果酒增香釀造技術研究進展
2023-08-12 00:41:50王曉宇趙擎豪趙越凡杜國榮趙鵬濤
食品科學 2023年13期
王 飛,王曉宇,2,3,趙擎豪,趙越凡,劉 炎,杜國榮,趙鵬濤,2,3,*
(1.陜西師范大學食品工程與營養科學學院,陜西 西安 710119;2.國家蘋果加工技術研發專業中心,陜西 西安 710119;3.西部果品高值利用教育部工程研究中心,陜西 西安 710119;4.西安文理學院生物與環境工程學院,陜西 西安 710065)
果酒因其高營養價值和良好的口感深受消費者青睞。然而,多數果酒存在香氣寡淡、風味典型性不足的問題,極大影響了果酒的感官品質,制約著我國果酒產業的進一步發展。根據香氣產生的釀造階段,果酒香氣可以分為品種香、發酵香和陳釀香。微生物在果酒釀造的各個階段都發揮著重要作用,其生長代謝、產酶類型和產酶活力以及菌種間相互作用均對果酒風味形成有較大影響[1-3]。因此,深入解析釀酒微生物、酶和發酵前冷浸漬等工藝處理對果酒香氣的影響機制,創制基于微生物香氣化合物調控技術,對于提高果酒香氣質量具有重要意義。
本文通過剖析釀酒微生物、酶及發酵前冷浸漬等工藝處理對果酒香氣化合物生成的影響,闡明主要香氣化合物的生成機理,并探討優良產香酵母選育的研究進展。最后,對未來果酒風味的研究方向進行展望,旨在為果酒增香釀造技術的創制提供一定理論支持。
1 果酒香氣
香氣是果酒最重要的感官品質指標。果酒中發現的香氣物質超過1 000 種,主要是在微生物生長代謝過程中形成,其產生機制如圖1所示。來源于水果的品種香是果酒香氣的重要組成部分,有利于果酒典型風味的塑造,然而大多品種香氣化合物在釀酒及貯藏過程中會發生一系列變化,果香特征減弱[4];乙醇發酵過程產生的發酵香氣構成了果酒香氣的基本骨架,對果酒最終風味形成具有決定作用;陳釀過程中品種香氣與發酵香氣進一步轉化,使果酒整體香氣更加平衡和諧。
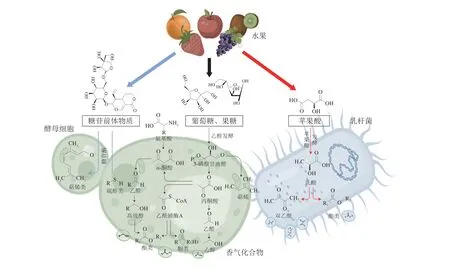
圖1 果酒微生物產香機制Fig.1 Mechanism of microbial aroma production in fruit wine
1.1 果酒香氣形成機制
在發酵過程中,以糖苷結合態形式存在于水果中的非揮發性香氣前體物質在釀酒酵母相關酶的作用下解離出游離態香氣,主要包括一些硫醇類和萜烯類化合物,這些物質貢獻了果酒主要的品種香氣,賦予果酒獨特的風味。
總體來說,果酒芳香大部分由發酵香氣組成,構成了果酒香氣的主體部分,香氣化合物包括酯類、醇類、有機酸類、醛酮類、含硫化合物以及萜烯類等[5]。乙醇發酵過程中由釀酒酵母代謝產生的香氣化合物貢獻了果酒的主要發酵香氣。糖酵解產生一些直接或次級代謝物如甘油、乙醇等,構成了果酒的基本香氣骨架。氨基酸代謝產生的風味物質主要是高級醇以及與其相關的酯類物質,還包括少量的醛類物質。乙醇發酵結束后,經過蘋果酸-乳酸發酵,在降酸的同時生成雙乙酰和一些酯類化合物,有助于果酒風味的進一步提升。
陳釀過程中果酒的芳香成分遷移轉化,進一步提升了果酒的香氣復雜性。關于果酒的陳釀增香機理,一般認為與氧有關:一是“氧化說”,如葡萄酒陳釀期間氧氣通過橡木桶的微孔進入酒液,使醇、醛類物質緩慢氧化,高級醇類含量降低;二是“酯化說”,即醇類物質氧化成相應的酸,酸與乙醇反應形成酯,使酯香增強。陳釀香氣會隨陳釀周期的變化呈現不同的結果。蒲彪等[6]研究了枇杷果酒陳釀期間香氣成分的變化規律,發現醇類含量總體減少,酯類含量總體增加,酯香復雜濃郁,且陳釀3 個月后特征香氣由辛酸乙酯更替為癸酸乙酯。基于此,果酒陳釀增香的研究方向應為促進醇的氧化和酯的形成。目前果酒催陳方法有電化學催陳、微波催陳、超高壓催陳、氧化催陳等[7]。
1.2 主要風味化合物的合成及調控機理
高級醇類和酯類物質是發酵香氣的重要組成部分,幾乎存在于所有發酵果酒中。因此,研究其合成機理有利于從分子水平上精確調控化合物的含量,提升果酒香氣的強度和復雜度[8]。
1.2.1 高級醇類物質
高級醇類物質主要由酵母在α-酮酸發酵過程中形成,與Ehrlich途徑降解氨基酸有關[8]。高含量的高級醇會產生威士忌等異味,而低含量的高級醇則會賦予酒青草香、果香等香氣[9]。因此,可以通過調節Ehrlich途徑調控高級醇的生成。酵母生長階段、相關酶活力和氨基酸含量等因素均會影響Ehrlich途徑的代謝過程,從而影響果酒香氣成分組成。如與Ehrlich途徑相關的基因BAT1在酵母指數生長期優先表達,而BAT2在穩定生長期過表達,這兩個基因的過表達會使異丁醇和異戊醇含量顯著增加[10]。
1.2.2 酯類物質
酯類物質是在果酒中發現的最豐富的香氣化合物,包括乙酸酯和乙酯兩類[11],它們由酵母通過不同的代謝機制形成,但均具有類似水果的芳香,如草莓、甜果、青蘋果、香蕉等氣味[12]。乙酸酯的形成依賴于乙酸和高級醇,也受釀酒酵母中ATF基因編碼的兩種醇乙酰轉移酶的影響[8]。例如,葡萄酒發酵過程中過表達ATF1基因會導致乙酸酯的產量增加約10~200 倍[13],但酵母過表達ATF1生產乙酸乙酯會對酒中的其他酯類以及揮發性物質產生抑制作用,從而掩蓋葡萄酒的品種香氣,并產生清漆或指甲油等不良氣味[14]。
乙酯的生物合成受乙醇含量的影響較大[15],較低含量的乙醇有利于乙酯的生成,因此研究人員特別重視降低葡萄酒中乙醇的水平[16-18]。最近一項研究發現,通過推動碳通量生產甘油能夠減少乙醇的生成[19],然而,這種方法會造成醋酸的積累,使揮發性酸度處于很高水平,導致釀酒故障。另一種能夠有效降低乙醇產量的方法是從分子水平去除乙醛脫氫酶基因(ALD6)[20]。與酵母代謝相關的主要化合物的合成如圖2所示。

圖2 與酵母代謝相關的主要化合物的合成[8]Fig.2 Synthesis of major compounds related to yeast metabolism[8]
2 果酒增香發酵
2.1 釀酒微生物對香氣的影響
2.1.1 酵母對香氣的影響
產香酵母又稱產酯酵母,是一類能夠改善發酵產品風味的酵母。自20世紀60年代開始用于白酒生產,之后陸續應用于黃酒、醬油、食醋、葡萄酒、果酒等行業,用于彌補香氣不足的缺點。產香酵母能分泌豐富的糖苷酶類,在代謝過程中產生大量酯類、醇類等香氣物質,使酒體協調,賦予酒體濃郁的發酵香味。果酒中常見的產香酵母包括克魯維畢赤酵母(Pichia kluyveri)、葡萄汁有孢漢遜酵母(Hanseniaspora uvarum)、鐵紅假絲酵母(Candida pulcherrima)、東方伊薩酵母(Issatchenkia orientalis)、季也蒙有孢漢遜酵母(Hanseniaspora guilliermondii)等[21]。
2.1.1.1 克魯維畢赤酵母
克魯維畢赤酵母具有很強的產酯能力,特別是乙酸苯乙酯和辛酸乙酯[22-23],能夠賦予酒類特色的果味、玫瑰味、甜味和蜂蜜味。如由克魯維畢赤酵母發酵的蘋果酒具有濃郁的熱帶水果味[24]。同時,有研究證明克魯維畢赤酵母能夠增強β-裂解酶活性,從而促進揮發性硫醇的釋放,克魯維畢赤酵母與釀酒酵母混合發酵產生的硫醇含量為釀酒酵母單發酵的7 倍[25]。但在含有克魯維畢赤酵母的發酵液中發現了戊酸(腐臭氣味)含量的增加和硫化氫(臭雞蛋味)的產生,對酒體產生不利影響[26]。然而,它們的來源尚不確定,有待進一步研究。
2.1.1.2 葡萄汁有孢漢遜酵母
葡萄汁有孢漢遜酵母的主要特征之一是高產乙酸酯[27]。Ciani等[28]檢測到由葡萄汁有孢漢遜酵母發酵的白葡萄酒中乙酸乙酯含量約為釀酒酵母發酵的9 倍。Romano等[29]比較葡萄汁有孢漢遜酵母和釀酒酵母生產香氣化合物的能力,發現葡萄汁有孢漢遜酵母發酵的酒中乙酸平均質量濃度約為2 g/L,遠高于釀酒酵母發酵酒(0.6 g/L)。但是與釀酒酵母相比,用葡萄汁有孢漢遜酵母發酵葡萄酒產生的總脂肪酸含量和總高級醇含量要少得多,這有利于減少酒精味、指甲油味、脂肪味等不良氣味[30]。
2.1.1.3 其他產香酵母
鐵紅假絲酵母與酒中高含量酯的產生有關。在純培養條件下,鐵紅假絲酵母產生的乙酸異戊酯含量顯著高于其他非釀酒酵母,當質量濃度超過0.001 g/L時,呈現糖果味、水果味和香蕉味[15]。而與釀酒酵母進行混菌發酵時會產生較高水平的乙酸乙酯以及一些與不良風味有關的揮發性物質,當乙酸乙酯質量濃度為0.2 g/L時,葡萄酒的果味與乙酸乙酯呈強相關[31]。季也蒙有孢漢遜酵母與酒中玫瑰和蜂蜜味有關[32],而Hanseniaspora vineae是果香椰子和香草味形成的主要原因[33],此外,Hanseniaspora vineae中支鏈氨基酸轉氨酶(BAT2)和醇酰基轉移酶(EEB1)基因的缺失會導致高級醇、脂肪酸和乙酯產量的減少[8]。東方伊薩酵母的存在會導致酒中乙酸乙酯(220~730 mg/L)的過量生成,但其較強的蘋果酸降解能力對于發酵是有利的[34]。
2.1.2 混菌發酵對香氣的影響
目前果酒工業生產采用釀酒酵母單菌發酵,其發酵速率快、效率高,但生產的果酒缺乏風味復雜性。非釀酒酵母雖然發酵性能差,但在發酵過程中可以通過酶和風味代謝產物的產生改善發酵酒的芳香特性。因此,利用釀酒酵母與非釀酒酵母混合發酵提升果酒感官質量是一種可行的辦法。
2.1.2.1 混菌發酵菌種的選擇
相較于傳統的發酵方式,利用非釀酒酵母與釀酒酵母混合發酵對改善果酒的風味和口感具有積極作用。尹雪林等[35]利用釀酒酵母與戴爾凱氏有孢圓酵母混合發酵獼猴桃酒,共檢出23 種揮發性物質,遠多于純種發酵,豐富了獼猴桃酒的香氣成分,其中β-大馬士酮、香茅醇、α-松油醇等萜烯類物質大量生成,使得獼猴桃酒的花果香更加濃郁。張文文等[36]對釀酒酵母和東方伊薩酵母混合發酵楊梅酒產生的揮發性風味成分進行比較,發現乙酯類和乙酸酯類物質含量較釀酒酵母單發酵分別提高了18.52%和57.34%,同時提高了發酵效率,降低了釀酒酵母單發酵產生的酵母味和癸酸乙酯帶來的澀味。采用不同種類的非釀酒酵母與釀酒酵母混合發酵黃桃果酒,產生的香氣化合物具有較大的差異,Lachancea thermotolerans組和Metschnikowia pulcherrima組分別有利于苯乙醇和甘油的生成。H.uvarum/S.cerevisiae組的乙酸乙酯含量最高,而T.delbrueckii組高級醇和脂肪酸含量降低,從而減弱了酒中刺鼻的氣味[37]。不同果酒混菌發酵的香氣特點如表1所示。

表1 不同果酒混菌發酵的香氣特點Table 1 Aroma characteristics of different fruit wines produced by mixed fermentation
2.1.2.2 接種方式的選擇
在混合發酵過程中,接種策略也會影響酒體香氣特征。研究發現順序接種發酵比同時接種發酵效果更好,因為順序接種時非釀酒酵母在發酵開始階段能進行更高程度的代謝表達[48-49]。在櫻桃酒發酵中,順序接種比同時接種發酵產生更多的酯類和高級醇類,包括3-甲基-1-丁醇、己酸乙酯、辛酸乙酯、β-苯乙醇和芳樟醇等,這可能是由于同時接種導致與釀酒酵母的氮競爭,抑制了戴爾凱氏有孢圓酵母的生長[50]。然而,在龍眼酒中顯示了與之矛盾的結果,同時接種發酵產生了最高含量的總酯和苯乙醇[45]。
接種比例也是一個必須要考慮的問題。據報道,以1∶1同時接種釀酒酵母和拜氏接合酵母有利于生產乙酯,包括乙酸乙酯、己酸乙酯、辛酸乙酯和癸酸乙酯等。然而,當接種比例為1∶100或1∶1 000時會生成過量的乙酸乙酯,產生不良氣味[51]。張文靜等[39]采用本土克魯維畢赤酵母與釀酒酵母分別以1∶1、10∶1、20∶1的比例混合接種發酵赤霞珠葡萄酒,發現克魯維畢赤酵母接種比例的增加有利于葡萄酒中丁酸乙酯、己酸乙酯、辛酸乙酯、異戊醇和乳酸乙酯等香氣物質含量的提高。Canonico等[52]比較了在曝氣條件下,3 種非釀酒酵母與釀酒酵母順序接種發酵對葡萄酒揮發性成分的影響,發現只有拜氏接合酵母能稍許增加酯類的產量。與釀酒酵母單發酵相比,接種拜氏接合酵母的葡萄酒乙醇體積分數降低了約2%[53];同時花青素含量增加了36%,改善了葡萄酒的顏色,這可能是由于酵母細胞壁中花青素吸附能力的差異[54]。
此外,隨著現代生物技術的迅猛發展,基因組學、蛋白質組學以及代謝組學等技術逐漸運用到提升果酒風味方面。Giorello等[8]利用全基因組測序技術分析發現2-苯乙酸酯和2-苯乙醇含量的增加可能與芳香氨基酸轉氨酶(ARO8和ARO9)和苯丙酮酸脫羧酶基因的復制有關。Kondo等[55]通過在釀酒酵母中過表達2-酮酸脫羧酶基因(KDC)和醇脫氫酶基因(ADH6、ADH7),檢測到更高含量(13 倍)的異丁醇。另外,通過改變纈氨酸生物合成的碳通量和敲除丙酮酸脫羧酶基因(PDC1),可進一步提高異丁醇的產量。ATF1和ATF2基因雙缺失會導致乙酸異戊酯無法合成[56],而IRC7基因是影響葡萄酒中4-巰基-4-甲基-2-戊酮形成的關鍵基因[57]。
2.1.3 蘋果酸-乳酸發酵對香氣的影響
蘋果酸-乳酸發酵是乳酸菌將蘋果酸轉化為乳酸和CO2的過程。該過程除了能夠降低酒液酸度外,還可以調節酒的香氣和口感。酒酒球菌、乳桿菌和片球菌等可以通過糖苷酶水解香氣前體物質影響酒體香氣特征。Spano等[58]報道了β-葡萄糖苷酶基因在乳桿菌中的表達情況,其受到乙醇含量、pH值以及菌株等因素的影響。Olguín等[59]推斷,在中等乙醇體積分數(8%~10%)條件下可能會誘導酒酒球菌中bgl基因的表達。然而,β-葡萄糖苷酶活性與基因表達之間的關系尚不清楚,還需要更多的研究。此外,菌種間糖苷酶活性的差異會影響香氣化合物的含量,Michlmayr等[60]報道,酒酒球菌組糖苷酶處理葡萄酒中產生的芳樟醇和香茅醇含量明顯高于乳桿菌組。此外,乳酸菌與酵母菌混合發酵對酯類的含量有顯著影響,最近一項研究顯示,混合接種發酵使乙醇發酵與蘋果酸-乳酸發酵同時進行,縮短發酵時間的同時提升了果酒香氣,乙酯含量顯著增加,尤其是乳酸乙酯和琥珀酸二乙酯[61]。
綜上,發酵香氣的形成不再單一地依賴釀酒酵母,利用釀酒酵母、非釀酒酵母以及乳酸菌等多菌種間的相互作用調控果酒風味品質將受到更多關注,今后對多菌種間的分子相互作用機理應作深入研究。
2.2 風味相關生物酶對香氣的影響
果酒釀造過程可將水果果實中以糖苷鍵合態形式存在的風味物質解離釋放到酒中。釀酒微生物能夠代謝產生一些風味酶,包括糖苷酶、果膠酶、蛋白酶等,這些酶能夠水解香氣前體物質,從而促進香氣化合物的釋放。在釀造前加入外源糖苷酶、果膠酶和糖基水解酶可提高果酒的風味品質[62]。
2.2.1 糖苷酶
糖苷酶可以促進水果中糖苷香氣前體物質的釋放,主要包括萜烯類和一些多酚類化合物,從而增強和豐富果酒的風味。糖苷鍵合態前體物質本身不具有香氣活性,但可以在β-糖苷酶等香氣形成關鍵酶作用下轉化為具有香味的香氣物質。萜類糖苷鍵合態前體物一般由單葡萄糖或雙葡萄糖和萜類糖苷配基組成,其中葡萄糖可以進一步與其他糖配基結合,例如α-L-阿拉伯呋喃糖基、α-L-鼠李糖基、β-D-吡喃木糖基,以及β-呋喃芹糖基等[63]。在葡萄冷浸漬中,萜類物質和C13-降異戊二烯類物質通過生物代謝途徑,如糖苷鍵合態前體的水解或生成,這一過程可能受到葡萄果實中β-糖苷酶的活性變化影響。
一些非釀酒酵母包括假絲酵母屬(Candida)、畢赤酵母屬(Pichia)、漢遜酵母屬(Hanseniaspora)等均表現出良好的β-葡萄糖苷酶活性[64]。例如,葡萄酒中萜烯類以及降異戊二烯類物質含量的增加與葡萄汁有孢漢遜酵母中β-葡萄糖苷酶的活性密切相關[65]。Liu Xiaozhu等[66]從刺梨中篩選出Wickerhamomyces anomalusC4菌株的β-葡萄糖苷酶活性最強,為釀酒酵母X16的1.65 倍,并且體積分數15%的乙醇、5 g/100 mL葡萄糖、Cu2+等因素均會抑制其活性。來源于不同菌株的β-葡萄糖苷酶活性及產香能力也有明顯差異。Hu Kai等[67]應用半定量比色法篩選了來自中國3 個不同地區的酵母菌——H.uvarum、P.membranifaciens、Rhodotorula mucilaginosa,發現來自H.uvarum中的β-葡萄糖苷酶活性最高,且對C13-正異戊二烯和一些萜烯的芳香糖苷物質具有催化特異性,豐富了葡萄酒中新鮮花香、漿果和堅果等香氣。
2.2.2 果膠酶與蛋白酶
果膠酶可破壞植物細胞壁結構,將果膠聚合物降解為半乳糖醛酸,從而促進果汁澄清過濾。同時,在果酒發酵前的浸漬過程中,果膠酶可以將糖苷鍵合的風味前體切割成芳香活性苷元,從而增強果酒的風味特性。例如,Rollero等[68]從設拉子葡萄汁中篩選出一株具有高果膠酶活性的酵母Kluyveromyces marxianus,此菌株可促進葡萄酒發酵過程中產生更多的苯乙醇和苯乙酸乙酯,而且還能夠增加多酚和花青素的含量。
蛋白酶主要通過為酵母提供氮源影響果酒香氣的形成。非釀酒酵母是蛋白酶的重要來源,在最近的一項研究中,將來自美極梅奇酵母異源表達的天冬氨酸蛋白酶MpAPr1添加到長相思葡萄汁中[69]。結果表明,該酶在發酵過程中具有活性,并在一定程度上降解了葡萄酒蛋白[70],促進氨基酸水解,為釀酒酵母的生長提供更好的氮源,從而對香氣化合物產生有利影響[71]。此外,E3泛素-蛋白連接酶被證明可通過增加Ehrlich通路相關基因ARO9和BAT1以及去甲基化通路基因STR3表達,抑制ARO10、PDC1、PDC5和PDC6基因表達,調節高級醇的生成[72]。這兩種途徑的控制為工業應用提供了巨大的潛力,包括乙醇生產過剩和風味質量控制。然而,目前為止仍未找到合適的控制Ehrlich途徑和去甲基化途徑的方法。
2.2.3 其他酶
關鍵的單萜和降異戊二烯香氣化合物在釀酒過程中被酵母和產香細菌分泌的糖苷酶、酯酶、C—S裂合酶和脫羧酶等降解為游離態釋放[62]。目前,提高單帖類化合物策略主要有以下兩種:一種是在發酵過程中或發酵后添加來自其他微生物的外源性酶制劑以改善糖基化結合物的水解,商業酶制劑一般包含果膠酶、葡聚糖酶和木聚糖酶,主要從曲霉中獲得,然而這些酶缺乏特異性,可能會導致對風味不利的二次反應;另一種是采用少數非釀酒酵母,如葡萄汁有孢漢遜酵母的水解酶活性較高,能夠促進與糖苷結合的單帖類物質前體水解,導致單萜類物質含量增加[73]。
從以上研究結果可以發現,發酵過程中添加風味胞外酶和某些非釀酒酵母可以有效改善成酒的品種香氣。其主要原理是利用外源酶或非釀酒酵母分泌的高活性糖苷酶水解水果果皮中的香氣糖苷前體,從而釋放游離態品種香氣成分,增強果酒的典型風味。
2.3 發酵前處理工藝對香氣的影響
在葡萄破碎期間,酶促氧化反應、β-胡蘿卜素物質(其他類胡蘿卜素)的降解被認為能夠產生多種類型的降異戊二烯類物。這類化合物主要包括β-大馬士酮(奇異水果的香氣)、β-紫羅蘭酮(紫羅蘭花的香氣)、3-氧代-β-紫羅蘭酮(煙草氣味)和Vitispirane(新鮮的花香果香和/或異國花卉和泥質木本植物的香氣)等。低溫冷浸漬工藝可以保留小分子香氣化合物,增加香氣復雜性。有研究表明,在冷浸漬和低溫發酵過程中,醇類、酯類、烷烴類物質等發酵香氣也會有所增加,脂肪味和化學味有所下降[74]。在釀酒酵母與畢赤酵母混合發酵過程中,適度延長冷浸漬時間(3 d),3-甲基丁醇、乙酸乙酯和乙酸異戊酯的含量會明顯增加。此外,如芳樟醇、α-萜品醇和β-香茅醇等萜烯類香氣成分主要來源于果皮,說明通過冷浸漬工藝可以充分浸提果皮中的芳香類物質,提高果酒的品種香氣,從而增加果酒的香氣濃郁度。
由于天然酵母、微生物和相關酶的作用,較高含量的己醇、順-6-壬烯醇主要是在浸漬過程中形成[75]。冷浸漬過程中一些耐低溫的產香酵母可以使葡萄酒的主香氣成分含量增加[76]。研究發現,通常當溫度超過25 ℃時就會抑制非釀酒酵母的生長。有時低溫更有利于其發酵作用,如檸檬型克勒克酵母在10 ℃較25 ℃時發酵更好,該酵母可產大量的甘油,可以調節細胞內的氧化還原電位,使葡萄酒具有更好的柔和性,這種甘油含量高的葡萄酒在新西蘭、澳大利亞都很受歡迎。
3 產香酵母的選育
酵母產香能力直接影響成酒的風味質量,因此酵母的選育備受重視。酵母的選育方法主要包括自然突變菌株的選育、雜交育種、基因工程育種、誘變育種等[77]。傳統的育種方法以自然突變為主,從不同原料(水果、土壤)中篩選并分離出具有特殊性能的新菌株。例如,吳卓凡等[78]從自然界中篩選出發酵能力強且風味優良的枇杷果酒專用釀酒酵母,測定并分析了其風味物質組成,相比商業菌株,醇類、酯類和酸類物質含量分別高31%、86%和25%,總揮發性物質增加了45%,具有良好的工業應用前景。馮濤等[79]從不同地區的葡萄園土壤中篩選分離出產香性能良好的釀酒酵母,測定并比較了其與商業酵母在產香方面的差異,證實了篩選出的酵母可以顯著提升發酵酒的整體香氣。
雜交育種可以將兩個或多個優良性狀結合,構建具有優良性狀的新菌株。Wang Zhouli等[80]通過原生質體融合技術構建了基于釀酒酵母和乙醇念珠菌的新型重組融合酵母,在釀造香氣濃郁且低酒精度的蘋果酒方面有巨大潛力。基因工程育種由于外源基因的安全性一直未得到認可,其在酵母選育中應用受到很大限制。
誘變育種方法可以使生物突變率顯著升高,但要獲得性狀優良的正向突變菌株需要花費相當長的時間,并且優良酵母選育涉及到較多的釀酒工藝學指標(如產乙醇能力、發酵速率、高糖耐受性、pH耐受性等),需要進行多年、多點的生產試驗,篩選進程相對較慢。近幾年常壓室溫等離子體誘變技術的出現有效地解決了這一難題,并廣泛用于篩選優良產香酵母。Tian Tiantian等[81]通過一種聯合策略,包括常壓室溫等離子體誘變系統、高通量篩選技術以及適應性進化策略,從突變庫中獲得了一株耐酸性強、產香和遺傳穩定性良好的菌株ET008-c54,將該菌株用于發酵青梅酒,使關鍵風味化合物的總質量濃度由17.8 mg/L提升至27.0 mg/L,提高了51.9%,特征香氣物質安息香醛質量濃度由1.8 mg/L提升到2.8 mg/L,解決了青梅酒風味不足和典型性不突出的問題。馮鵬鵬等[82]同樣采用常壓室溫等離子體誘變方法獲得了低產高級醇的誘變菌株,提高了成酒的風味品質。
4 結語
本文綜述了釀酒微生物、酶對果酒香氣的影響,盡管利用產香酵母提高葡萄酒風味的復雜性越來越普遍,但非釀酒酵母在其他果酒生產中的應用仍然很少。且果酒中部分關鍵風味化合物的合成途徑及微生物代謝通路仍需要進一步研究。當前,果酒釀造過程中釀酒酵母與非釀酒酵母分子相互作用機制仍不清晰,高通量技術可用于揭示微生物的相互作用,從而有利于更好地設計混合培養物,提升混菌發酵的控制水平。此外,果酒陳釀周期較長,但目前催陳技術仍不成熟,有待進一步研究。未來,在全基因組測序技術快速發展的帶動下,基因組學、蛋白質組學、代謝組學以及風味組學的相關技術也將促進對微生物合成風味化合物代謝途徑的認識和對風味代謝物質調控機理的探索,為創制果酒增香釀造技術奠定基礎。
