發酵乳中乳酸菌菌株間互作機制及其對產品特性影響的研究進展
2023-08-25 07:16:28謝佳琪
食品工業科技 2023年17期
謝佳琪,趙 潔
(內蒙古農業大學,乳品生物技術與工程教育部重點實驗室,農業部奶制品加工重點實驗室,內蒙古乳品生物技術與工程重點實驗室,內蒙古呼和浩特 010018)
發酵乳是以生牛(羊)乳或乳粉為原料,經殺菌、發酵后制成的pH 降低的產品[1]。由于發酵乳具有更高的消化率,更高的營養價值,兼具抗高血壓、降低膽固醇、抗氧化或免疫調節等健康功效,使消費者對發酵乳的興趣與日俱增[2]。在過去的幾十年里,人們對酸奶、奶酪、開菲爾等發酵乳制品進行了大量的研究,發現乳制品在發酵過程中發生的主要生化反應是乳酸菌作用于乳糖,將乳糖轉化為乳酸,引起發酵乳的質構改變,以獲得理想的質地和粘度。同時,研究發現乳酸菌可以產生代謝物和生物活性化合物(如胞外多糖、細菌素等),賦予了發酵乳一定的功能特性[3]。
乳酸菌指可以發酵乳糖產生乳酸的一類無芽孢、革蘭氏陽性細菌的總稱,主要包括雙歧桿菌屬、乳桿菌屬、鏈球菌屬和片球菌屬等約41 個屬[4-5],常被用作發酵劑以開發發酵食品,加速和控制發酵食品的發酵過程。乳酸菌產生的有機酸可以促進原料乳快速酸化,產生醋酸、乙醇、胞外多糖等風味化合物,賦予發酵乳制品各種理想的功能屬性,同時還可以延長食品貨架期。此外,乳酸菌因具有改善腸道菌群平衡[6]、降低膽固醇[7]、提高機體免疫力[8]等多種潛在的生物功能,已被廣泛用于乳品加工、健康食品、動物養殖以及活菌藥物開發,成為食品、農業和醫藥領域重要的微生物資源。
盡管乳酸菌具有諸多潛在的益生特性,但存在其在原料乳發酵過程中存在生長速度緩慢、菌體密度低、營養物質需求復雜等局限性,因此可以通過不同乳酸菌菌株混合培養的方式,使物種間相互刺激生長,以達到提高菌體生長速率、增加菌體密度、獲得目標產物等目的。本文主要介紹了發酵乳中不同乳酸菌菌株間的相互作用方式和作用機制以及不同菌株共培養對發酵乳制品感官特性、營養特性等產品特性的影響,為將乳酸菌更好地應用于實際提供一定理論基礎。
1 發酵乳中乳酸菌菌株間的相互作用
傳統發酵乳生產中,乳酸菌不僅同發酵過程中的環境有著密切聯系,乳酸菌間的生態學關系也十分復雜,不同種類的乳酸菌產生的各類代謝物,使各菌株間存在著多樣的互作形式,影響彼此的生長。將乳酸菌菌株共培養時,營養缺陷型菌株可以利用其他菌株產生的代謝產物達到加快自身生長的目的,但同時,共培養也會改變發酵微環境、刺激菌株沉默基因的表達,改變菌株的生存狀態,使某些菌株的生長繁殖受到抑制。乳酸菌間的互生作用、拮抗作用等對發酵乳最終風味的形成和新物質的產生有著深刻影響。
1.1 互生作用
1.1.1 保加利亞乳桿菌和嗜熱鏈球菌間的互生作用
保加利亞乳桿菌和嗜熱鏈球菌作為傳統的酸奶發酵劑被廣泛研究。二者間的互生作用主要體現在氨基酸和氮源利用方面(如圖1)。YONEZAWA 等[9]研究發嗜熱鏈球菌可以通過上調amiABCDE(負責轉運寡肽的操縱子)等方式獲取保加利亞乳桿菌降解蛋白后產生的組氨酸、脯氨酸、甲硫氨酸等氨基酸,從而促進自身的生長[10-11]。同時,嗜熱鏈球菌還可以通過脲酶調控二氧化物濃度從而影響保加利亞乳桿菌的酸化[12],并促進后者對天冬氨酸、谷氨酸、精氨酸以及核苷酸的合成速率[13]。
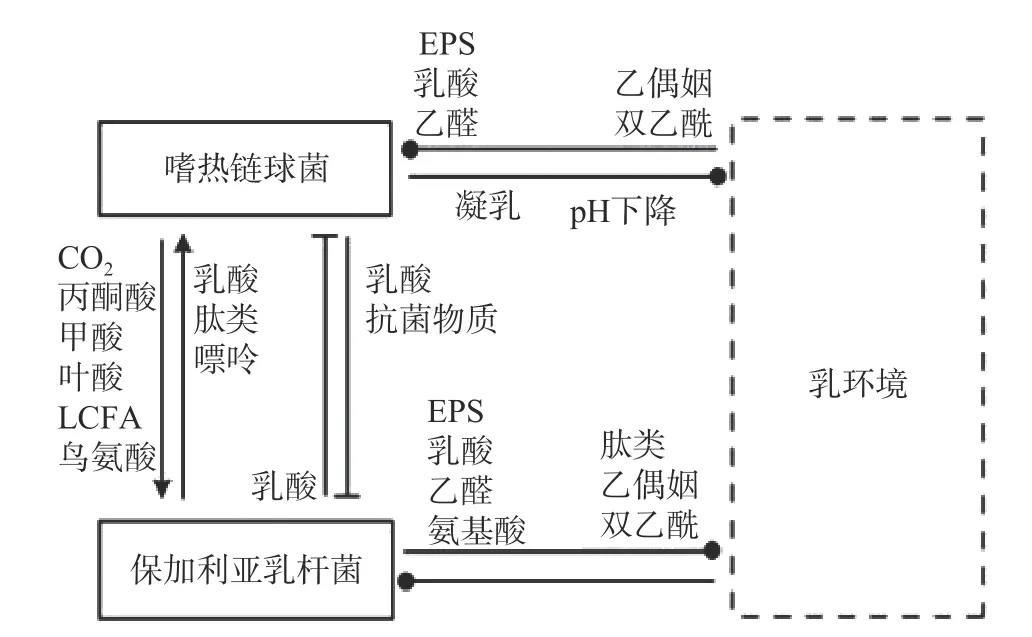
圖1 保加利亞乳桿菌和嗜熱鏈球菌在牛乳發酵中的相互作用[17]Fig.1 Interaction between Lactobacillus bulgaricus and Streptococcus thermophilus in milk fermentation[17]
其次,在氮源利用方面,由于保加利亞乳桿菌和嗜熱鏈球菌在共培養過程中,對不同的肽利用程度不同,從而提高了氮源的利用率。在二者共培養期間主要增加了C 末端含有焦谷氨酸的肽的相對濃度,這種肽主要被保加利亞乳桿菌轉運并被其多肽酶進一步降解以產生更多谷氨酸[14]。進入對數階段后,保加利亞乳桿菌更有效地運輸在N 末端含有半胱氨酸的肽,而嗜熱鏈球菌對這種肽的利用率較低。在環境肽利用方面,兩種菌株主要在同一時間點使用不同類型的肽,從而提高了每種菌株對必需肽的轉運效率[13]。最后,在核苷酸代謝方面,嗜熱鏈球菌可以為保加利亞乳桿菌提供甲酸以及嘌呤和氨基酸生物合成的輔助因子葉酸促進其生長[15-16]。
1.1.2 雙歧桿菌和干酪乳桿菌間的互生作用 雙歧桿菌能夠定殖于人體腸道中,通過代謝產生乙酸、乳酸等,降低腸道pH,進而抑制致病菌的生長,被廣泛應用于發酵乳制品的生產中[18]。干酪乳桿菌是乳酸菌中研究和應用最廣泛的益生菌,由于干酪乳桿菌能產生許多生物活性代謝物,因具有預防或治療與腸道微生物區系紊亂等疾病的健康功效而被廣泛研究。將其應用于發酵乳的制備時,可以賦予產品更好的感官特性和益生特性[19]。在干酪乳桿菌Zhang 與乳雙歧桿菌V9 生長與代謝機制的試驗中發現,乳雙歧桿菌V9 通過上調乳糖降解,丙酮酸代謝和維生素代謝來促進其快速生長,干酪乳桿菌Zhang 可能通過上調丙酮酸代謝與脂肪酸合成,下調糖酵解保持快速生長[20]。與單獨培養相比,在42 ℃條件下兩株菌共培養后乳雙歧桿菌V9 的活菌數增加了1.04 倍。
1.1.3 乳酸乳球菌和其他菌株間的互生作用 乳酸乳球菌可作為多種發酵乳制品的發酵劑,具有發酵乳糖、水解蛋白及產生胞外多糖的能力,是發酵乳制品最終香氣、質地和酸度形成的關鍵因素。發酵乳中乳酸乳球菌可以產生蛋白水解酶,并降解乳蛋白,為雙歧桿菌在脫脂乳中的生長提供寡肽、氨基酸等氮源,與雙歧桿菌共培養時通過氮需求的刺激性相互作用促進彼此的生長[21]。一項關于乳酸乳球菌MCC857與雙歧桿菌共培養的研究也發現乳酸乳球菌通過向后者提供氮源的方式刺激其生長[9]。但在CANON等[22]的研究中發現乳酸乳球菌NCDO2111 不能水解蛋白,將其與能進行蛋白水解的糞腸球菌CIRMBIA2412 共同培養時,乳酸乳球菌NCDO2111 生長速率提高,活菌數高達109CFU/mL,兩菌株之間顯示出強烈的相互作用。這是由于糞腸球菌為乳酸乳球菌NCDO2111 提供了較高濃度的色氨酸、纈氨酸、苯丙氨酸、亮氨酸、異亮氨酸和多肽,這些氨基酸和多肽可以加快乳酸乳球菌NCDO2111 的產酸速率,提高棉子糖利用率,增加揮發性化合物的濃度。
1.2 拮抗作用
發酵乳中不同乳酸菌菌株在共培養過程中也存在著拮抗作用。如將保加利亞乳桿菌與干酪乳桿菌混合培養后得到的實際菌體濃度為理論濃度的0.3倍[23]。劉學云等[24]通過研究九種益生菌之間的相互作用發現,三分之一的益生菌之間存在拮抗作用,其中嗜酸乳桿菌的代謝產物對植物乳桿菌、乳雙歧桿菌、副干酪乳桿菌有抑制作用;副干酪乳桿菌的代謝產物對嗜酸乳桿菌也有抑制作用;嗜熱鏈球菌的代謝產物產物對兩歧雙歧桿菌有抑制作用。
乳酸菌間的拮抗作用通常認為是由于不同菌株之間資源競爭或某些菌株產生了有害毒素而引起的[25]。盡管保加利亞乳桿菌和嗜熱鏈球菌共生現象普遍存在,但在LIU 等[13]的研究中發現嗜熱鏈球菌對肽的吸收能力強于保加利亞乳桿菌2038,在氮源利用方面二者也存在一定的競爭關系;另一方面,保加利亞乳桿菌2038 失去了優先利用環境中游離支鏈氨基酸(BCAA)的能力,在與嗜熱鏈球菌共培養時被誘導停止將天冬氨酸轉化為碳骨架中間體,使得共培養期間保加利亞乳桿菌的生長速度低于單菌株培養時的速度。因此并不是所有的保加利亞乳桿菌和嗜熱鏈球菌菌株之間都存在共生關系,還有一部分菌株之間可能存在拮抗作用[16]。
乳酸菌在發酵過程中會產生細菌素、胞外多糖、生物活性肽等抑菌化合物,具有抗炎、抗氧化等益生作用,但與其他微生物共培養時這些代謝產物也會抑制其它菌種的生長繁殖[23]。VAN 等[26]的研究表明,保加利亞乳桿菌產生的過氧化氫和細菌素等抑菌物質,破壞了其與嗜熱鏈球菌之間的平衡。由于產生細菌素而導致中間關系受到影響的還有保加利亞乳桿菌和嗜酸乳桿菌,在VINDEROLA 等[27]的研究中發現保加利亞乳桿菌的生長完全受到嗜酸乳桿菌CNRZ1881 的抑制,這是由于后者產生類細菌素導致的。了解這些拮抗作用為進一步探究乳酸菌間的作用機制以及開發有益生特性的產品奠定基礎。
2 互作機制
乳制品發酵過程中,不同乳酸菌菌株間的相互作用可以通過多種機制進行,如通過鞭毛等直接接觸的方式,或通過信號分子等相互作用,此外,乳酸菌在共培養時還會在介質中釋放一些可以共享的分子,如酶、生物表面活性劑等,這些可以被共用的分子稱為公共產品[28]。近幾十年來,這些互作背后的機制逐漸成為研究的焦點。
2.1 營養機制
發酵乳中的許多乳酸菌可以共同利用環境中的某些營養物質,如共享轉化酶、脂肪酶和蛋白水解酶等胞外水解酶,乳酸菌通過這些酶將“公共分子池”中直接可用的底物分解并利用。此外,乳酸菌也可以通過不同菌株之間的交叉喂養發生互生作用。交叉喂養,即一種微生物(稱為供體)吸收初級底物并將其轉化為作為“公共物品”排出,隨后由接受者使用的現象[28-30]。如雙歧桿菌通過為乳桿菌提供胞外多糖作為發酵底物和生長基質來提高乳桿菌的生物量[31]。保加利亞乳桿菌水解蛋白產生多肽后與嗜熱鏈球菌共享也被證明是兩種菌株發生相互作用的關鍵[32]。乳酸菌之間也存在營養物質的非特異性競爭,這些營養物質是發酵體系中微生物生長所必需的,其中優勢菌群會消耗其他菌種用于生長的基質,從而抑制它們的生長。
2.2 代謝產物作用機制
乳酸菌在發酵乳的生產過程中會產生代謝分泌物,對周圍環境中的其他微生物產生協同作用或拮抗作用。劉學云等[33]將嗜酸乳桿菌、嗜熱鏈球菌及干酪乳桿菌的無細胞上清液分別經高壓滅菌法和過濾法處理后與另一菌株共培養,結果顯示,三者之間存在協同共生作用,值得注意的是,與過濾除菌法相比,無細胞上清液經高壓滅菌法處理后再與其他菌株混合培養時活菌數明顯降低,這是由于上清液經高溫處理后菌株的部分代謝產物被熱破壞,菌株間的協同作用減弱,導致活菌數下降,說明乳酸菌的協同作用與其代謝產物有關。
乳酸菌產生的主要抗菌化合物是糖發酵過程中形成的有機酸,這會導致環境快速酸化,氧化還原位點降低,從而抑制其他微生物的生長。此外,乳酸菌也會產生細菌素、過氧化氫等抗菌化合物。乳酸菌產生的細菌素是核糖體合成的抗菌肽,具有綠色、安全的優點,是天然抗菌劑的重要來源[34]。細菌素主要作用于靶細胞的細胞膜,增加細胞通透性,使胞內物質被釋放,菌體細胞裂解死亡,如腸球菌產生的細菌素使單增李斯特菌胞內蛋白質類和核酸類釋放到細胞外環境中從而導致其死亡[35]。過氧化氫的抗菌活性與其強烈的氧化作用有關。在有氧存在條件下,益生菌通過輔酶Q 的作用進行電子轉移而產生過氧化氫,導致超氧化陰離子形成,破壞性氫氧游離基。這類超氧游離基對細菌細胞壁有強氧化作用,可破壞核酸及細胞內蛋白質從而影響細胞活性[36]。
2.3 信號分子形成機制
在乳制品發酵過程中,乳酸菌會形成一種群體水平上的相互作用,即群體感應。乳酸菌群體感應系統可以通過感知細胞濃度來調控特定的基因表達[37],并進一步調節乳酸菌的生物膜形成[38]、酸脅迫應答[39]、抗菌肽和胞外酶的合成[40-41]等重要的生理過程。乳酸菌自身會分泌被稱為信號分子的胞外小分子,信號分子能出入細胞并在環境中積累,隨著乳酸菌菌體密度的增加,信號分子在環境中的濃度也增加,當這些信號分子在乳酸菌的生長環境及細胞內的積累都達到一定的閾值時,就會被特異受體識別,啟動某些特定基因的表達,相關的蛋白合成上升,因此形成了種內或種間的相互作用,達到對自身或其他微生物的協同或抑制作用[42]。目前報道中,乳酸菌的信號分子有AIPII、LamD、PltA、IP-TX、PlnA 和IP-673 等[43]。
3 發酵乳中乳酸菌菌株相互作用對發酵乳制品的影響
乳酸菌有助于發酵乳最終感官、營養和健康特性的形成。隨著乳酸菌理論研究的不斷突破,其技術成果被加速轉化。發酵乳制品是乳酸菌的優良載體,以乳酸菌為發酵劑制備發酵乳制品已有上百年的歷史。對于傳統發酵過程,除發酵條件影響乳酸菌群落結構變化外,其群落結構內部的相互作用貫穿了整個發酵過程。乳酸菌菌群間的相互作用可以賦予發酵食品特殊的風味,也對發酵食品質量的穩定性有著很大的影響。
3.1 感官特性
乳酸菌產生的乳酸、醋酸等有機酸,氨基酸、雙乙酰、多肽、胞外多糖等芳香化合物,以及水解酶、細菌素和過氧化氫等物質為發酵食品提供了各種理想的感官特性。發酵乳中具有不同功能特性的乳酸菌菌株通過優勢互補,可以產生單菌株無法獲得的發酵產物,改善發酵乳的風味、質地、口感等感官特性。張筠等[44]將鼠李糖乳桿菌和嗜熱鏈球菌混合發酵,發現菌種產酸產黏性能良好,產品中乳酸菌活菌數增加,在短時間內即得到風味俱佳的酸花生乳。將瑞士乳桿菌LH-3 和嗜熱鏈球菌427 混合發酵后凝乳時間比瑞士乳桿菌LH-3 單獨發酵縮短了3.5 h,粘度增加1.8 倍,改善了發酵乳乳清析出和質構,此外,兩者混合發酵可以降低發酵乳中谷氨酸脫羧酶的活性,抑制乳糖的消耗及乳酸的產出,從而延緩了發酵乳的后酸化[45]。
發酵劑培養物與益生菌共發酵有助于產生乙醛和雙乙酰等理想的揮發性代謝物,改善酸奶的風味。當發酵乳中添加干酪乳桿菌Zhang 與發酵劑共培養可以增加己酸和3-羥基-2-丁酮等揮發性風味物質的產生[19]。通過比對不同菌株組合在貯藏過程中主要風味物質組分與含量的變化情況,發現多菌株發酵的發酵乳揮發性風味物質含量顯著高于單一菌株的平均水平[46]。在傳統乳制品的發酵過程中,微生物不僅與發酵環境有著密切聯系,不同菌株間的生態學關系也十分復雜,彼此間的協同作用或拮抗作用對發酵食品最終的感官特性有著深刻影響。
3.2 營養特性
盡管感官指標仍然是決定發酵乳制品質量的主要指標,但消費者對發酵乳的營養需求也是決定其未來發展、新產品開發和產品規模的重要因素。
發酵過程提高了乳制品的營養價值,增加了營養物質的生物利用率。傳統發酵劑嗜熱鏈球菌和保加利亞乳桿菌通常代謝乳糖的葡萄糖部分,并將半乳糖釋放到胞外介質中,導致酸奶中殘留的半乳糖和未發酵的乳糖水平較高,不利于乳糖不耐癥患者的消化吸收,而植物乳桿菌能夠有效地代謝乳糖和半乳糖,以乳糖為基礎將植物乳桿菌WCFS1 與基礎酸奶發酵劑共培養,結果表明,乳糖消耗完全,半乳糖代謝效率高,與傳統發酵工藝相比,添加植物乳桿菌WCFS1和酸奶發酵劑結合降低了乳制品的總糖含量,產品在外觀、質地和風味上也都得到了消費者的認可,具有發酵低糖酸奶的潛力[47]。KIMOTO-NIRA 等[48]將乳酸乳球菌和棉子乳球菌共培養用于發酵牛奶和植物基質混合物,非蛋白水解型棉子乳球菌能夠降解引起腸道不適的棉子糖、蜜二糖和水蘇糖,且當這兩種菌株一起生長時可以更好地酸化牛奶。
3.3 功能特性
發酵乳制品既保留了發酵乳本身的營養特性,又賦予了產品一定的益生特性。NISHIYAMA 等[49]通過雙盲對照實驗,發現含動物雙歧桿菌Bb12 和乳酸乳球菌11/19-B1 的酸奶能降低血清低密度脂蛋白的含量,并加強細胞免疫刺激功能。由青春雙歧桿菌B8589和副干酪乳酪桿菌PC-01 復合發酵的益生菌發酵乳飲料通過調控三羧酸循環和谷氨酸代謝途徑,顯著提升益生菌發酵乳飲料中γ-氨基丁酸和L-蘋果酸等生物活性物質的含量,且整個貯藏期(30 d)內,該復合益生菌發酵乳飲料的活菌數高于5×108CFU/g ,足量的益生菌及發酵乳飲料中大量的生物活性代謝物可共同賦予復合益生菌發酵乳飲料雙重的益生功效[50]。連續四周食用含有干酪乳桿菌Zhang 和動物雙歧桿菌乳酸菌亞種V9 的發酵乳后可以改善便秘患者排便頻次、排便時間和糞便黏稠度等癥狀[51]。人群隨機對照實驗表明,乳雙歧桿菌CNCMI-2494 發酵牛乳可以在2 周內快速改善腹脹、腹痛、腸胃脹氣等輕微消化不良癥狀,且癥狀改善與膳食纖維補充及運動無關[52]。
在長期的生產實踐中發現,對于成分或生化過程過于復雜的發酵來說,單一菌種發酵的局限性日益突出,而混合培養因菌種之間可以優勢互補,從而提高活菌數、縮短發酵周期,并有效改善產品特性,為發酵乳生產開啟了新的途徑。
4 總結與展望
近年來,乳酸菌越來越多的益生特性被揭示出來,發酵乳中乳酸菌菌株共培養在產量調控、產品功能化和資源利用等方面為發酵食品提供了新的機遇。但從目前的研究報道發現,由于菌株的多樣性和發酵底物的復雜性,發酵產物的確切組成和分子結構尚不完全清楚,大多數混合菌體系中菌株間相互關系和作用機制的研究也不夠深入,微生物混合培養研究更多地局限于工藝的優化。因此,在未來實際應用中,全面深入的了解發酵乳制品中乳酸菌菌株之間的相互作用方式及作用機制,探究微生物組的特定功能及其與風味、香氣和其他特性的關系,合理地利用乳酸菌資源,開發出更加高效、安全、且健康的乳酸菌產品對乳酸菌后續的開發與應用具有顯著的意義。
猜你喜歡
小讀者(2021年2期)2021-03-29 05:03:48
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
中國乳業(2018年3期)2018-04-13 01:05:08
食品與機械(2017年4期)2017-07-05 14:46:17
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
現代食品(2016年24期)2016-04-28 08:12:06
工業設計(2016年12期)2016-04-16 02:51:53
