富集γ-氨基丁酸鷹嘴豆酸面團的工藝及其流變特性
2023-08-15 13:47:02郭小雨張濤
食品與發酵工業 2023年15期
郭小雨,張濤
(江南大學 食品科學與技術國家重點實驗室,江蘇 無錫,214122)
酸面團是由谷物、水和活性微生物,如乳酸菌和(或)酵母菌,經過發酵制得的面團,是一種歷史悠久的面食發酵劑。這些微生物將內在化學成分發酵成有機酸、風味化合物和其他重要代謝物,影響發酵面團或烘焙食品的質地、感官、營養和保質期特性[1]。目前,除小麥或黑麥外,其他谷物面粉,如鷹嘴豆,由于其獨特的營養和功能特征,已被提議用于制作烘焙產品的酸面團[2]。在小麥粉中加入豆類面粉可以制成富含蛋白質的面制品,鷹嘴豆的氨基酸分布均衡和纖維含量高[3],可提高面包營養特性,為消費者提供一系列健康益處。
γ-氨基丁酸(gamma-aminobutyric acid, GABA)是神經系統中重要的抑制性神經遞質[4],在預防糖尿病[5]、抗焦慮[6]、降血壓[7]以及促進睡眠[8]等方面具有十分重要的作用。同時GABA還是食品當中重要的生物活性成分添加劑。因此富含GABA的功能性新食品的開發越來越受研究者關注,可將其用于發酵健康食品,制作富含GABA的功能性食品,以滿足消費者需求。豆類含有豐富的底物谷氨酸,是乳酸菌產GABA重要的發酵基質。其中,鷹嘴豆蛋白質含量是小麥等谷物的2倍,它們是高質量、低成本且環境友好的蛋白質來源,可以用于控制和預防高血壓等疾病[9]。利用高產GABA乳酸菌發酵酸面團以增加面制品中GABA含量,將GABA以面包等成品的形式呈現,使其成為消費者日常補充GABA的良好途徑。
本研究以鷹嘴豆為發酵基質,通過胞外多糖(exopolysaccharides, EPS)、GABA等活性成分指標,選擇短乳桿菌作為發酵菌株。以小麥面包面團(wheat bread dough, WBD)、鷹嘴豆面包面團(chickpea bread dough, CBD)和化學酸化鷹嘴豆面包面團(lactic/acetic acid acidified bread dough, LA/AA-BD)為對照,研究富集GABA乳酸菌發酵鷹嘴豆添加對面包面團的流變特性等影響,為開發富集GABA功能性發酵食品的應用奠定理論基礎。
1 材料與方法
1.1 主要材料和試劑
金龍魚面包用高筋小麥粉(淀粉質量分數68.54%,蛋白質質量分數11.56%),益海嘉里糧油;高活性干酵母(高糖型),安琪酵母有限公司;優級白砂糖,中糧集團(中國)有限公司;食用鹽、新疆卡布里鷹嘴豆,市售;上述均為食品級。MRS固/液體培養基、酸面團,由實驗室自制。
1.2 主要儀器設備
ZCZY-CS8型恒溫培養搖床、DNP-9082型恒溫培養箱,上海知楚儀器有限公司;GI54DWS型立式高壓滅菌鍋,廈門致微儀器有限公司;高效液相色譜系統(VWD檢測器),美國安捷倫(Agilent)公司;TA.XT Plus物性分析儀,英國SMS公司;DISCOVERY HR-3流變儀,美國TA儀器公司。
1.3 實驗方法
1.3.1 具GABA富集能力的乳酸菌初篩及酸面團發酵
1.3.1.1 鷹嘴豆發酵基質的制備
將保藏的6株乳酸菌作為發酵菌株,分別為嗜熱乳桿菌ST(StreptococcusthermophilusHH-ST08)、保加利亞乳桿菌IB(LactobacillusbulgaricusHH-IB57)、植物乳桿菌LP(LactobacillusplantarumHH-LP56)、植物乳桿菌FS(LactobacillusplantarumFSB7)、植物乳桿菌CI(LactobacillusplantarumCICC21796)和短乳桿菌CG(LactobacillusbrevisCGMCC1.214)。取上述乳酸菌以體積分數1%接種量接種至液體MRS培養基中,30 ℃培養6 h。將菌液在4 ℃、6 000 r/min離心10 min,棄上清液,用生理鹽水洗滌菌體2次并重懸于無菌水中。水與生鷹嘴豆粉按質量比1∶1混合,使其總菌落數達到107CFU/g,于30 ℃發酵24 h。為提高乳酸菌鷹嘴豆酸面團中EPS產量,將10%(質量分數)的鷹嘴豆粉用蔗糖替代。發酵完成后得到上述6株乳酸菌對應的鷹嘴豆酸面團,分別簡稱為SCSD、ICSD、LCSD、FCSD、CCSD和BCSD;未發酵的鷹嘴豆面團為CD。
1.3.1.2 不同乳酸菌發酵鷹嘴豆中pH、可滴定酸度(titratable acid,TTA)和菌落總數的測定
按配方制備的發酵基質在30 ℃培養箱中觀察24 h,定時取樣。參考KATINA等[10]的方法,測定鷹嘴豆酸面團發酵前后pH和TTA。
用90 mL 8.5 g/L 的生理鹽水稀釋10 g鷹嘴豆酸面團,測定樣品中的乳酸菌菌落數,繼續稀釋并在MRS瓊脂上接種100 μL的稀釋液,30 ℃恒溫培養箱中培養48 h用于菌落計數。
1.3.1.3 鷹嘴豆酸面團凍干粉的制備
將樣品放入-80 ℃冰箱中冷凍2 h后,置于冷凍干燥機中凍干。完成后使用研缽研磨成粉狀,置于干燥器中以備后續實驗使用。
1.3.1.4 鷹嘴豆酸面團中理化性質的測定
參照程新[11]的方法利用HPLC測定有機酸含量,并計算發酵熵。發酵熵是酸面團產生的乳酸與乙酸的比值。
參考TANG等[12]的方法測定EPS的含量;參考羅昆等[13]的方法測定多肽分子質量的分布;參考張賓樂[14]的方法測定游離氨基酸和GABA含量。
1.3.2 鷹嘴豆酸面團對面包面團性質的影響
1.3.2.1 面包面團制作
面包面團配方詳見表1。將除黃油外的所有配料倒入攪拌缸中,設置攪拌參數:慢速攪拌3.0 min,使原料混合均勻;快速攪拌3.0 min,將原料攪拌成團。加入黃油后慢速攪拌1 min,再快速攪拌3.0 min,至面筋網絡結構形成。將攪拌好的面團取出,覆上保鮮膜,室溫下擠壓走氣泡,松弛10 min后收于自封袋中,備用。WBD、CBD、LA/AA-BD和BCSBD分別代表小麥面包面團、鷹嘴豆面包面團、乳酸/乙酸酸化面包面團和鷹嘴豆酸面包面團。其中,LA/AA-BD是在CBD配方的基礎上,將850 g/L乳酸與醋酸按4∶1的體積比額外添加,面團pH值約為4.20,30 ℃培養箱中密封孵育24 h。
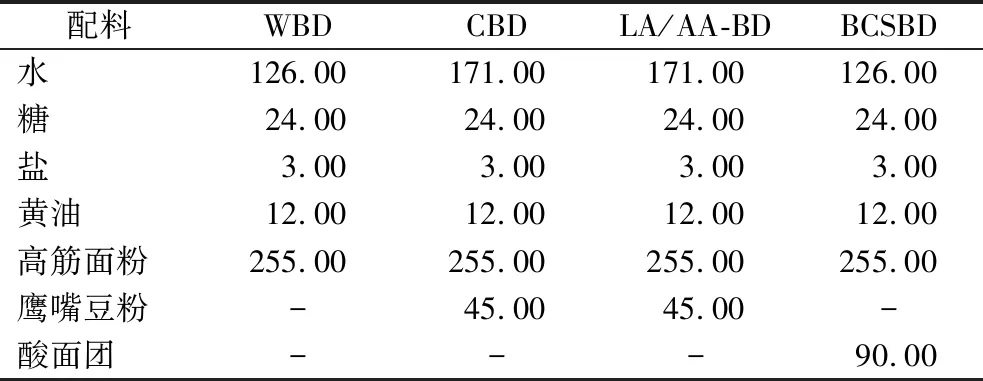
表1 四種面包面團的配方 單位:g
1.3.2.2 酸面團動態流變學特性分析
應用具有平板(直徑40 mm,間隙1 mm)的動態流變儀,在25 ℃下對面團進行振蕩頻率掃描測試。取2~3 g面團放置于帕爾貼板上,平衡松弛1 min,進行頻率掃描(0.1~10 Hz)測試,應變為0.20%。記錄彈性模量(G′)、黏性模量(G″)和損耗角正切(tanδ=G″/G′)。
1.3.3 數據處理分析
采用Origin Pro 2018以及SPSS 26等軟件對數據進行分析,數據重復3次,數值以平均值±標準差表示。
2 結果與分析
2.1 不同菌株發酵鷹嘴豆酸面團pH、TTA和菌落總數的測定
如圖1所示,酸面團的初始接種量為7 lg CFU/g。在鷹嘴豆發酵基質中,乳酸菌經過短暫的遲緩期迅速進入對數生長期。短乳桿菌CG(即BCSD中乳酸菌)在12 h趨于穩定,菌落總數達到10.30 lg CFU/g左右。隨著發酵時間的延長,乳酸菌繼續保持良好的生長繁殖的平衡態勢。短乳桿菌CG在整個過程中對數期最長、后期生長最穩定。
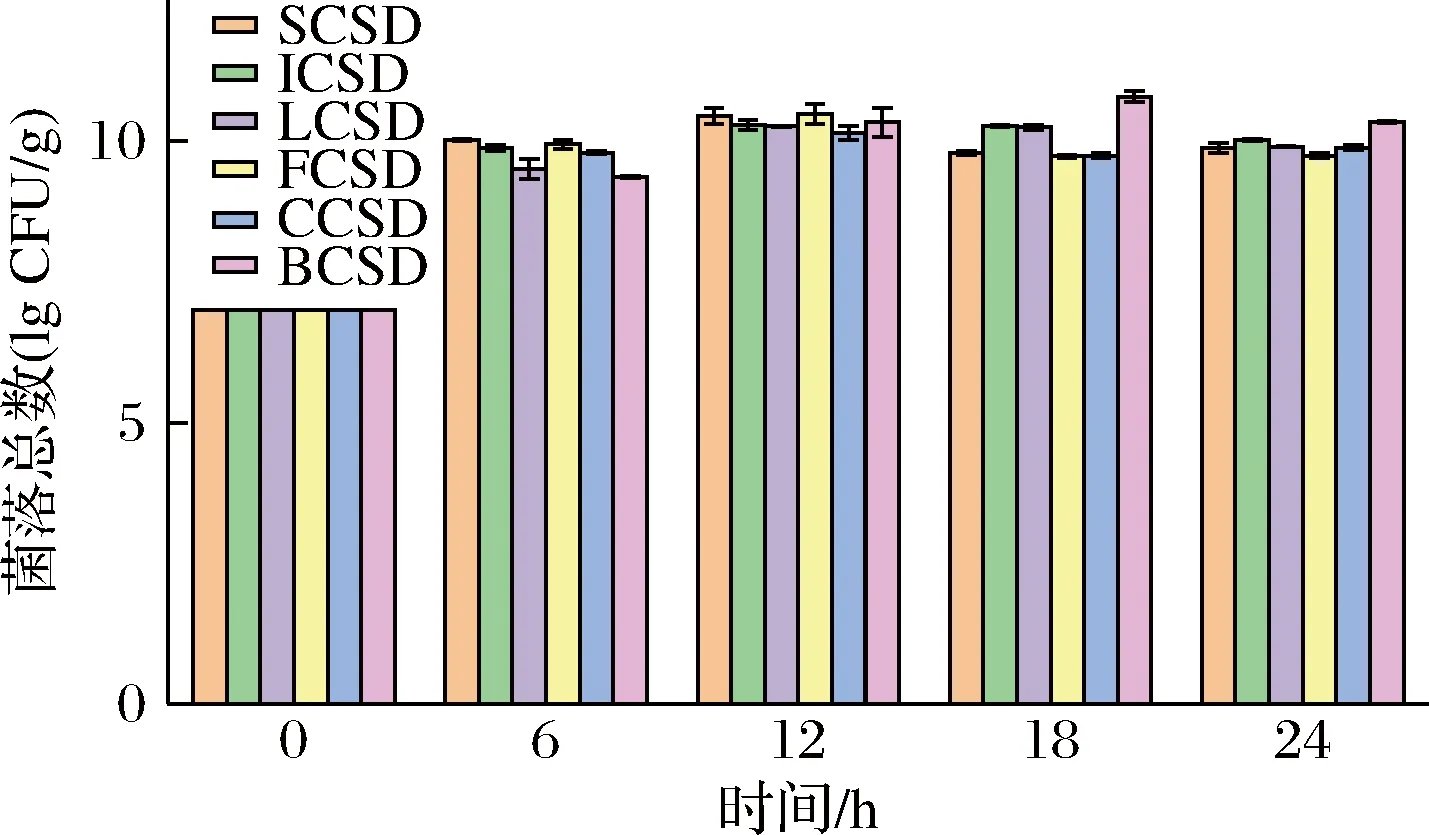
圖1 不同菌株發酵時期的菌落總數
發酵過程中,乳酸菌代謝產生多種有機酸。這一過程降低了酸面團的pH,提高了TTA值,并影響了發酵基質中的內源性酶活力,進而改變了酸面團的各種生化特性[15]。整個對數生長期正是乳酸菌快速累積產酸的時期,發酵后期pH變化趨于平緩,可能是因為代謝產物的積累通過反饋調節從而減緩后期pH下降速率。如表2所示,酸面團在30 ℃下發酵24 h時pH下降至4.00左右。與酸面團的pH變化趨勢相反的是,鷹嘴豆酸面團的可滴定酸含量隨著發酵時間的延長而逐漸上升,從最初的TTA為5.75~6.35 mL增加到發酵24 h時26.20~27.65 mL。6株乳酸菌中,BCSD的pH值最高,且TTA最大,表明反應到達滴定終點時所需的堿液含量最多,推測其發酵后可能形成了更多pKa大的有機酸,從而使酸性環境更加穩定、緩沖能力更強。
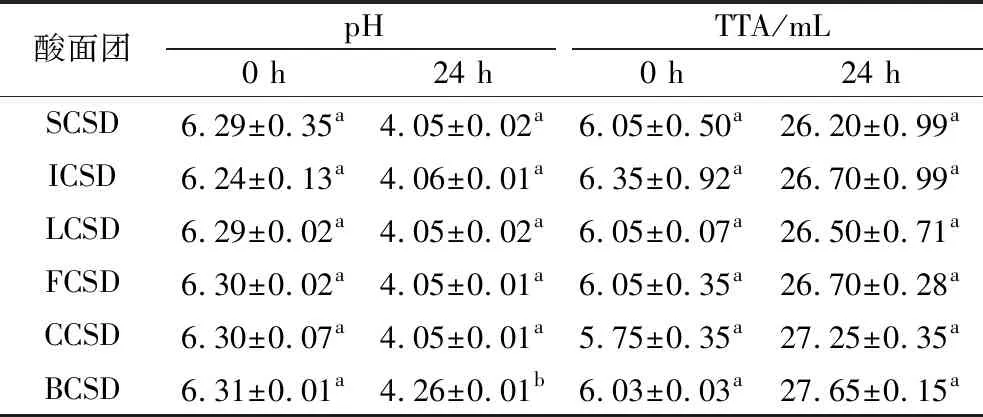
表2 不同菌株發酵時期的pH和TTA
2.2 酸面團有機酸分析
酸面團發酵時,微生物代謝發酵基質中的糖類物質并生成以乳酸和乙酸為主的有機酸[16]。發酵熵是影響酸面團發酵品質的重要參數,常用來關聯酸化和風味的發酵參數之一[17]。發酵24 h后,BCSD的乳酸和乙酸的含量最低,分別為28.10和7.38 mmol/100 g。有機酸的差異導致不同酸面團的TTA值不同,主要是因為不同有機酸躍變范圍不同[18],從而滴定時消耗的堿液量不同。從表3中可知,CCSD和BCSD最終發酵熵均在1.5~4的適宜范圍內。
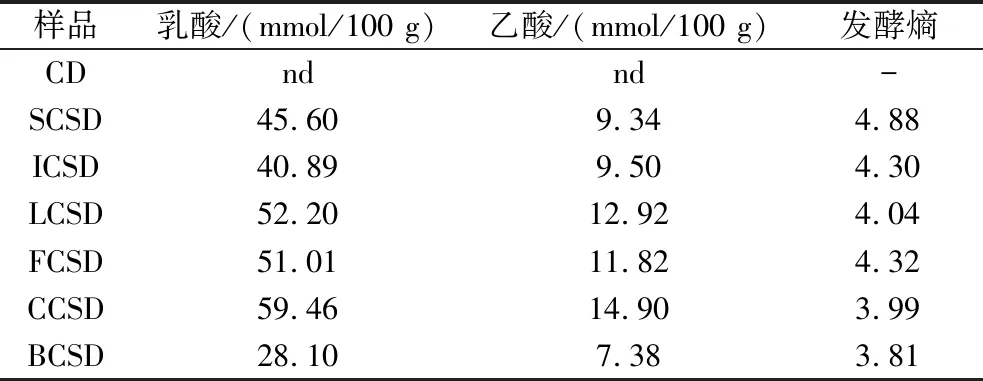
表3 不同菌株發酵前后有機酸含量和發酵熵
2.3 酸面團EPS含量測定
如圖2所示,BCSD中發酵產生的EPS量最多,為1 396.29 mg/L。結果表明,在乳酸菌發酵過程中,添加質量分數10%的蔗糖更加有利于菌種的生長代謝,加快其產酸速率。高糖和低pH等壓力條件使得乳酸菌在保護機制作用下更偏向于合成EPS來增強自身存活能力[19]。
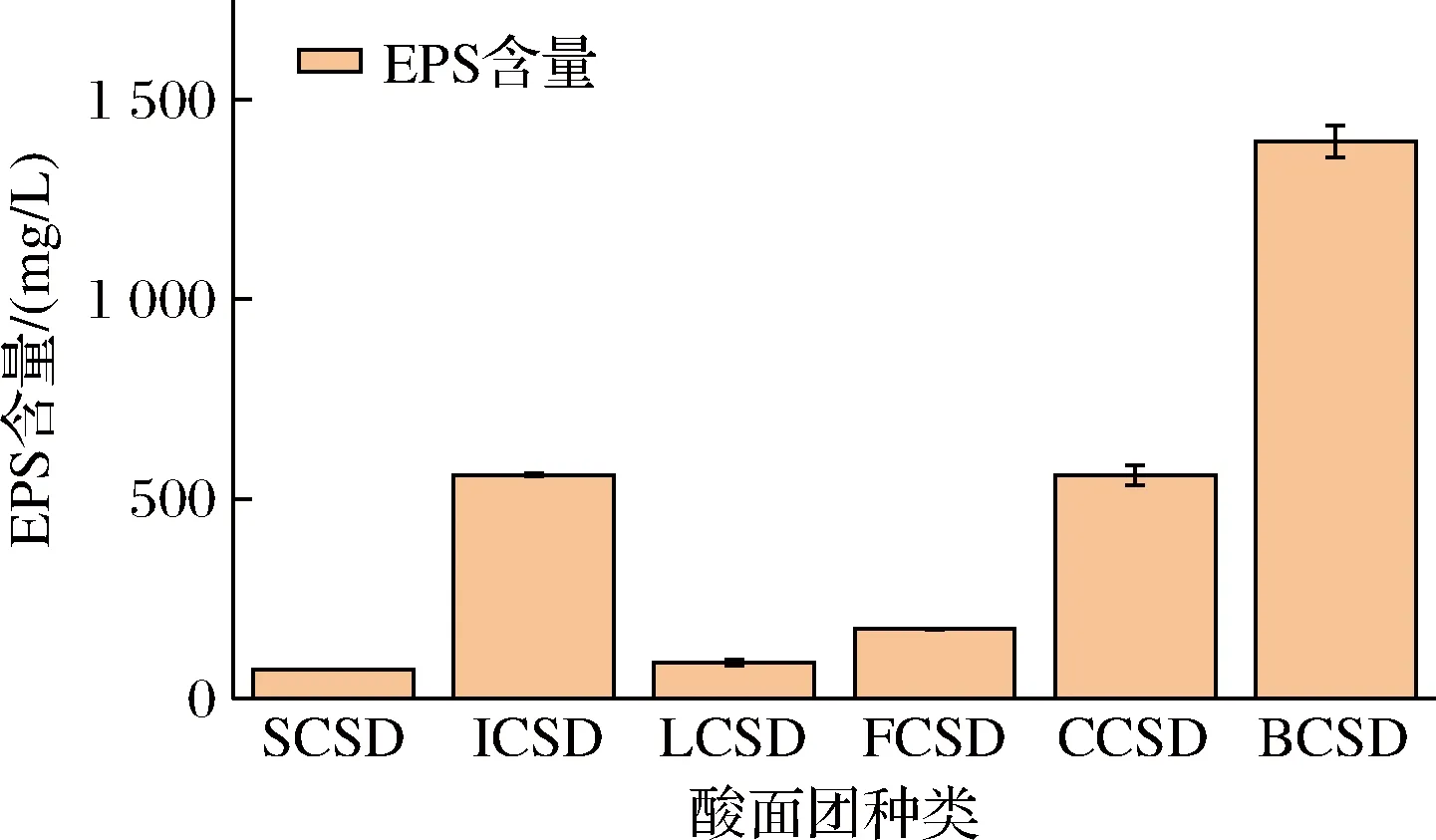
圖2 不同菌株發酵產生的EPS含量
2.4 酸面團游離氨基酸和GABA含量的分析
在乳酸菌發酵鷹嘴豆酸面團過程中,鷹嘴豆的內源蛋白酶和乳酸菌自身酶系會促進蛋白和糖類代謝產生氨基酸,其對面包的營養性、特征風味的形成以及整體感官有很大的積極影響[20]。圖3-a顯示,經短乳桿菌CG發酵得到的鷹嘴豆酸面團(BCSD)中總游離氨基酸含量為1 645.73 mg/100 g,與未經發酵的鷹嘴豆酸面團(755.18 mg/100 g)相比,游離氨基酸含量顯著提高。經乳酸菌發酵后,BCSD中必需氨基酸含量占總游離氨基酸的比例從鷹嘴豆面團(CD)的10.92%增至31.53%;由圖3-b可知,6組發酵乳酸菌組中,BCSD組的GABA含量最高,是CD組的49.43倍。GABA在鷹嘴豆酸面團發酵過程中得到富集,面團的營養價值增強。
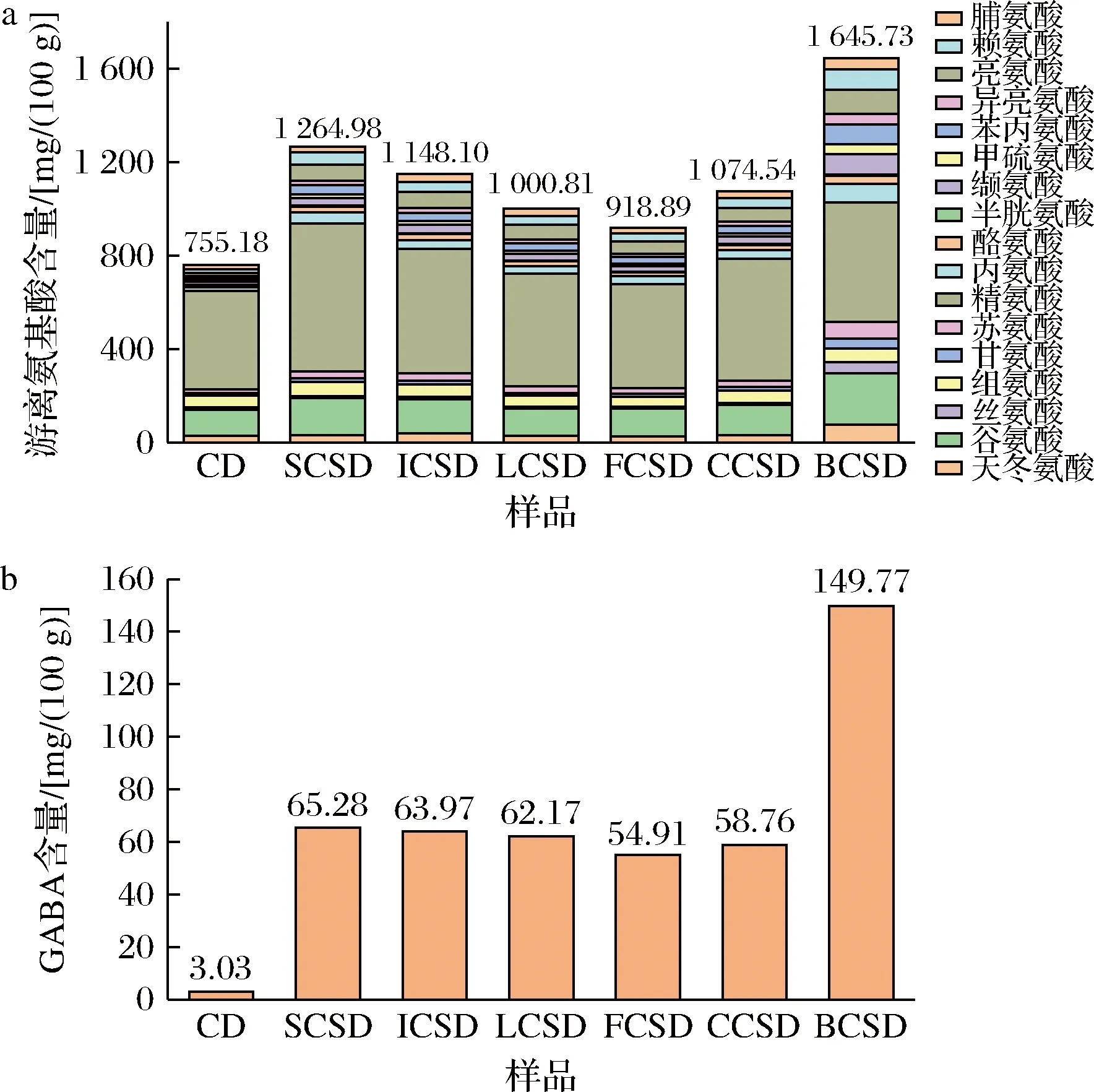
a-游離氨基酸含量;b-GABA含量
2.5 酸面團多肽分子質量分布的分析
由圖4所示,與對照組CD相比,經過6株乳酸菌發酵的鷹嘴豆酸面團的蛋白質得到很大程度的水解。經過24 h發酵,分子質量>5 000 Da的部分占比顯著降低,從65.95%降到5.89%~3.9%,而分子質量在3 000~5 000 Da的大肽含量提高了1.25~2.08倍。尤其是分子質量<1 000 Da的低聚肽含量增加的最為顯著,分別提高了47.37%~49.48%,BCSD中的低聚肽含量最多。小分子低聚肽直接被人體吸收,進而更好地發揮其生物學功能[21],增強產品的營養價值。實驗表明乳酸菌發酵能有效地水解蛋白質,提高鷹嘴豆酸面團的低聚肽含量。6株乳酸菌中短乳桿菌CG水解效果最好,可能是因為其自身產生的蛋白酶和肽酶更活躍[22],酸面團酸化的環境更有利于激活蛋白酶水解,從而提高了低聚肽含量。小分子低聚肽能參與酶的合成、激發酶的活性,從而形成蛋白水解的良性循環促進。該結果與鷹嘴豆酸面團中游離氨基酸的含量較一致。
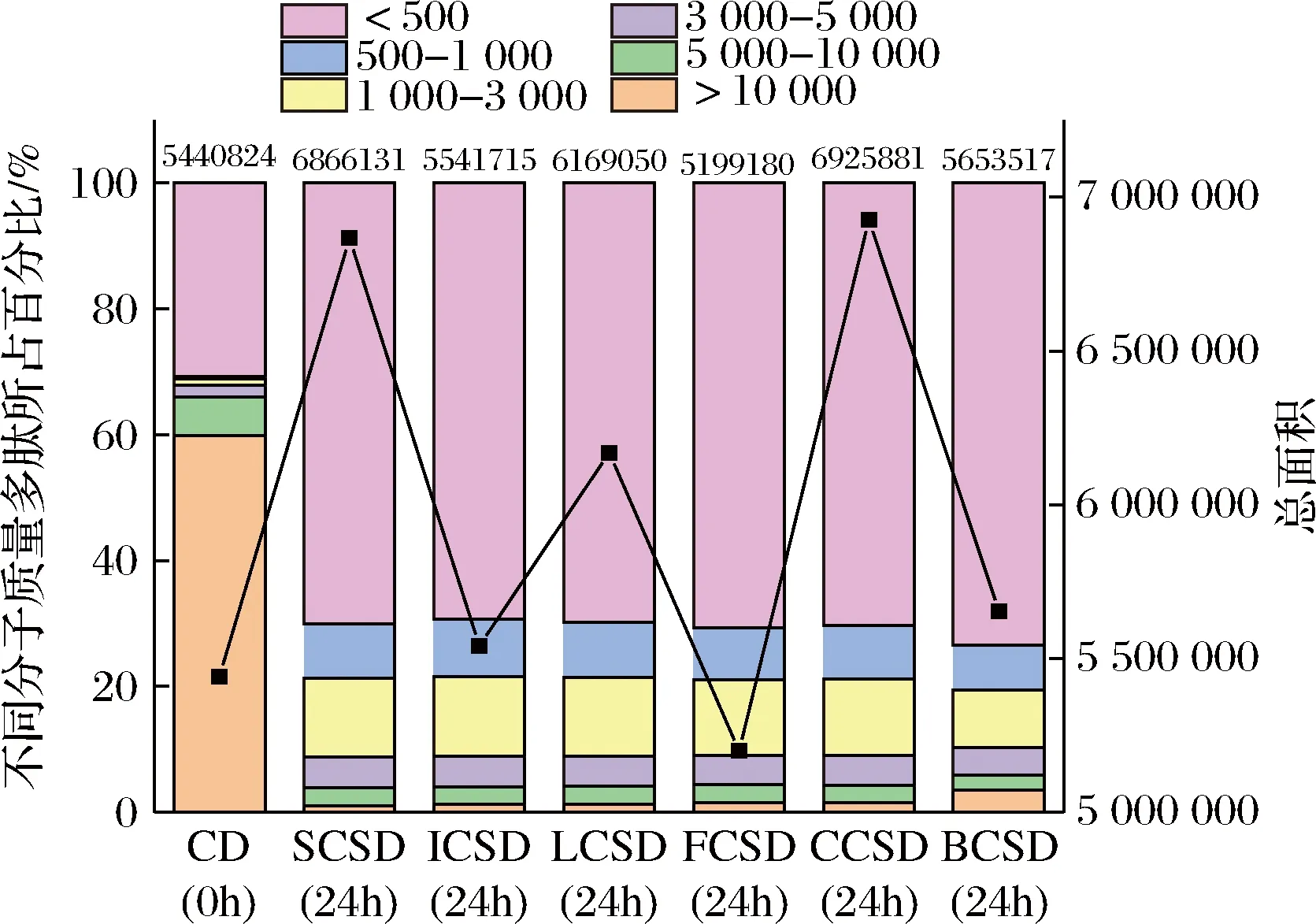
圖4 不同菌株發酵前后的多肽分子質量分布圖
2.6 流變學特性分析
如圖5所示,在同一掃描頻率下,各面團的G′、G″大小為WBD>BCSBD>LA/AA-BD>CBD。與WBD比,BCSBD的G′和G″均明顯下降。OSBORNE[23]和TAKEDA等[24]認為面筋蛋白在酸性pH下溶解度增加,可能是酸性環境中存在相當多的正凈
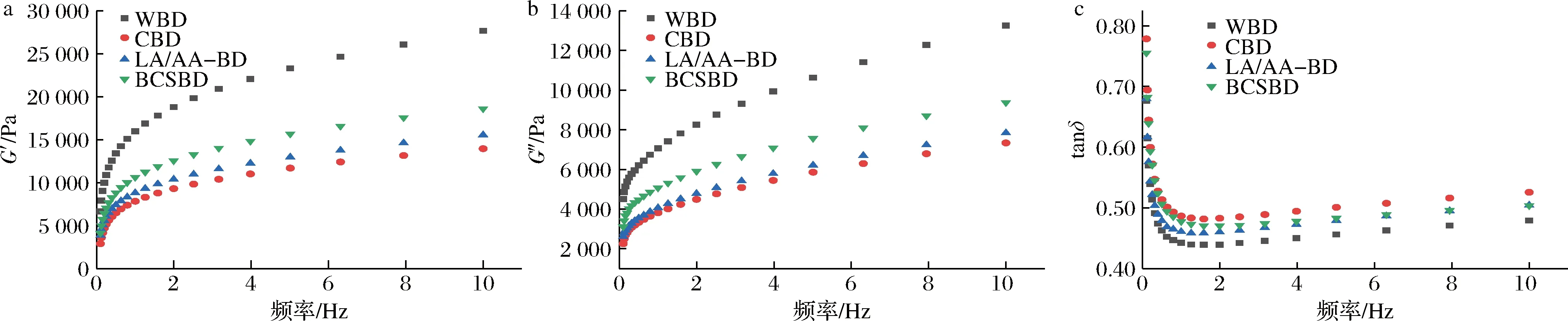
a-彈性模量;b-黏性模量;c-損耗角正切值
電荷[25],分子間靜電排斥增大導致面筋蛋白的展開和疏水基團的暴露增加,促進部分化學鍵的斷裂和阻止新鍵的形成,從而導致酸面團中的面筋網絡更加柔軟[26]。與LA/AA-BD相比,BCSBD的G′和G″都顯著提高。這可能因為乳酸菌產生的EPS可以作為親水膠體[27],改善面團的網絡結構并促進其與谷蛋白交聯,增強面團的黏彈性。與CAD相比,BCSBD的G′和G″都顯著提高。這可能是因為鷹嘴豆粉的添加稀釋了面團面筋,鷹嘴豆中的大分子蛋白質和淀粉顆粒等影響蛋白質分子之間二硫鍵的生成。損耗角(δ)表示面筋蛋白的弱化程度,tanδ值越大,表明弱化程度越顯著。從圖5-c可看出,隨著頻率的增加,4組面包面團的tanδ值均呈現先降低、后增大的趨勢。BCSBD的tanδ值較接近于WBD,說明其有類似的硬度,易于加工成型。而CBD的tanδ值最大,面團的流動性較強,不易成型。
3 結論
本研究以新疆鷹嘴豆作為發酵基質,篩選短乳桿菌CG發酵成富集EPS和GABA的酸面團。鷹嘴豆酸面團發酵工藝參數為:菌種接種時期為6 h,發酵溫度為30 ℃,發酵時間為24 h。此條件下,短乳桿菌CG在鷹嘴豆酸面團中生長良好,酸化能力強且GABA產量高。發酵后,鷹嘴豆蛋白降解效果最為明顯,能促進大分子質量蛋白降解為小分子蛋白和低聚肽,且游離氨基酸和必需氨基酸含量顯著增加。鷹嘴豆直接添加和化學酸化均不利于面團的形成及穩定,并且破壞面團的黏性和彈性之間的平衡。而乳酸菌發酵能夠降低其產生的不利影響,賦予面團良好的流變特性,使其更加穩定。為開發富集GABA功能性發酵食品的應用奠定理論基礎,此外,提高了鷹嘴豆在生產中的應用潛力。