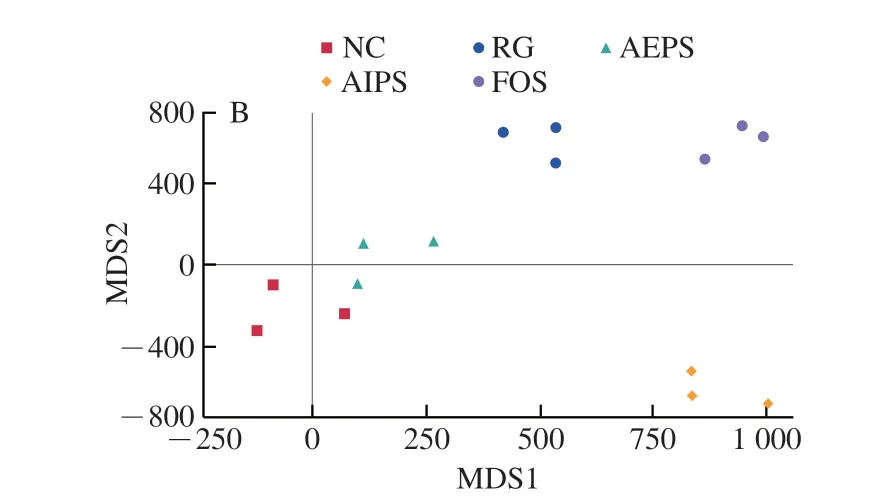
圖6 腸道菌群結(jié)構(gòu)的PCoA(A)和NMDS分析(B)Fig.6 Principal co-ordinates analysis (A) and non-metric multidimensional scaling analysis (B) of intestinal flora structure
2.5.2.2 腸道菌群結(jié)構(gòu)分析
為直觀查看各組小鼠腸道菌群在不同分類水平上相對(duì)豐度較高的物種及其比例,根據(jù)各樣本在屬水平上的物種注釋和豐度信息,分別繪制了各組小鼠腸道菌群在門水平、科水平及屬水平上豐度排名前10的微生物相對(duì)豐度柱形圖。結(jié)果表明,從門水平來(lái)看,厚壁菌門(Firmicutes)、擬桿菌門(Bacteroidota)和變形菌門(Proteobacteria)為主要的優(yōu)勢(shì)菌群,所占的比例約為總菌群的95%(圖7A)。此外,在變形菌門的占比上,NC組明顯低于RG、AEPS、AIPS及FOS組,但在擬桿菌門的占比上,NC組明顯高于RG、AEPS、AIPS及FOS組;厚壁菌門在各處理組小鼠腸道中的相對(duì)豐度較低,但與NC組相比,AEPS組中厚壁菌門的相對(duì)豐度大幅增加(圖8A)。
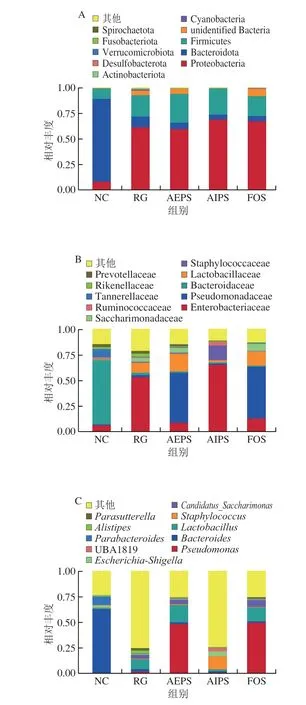
圖7 各組小鼠腸道菌群在門水平(A)、科水平(B)及屬水平(C)的相對(duì)豐度Fig.7 Relative abundance of intestinal flora in mice from different groups at the phylum (A),family (B),and genus (C) levels
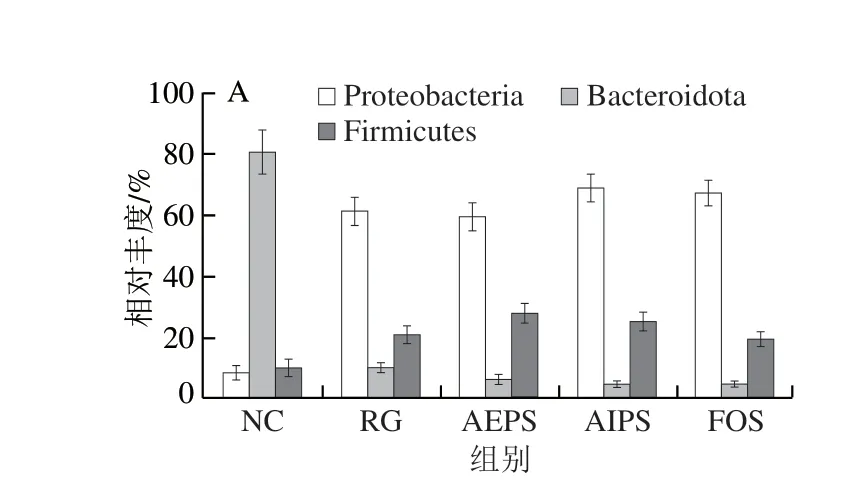
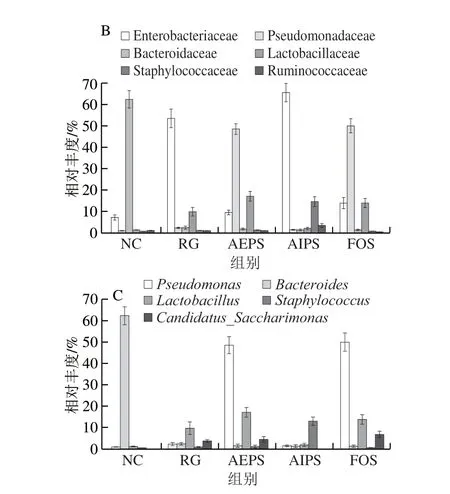
圖8 各組小鼠腸道菌群在不同水平上的相對(duì)豐度Fig.8 Relative abundance of intestinal bacteria in different groups at different levels
從科水平來(lái)看,葡萄球菌科(Staphylococcaceae)、乳酸桿菌科(Lactobacillaceae)、擬桿菌科(Bacteroidaceae)、假單胞菌科(Pseudomonadaceae)、腸桿菌科(Enterobacteriaceae)及疣微菌科(Ruminococcaceae)為主要的優(yōu)勢(shì)菌群,所占的比例約為總菌群的80%(圖7B)。乳酸桿菌是益生菌群,AEPS組中乳酸桿菌科的相對(duì)豐度明顯高于NC、RG及AIPS組(圖8B),表明灌胃AEPS可以大幅提升小鼠腸道內(nèi)乳酸桿菌科的相對(duì)豐度;腸桿菌科在RG及AIPS組中的相對(duì)豐度明顯高于NC、AEPS及FOS組。從圖8B中可看出,灌胃AIPS后,小鼠腸道內(nèi)Staphylococcaceae的相對(duì)豐度較NC及RG組急劇上升,這一現(xiàn)象在AEPS及FOS組中均未出現(xiàn),而Staphylococcaceae為有害菌[23]。此外,AIPS組中Ruminococcaceae的相對(duì)豐度較其他組明顯增加,而Ruminococcaceae亦為有害菌[24]。
從屬水平來(lái)看,假絲酵母菌屬(Candidatus_Saccharimonas)、葡萄球菌屬(Staphylococcus)、乳酸菌屬(Lactobacillus)、擬桿菌屬(Bacteroides)及假單胞菌屬(Pseudomonas)為主要優(yōu)勢(shì)菌群(圖7C、8C)。
為了進(jìn)一步分析不同組間小鼠腸道菌群相對(duì)豐度差異較大的微生物物種,選取微生物相對(duì)豐度排名前35 的屬,根據(jù)其在每個(gè)組中的豐度信息,從物種和樣本兩個(gè)層面進(jìn)行聚類,繪制成熱圖。如圖9 所示,另枝菌屬(Alistipes)、擬桿菌屬(Bacteroides)、副桿菌屬(Parabacteroides)、丹毒桿菌屬(Erysipelatoclostridium)、普拉梭菌屬(Faecalibacterium)、腸球菌屬(Enterococcus)、葡萄球菌屬(Staphylococcus)及乳酸菌屬(Lactobacillus)等為主要菌群,且在不同組中的相對(duì)豐度差異明顯。
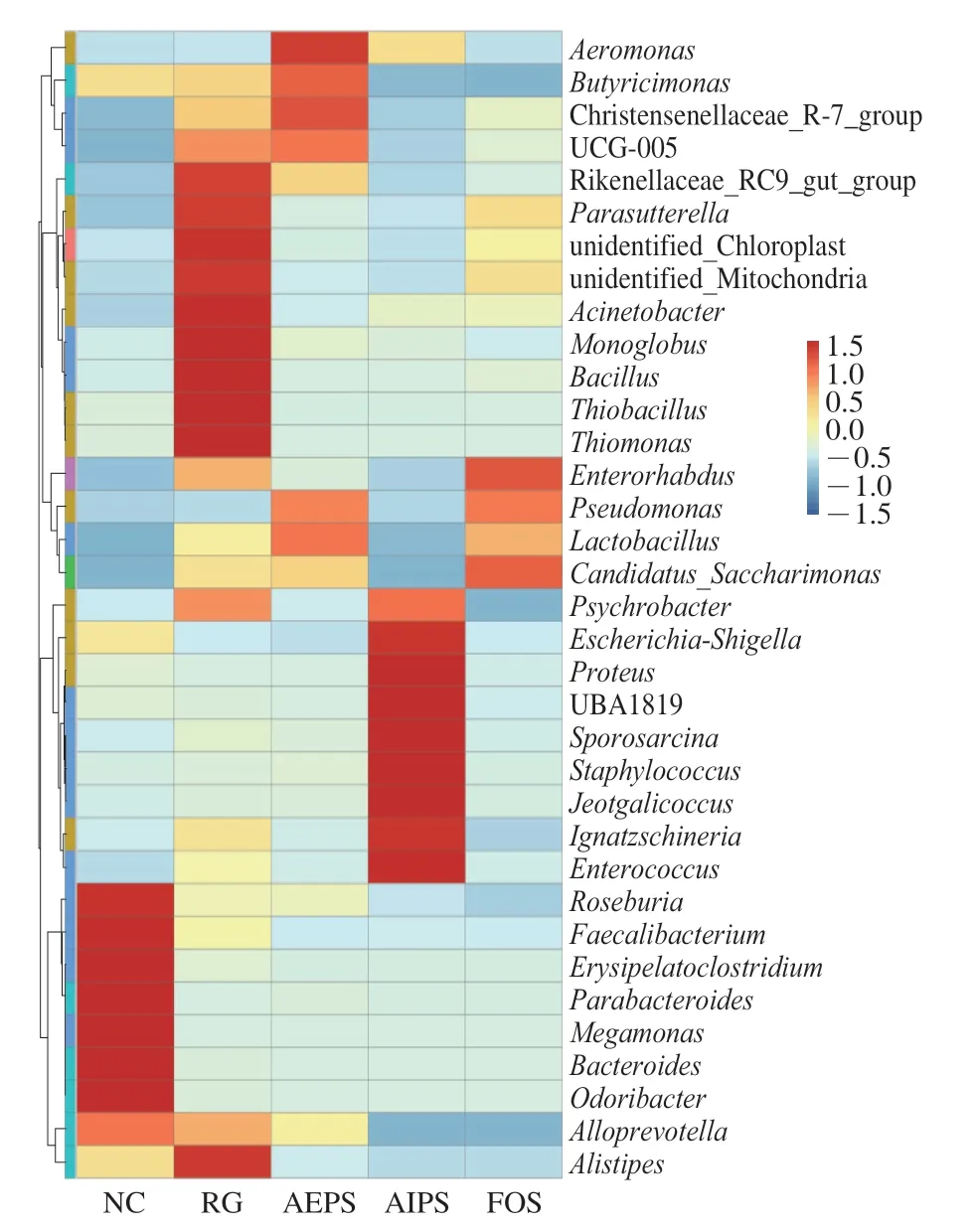
圖9 各組小鼠腸道菌群在屬水平的相對(duì)豐度Fig.9 Relative abundance of intestinal flora at the genus level in mice from different groups
此外,從圖9中可以看出,與NC、AEPS、AIPS及FOS組相比,RG組小鼠腸道中Parasutterella、Thiomonas、Thiobacillus、Bacillus等菌屬的相對(duì)豐度明顯增加。與AEPS及FOS組相比,AIPS組小鼠腸道菌群中Enterococcus及Staphylococcus的相對(duì)豐度明顯增加,而Enterococcus和Staphylococcus的相對(duì)豐度與炎癥程度呈正相關(guān)[25]。此外,在AEPS和FOS組中,Lactobacillus的相對(duì)豐度較RG及AIPS組明顯增高,而Lactobacillus對(duì)腸道疾病有治療作用,能減輕高濃度抗生素引起的腹瀉癥狀[26]。
3 討論
多數(shù)真菌多糖具有與益生元相似的功能,能通過(guò)刺激特定腸道菌群的生長(zhǎng)或活性而使宿主受益。這是因?yàn)榇蠖鄶?shù)真菌多糖屬于β-葡聚糖類,對(duì)胃及腸道中的消化酶具有較強(qiáng)的抵抗性,通常是不可被消化的[27]。Nowak等[28]提取了53 種真菌子實(shí)體多糖,同時(shí)測(cè)定了這些多糖對(duì)嗜酸乳桿菌(Lactobacillus acidophilus)和兩株鼠李糖乳桿菌(Lactobacillus rhamnosus)生長(zhǎng)的影響,并通過(guò)人工胃液對(duì)真菌多糖的體外抗消化能力進(jìn)行了研究。結(jié)果發(fā)現(xiàn),真菌多糖比菊粉或FOS等益生元對(duì)乳酸桿菌生長(zhǎng)的促進(jìn)效果更為明顯;檢測(cè)這些多糖的體外消化率得知,真菌多糖經(jīng)人工胃液處理后,90%以上未被消化,表明這些真菌多糖具有良好的抗胃液消化能力。本研究中體外消化模擬實(shí)驗(yàn)結(jié)果表明,AEPS及AIPS均呈現(xiàn)出較強(qiáng)的抗消化能力。
真菌多糖相比較菊粉、低聚果糖等益生元更能促進(jìn)有益菌(如雙歧桿菌和乳酸桿菌等)的生長(zhǎng)[28]。Zhang Rongjun等[29]研究發(fā)現(xiàn),金針菇多糖可通過(guò)調(diào)節(jié)腸道菌群導(dǎo)致的Toll樣受體4(Toll-like receptor 4,TLR4)/核因子(nuclear factor,NF)-κB炎癥信號(hào)通路下調(diào),最終改善了葡聚糖硫酸鈉(dextran sulfate sodium,DSS)引發(fā)的結(jié)腸炎癥狀;Ren Yilin等[30]研究發(fā)現(xiàn),猴頭菇多糖可下調(diào)氧化應(yīng)激和炎癥因子標(biāo)志物,并逆轉(zhuǎn)DSS誘導(dǎo)的炎癥C57BL/6小鼠腸道菌群紊亂,維持腸道屏障的穩(wěn)定性。而樟芝作為一種新興的珍貴藥食兩用真菌,其多糖具有抗氧化、抗炎癥、抗病毒及調(diào)節(jié)免疫等多種生物活性,在益生元方面亦具有較大的研究?jī)r(jià)值及開(kāi)發(fā)潛力[31]。
本研究結(jié)果表明,AEPS和FOS的攝入會(huì)顯著增加小鼠腸道中Lactobacillus相對(duì)豐度(P<0.05),且AEPS組中志賀氏菌屬(Shigella)相對(duì)豐度明顯低于AIPS組(圖7C)。益生菌乳酸菌屬(Lactobacillus)是人類胃腸道中最豐富的微生物之一,與良好的腸道健康有關(guān)。研究表明,益生菌能夠治療和預(yù)防多種腸道疾病,包括Clostridium-difficile誘導(dǎo)的結(jié)腸炎[32],Kanwal等[33]通過(guò)灌胃菱形藻多糖提升了炎癥小鼠腸道中Lactobacillus的相對(duì)豐度,同時(shí)降低了Proteobacteria、Gammaproteobacteria和Bacteroides的相對(duì)豐度,從而改善了DSS誘導(dǎo)的炎癥反應(yīng)。Hempel等[34]研究證明了Lactobacillus和Bifidobacterium能夠治療抗生素引起腹瀉的癥狀。Friedman[26]認(rèn)為,益生菌Lactobacillus可以通過(guò)分泌黏蛋白增加上皮細(xì)胞緊密連接,提供定植抗性,產(chǎn)生細(xì)菌素,促進(jìn)免疫球蛋白A的分泌,產(chǎn)生平衡T輔助細(xì)胞反應(yīng),增加IL-10的含量,從而增強(qiáng)腸道黏膜屏障功能,有助于抗生素?fù)p傷后正常腸道平衡的恢復(fù)。Shigella能夠侵入人類腸黏膜,從而導(dǎo)致細(xì)菌性痢疾(一種急性直腸結(jié)腸炎)[35]。
在AIPS組中,Staphylococcus的相對(duì)豐度明顯高于AEPS、FOS及RG組。而Staphylococcus是醫(yī)院感染和術(shù)后感染中常見(jiàn)的微生物,它被認(rèn)為是引發(fā)抗生素相關(guān)性腹瀉最常見(jiàn)的原因之一[25]。Bendali等[36]發(fā)現(xiàn),健康兔子灌胃Staphylococcus誘發(fā)持續(xù)性腹瀉發(fā)癥狀后,灌胃Lactobacillus paracasei可使腹瀉停止并使腸道絨毛恢復(fù),且能夠明顯降低Staphylococcus在糞便中的相對(duì)豐度。Liu Guanwen等[23]發(fā)現(xiàn),Lactobacillus rhamnosus在體外和體內(nèi)均能有效抑制具有多重耐藥性的Staphylococcus,具體表現(xiàn)為有效降低Staphylococcus的相對(duì)豐度,抑制炎癥因子腫瘤壞死因子(tumor necrosis factor,TNF)-α和白細(xì)胞介素(interleukin,IL)-6的表達(dá),恢復(fù)血液中白細(xì)胞和中性粒細(xì)胞的水平,有效修復(fù)因Staphylococcus感染而受損的腸道屏障和恢復(fù)免疫器官的結(jié)構(gòu)損傷。從圖7C可以看出,AEPS組中Lactobacillus的相對(duì)豐度明顯高于RG組(P<0.05),說(shuō)明AEPS可明顯降低Staphylococcus的相對(duì)豐度,從而緩解腹瀉癥狀及腸道損傷。
從圖9可以看出,Parasutterella在RG組中的相對(duì)豐度最高。Parasutterella是來(lái)自變形桿菌門的一種革蘭氏陰性、嚴(yán)格厭氧的球菌屬。有報(bào)道稱,腸道內(nèi)Parasutterella相對(duì)豐度的增加會(huì)導(dǎo)致胃腸道生態(tài)失調(diào)及腸道菌群多樣性的降低,進(jìn)而引發(fā)腸道疾病或代謝疾病,如炎癥性腸病和肥胖[37]。同時(shí),在AIPS組中,Enterococcus的相對(duì)豐度較高,而Onderdonk等[38]發(fā)現(xiàn),大鼠結(jié)腸炎與Enterococcus豐度之間存在正相關(guān)性。
4 結(jié)論
AEPS及AIPS經(jīng)過(guò)8 次Sevag法脫蛋白處理后,蛋白質(zhì)量分?jǐn)?shù)均低于1.5%,總中性糖質(zhì)量分?jǐn)?shù)分別約為83%及89%,純度較高;采用HPLC法分析樟芝多糖(AEPS及AIPS)的單糖組成,結(jié)果表明AEPS主要由半乳糖、葡萄糖、甘露糖、葡萄糖醛酸、半乳糖醛酸組成,相對(duì)含量分別為7.84%、84.73%、5.27%、0.76%及1.40%;而AIPS主要由半乳糖、葡萄糖、甘露糖、氨基半乳糖組成,相對(duì)含量分別為28.52%、55.31%、14.34%及1.83%;采用HPGFC法分析樟芝多糖的分子質(zhì)量分布,發(fā)現(xiàn)AEPS的平均分子質(zhì)量為4.16×105Da,AIPS的平均分子質(zhì)量為3.52×106Da,二者之間存在明顯差異;分析樟芝多糖在4 000~400 cm-1范圍內(nèi)紅外光譜發(fā)現(xiàn),AEPS具有吡喃糖環(huán),且存在β-型糖苷鍵;而AIPS具有酰胺結(jié)構(gòu),且存在(—C≡C—H)、C—O官能團(tuán);體外模擬消化實(shí)驗(yàn)結(jié)果表明,AEPS及AIPS均具有較強(qiáng)的抗消化能,表明口服樟芝多糖后,其可順利進(jìn)入腸道并發(fā)揮作用;小鼠體內(nèi)實(shí)驗(yàn)結(jié)果表明,灌胃AEPS可大幅增加小鼠腸道菌群中的部分有益微生物(如Lactobacillus)的相對(duì)豐度,同時(shí)明顯降低小鼠腸道菌群中部分有害微生物(如Enterococcus、Staphylococcus、Parasutterella及Shigella)的相對(duì)豐度。其中,AEPS對(duì)小鼠腸道菌群的調(diào)節(jié)效果明顯優(yōu)于AIPS。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級(jí))(2025年8期)2025-08-18 00:00:00
東方少年·布老虎畫刊(2023年8期)2023-08-01 15:45:12
科學(xué)大眾(2021年6期)2021-07-20 07:42:44
科學(xué)(2020年3期)2020-11-26 08:18:30
學(xué)苑創(chuàng)造·A版(2020年9期)2020-10-13 09:41:02
娃娃樂(lè)園·綜合智能(2019年3期)2019-04-03 09:17:36
中成藥(2018年2期)2018-05-09 07:19:34
小學(xué)生學(xué)習(xí)指導(dǎo)(低年級(jí))(2017年10期)2017-10-10 01:00:05
湖北師范大學(xué)學(xué)報(bào)(自然科學(xué)版)(2015年2期)2016-01-10 08:41:55
云南中醫(yī)學(xué)院學(xué)報(bào)(2014年3期)2014-07-31 18:57:34
