浙江省銅山源水庫水體偏堿污染成因研究
2023-08-26 00:09:07錢旭蔡梅王元元白君瑞
人民長江 2023年8期
錢旭 蔡梅 王元元 白君瑞
摘要:
針對中國東南丘陵地區水庫出現不同程度的水質偏堿,部分水庫在局部時段內水體pH值指標頻繁超出地表水質量標準的問題,以浙江衢州大(2)型水庫銅山源水庫為例,實施水質監測和水生態調查。通過水文、氣象、水質、水生態指標之間的相關性分析,揭示了自然環境本底水質、關鍵氣象水文過程、外源污染以及藻類生長對水庫水質偏堿污染的影響機制。結果表明:銅山源水庫水體偏堿污染具有長期性的特征,且在每年的4~8月偏堿現象最為嚴重。水庫周邊自然環境下的本底水質偏堿,銅山源水庫入庫河流水體pH值高達8.02~8.47,局部時段高于地表水標準值。庫區水體pH值指標與水溫、DO、營養性物質(TP、TN)、葉綠素a等指標在一定程度上隨著時間變化呈現出同步升降的正相關性。pH值變化受多種因素的綜合影響,主要表現為直接或間接地促進藻類光合作用增強,導致光合作用效應大于水體水生動植物的呼吸作用,從而導致水體偏堿污染。
關 鍵 詞:
水庫污染; pH值; 藻類; 溶解氧; 葉綠素a; 銅山源水庫
中圖法分類號: X131.2
文獻標志碼: A
DOI:10.16232/j.cnki.1001-4179.2023.08.010
0 引 言
近年來,水庫逐漸成為城鎮生產、生活用水的重要來源之一,尤其是中國東南沿海省市,水庫型水源地日益成為經濟社會可持續發展的重要保障[1]。然而,中國地表湖庫近年來多發生水體偏堿污染現象,北方地區天津市的于橋水庫[2-3]、山東省的大沽夾河流域[4],東部地區上海市的東風西沙水庫[5],東南地區福建省的東牙溪水庫[6],南部地區的廣西大王灘水庫[7]、云南漁洞水庫[8]等,均出現過不同程度的水體pH值偏高現象。水庫作為一種天然或人工的湖泊,水質特點與天然地表湖泊相似。有統計表明:中國湖泊的pH值具有地帶性的分布特點[9],蒙新、青藏地區絕大多數呈現堿性或強堿性;云貴和黃淮海地區呈弱堿性;而東北及長江中下游地區地表湖泊pH值較低,一般在6.5~8.3之間,呈現中性或微堿性。在21世紀前10 a,浙江省部分水庫,如千島湖(新安江)水庫、老虎潭水庫、長潭水庫等,水體長期呈現中性偏堿性(pH值為6.70~9.17)[10]。2016年,浙江省水庫水質安全指標評價以Ⅰ~Ⅲ類為主,藻類大量生長的4~9月,部分水庫存在pH值超出地表水環境質量標準的現象[11]。位于浙江省衢州市境內的銅山源水庫作為衢江流域的大(2)型水庫,承載了浙西地區的數千萬公頃農田的灌溉和周邊地區水力供電及漁業和觀賞旅游功能,發揮著極其重要的作用[12]。銅山源水庫國控斷面監測數據顯示,2016~2020年銅山源水庫水體pH值在春末及夏季(4~8月)較地表水環境質量標準規定的pH值標準高值偏高(pH值為9)。
湖庫水體pH值受多種因素綜合影響,是水體中二氧化碳含量、有機酸和水體污染的表征指標[13],同時也是反映水體化學的一個重要指標,常被作為水體生態環境影響因素來考慮[14]。因此,此類酸堿平衡失調現象是水庫水生生態系統失衡的具體表現形式之一。目前,針對地表水體pH值超標的研究較少,國內一些研究認為,湖庫水體的pH值主要與水體中積累的營養鹽和藻類的繁殖有關[8],當水體中藻類數量上升到一定數量級時,其生物規模、生命活動的旺盛程度等必然對水體酸堿性變化起到主導作用[15-16]。黃歲樑等[17-18]研究指出在水體交換強的夏季和秋季,養殖和非養殖水體中pH值與溶解氧、葉綠素a濃度存在一定程度的正相關關系,王志紅等[15]指出晝夜pH值差值與藻類的生長態勢和生命活性之間存在一定相關性。
目前,國內外學者多聚焦于對浮游植物本身生長習性及其與營養物質等因素之間的關系研究,而浮游植物與水體pH值之間的關系尚未引起普遍關注,也未有權威的共識。針對中國東南沿海地區水庫水體偏堿污染現象研究也尚未全面開展,湖庫水體偏堿問題成因不明,現階段已嚴重阻礙國控、省控斷面水質穩定達標,亟需開展水體偏堿問題成因探究。此外,目前已有的研究多是基于室內研究和原位試驗,但作為實際地表水體的水庫是一個較為復雜的綜合水生態系統,水體酸堿性可能受到包括水文氣象因素、外源來水以及沉積物中的內源污染等在內的多種因素影響,更需要結合實際水質和水生態監測調查成果開展綜合性研究。因此,本研究針對東南沿海地區水庫普遍性偏堿問題,以浙江銅山源水庫為研究對象,采用現場水質監測和水生態調查,科學評價水庫水體的酸堿狀況,探究水庫水體偏堿污染成因,為水庫的水生態安全保障提供支撐。
1 材料與方法
1.1 研究對象概況
銅山源水庫(118°15′~119°E,29°~29°15′N)位于浙江省衢州市衢江流域(圖1),集水面積180 km2,總庫容1.715億m3,年平均入庫水量2.103億m3,換水周期約為148~220 d,是一座以農業灌溉為主,結合防洪、發電、潔水漁業、水資源保護等功能的大(2)型水庫,水質保護目標為Ⅱ~Ⅲ類。水庫所在區域屬亞熱帶季風性氣候區,受東南季風影響,有溫暖濕潤、四季分明、冬夏長春秋短、光照充足、降水豐沛而季節分配不均的地帶性特征。庫區雨量充沛,多年平均降雨量1 500~2 300 mm。降雨年內分配不均,主要集中在3~7月,其降水量占全年降水量的60%以上。
1.2 樣品采集
1.2.1 監測布點
根據HJ/T 91-2002《地表水和污水監測技術規范》中對于湖泊、水庫水質監測布點原則和方法的相關要求,結合水庫水深情況,在水庫進水區、出水區、深水區、淺水區、湖心區以及岸邊區相關位置布設6處(St1~St6)水質和水生態監測點位。其中,St6監測點為全庫水深最深處,水深達30~40 m,且處于水庫的國控斷面[19]附近。同時,在3條入庫河道(從西到東分別為銅山溪、廟前溪、白鶴山入湖河道)入庫口上游設置St7~St8兩處水質監測點。監測布點位置見圖2。
1.2.2 樣品采集
2021年4~8月,對St1~St8八處監測點實施水質加密監測,對St1~St6六處庫區點位開展水生態調查。其中,水質監測頻次為1旬1次;水生態調查在3、5、7月各開展1次。
此外,為進一步明晰庫區水質pH值和溶解氧指標的相關性,在4~8月每月各選取1 d開展全天(2 h間隔)的pH值和溶解氧樣本采集。為了分析庫區水體垂向pH值變化,選取深水區St6監測點,每月開展1次垂向分層采樣。
1.3 樣品處理與測定
(1) 水質指標測定。水體pH值、水溫、溶解氧、電導率在現場采用YSI Pro plus(美國)儀器測定總氮、氨氮、總磷、高錳酸鹽指數等其他水質指標取水樣冷藏運輸至實驗室,分別采用堿性過硫酸鉀紫外分光光度法、奈氏試劑分光光度法、鉬酸銨分光光度法和高錳酸鹽法測定[20];葉綠素a在采樣后2 h內,抽濾處理并用液氮(-180 ℃)保存后采用丙酮-分光光度法測定。
(2) 水生態指標測定。水生態調查內容主要包括浮游植物、浮游動物、底棲動物、魚類、水生植物及底質等。浮游植物、浮游動物、底棲動物的調查指標主要是密度、生物量、種類組成、優勢種;魚類調查指標主要是魚類種類組成、優勢種;水生植物主要調查水生植物種類組成、覆蓋度、優勢種。
1.4 數據處理與分析
為開展水庫水體pH值與其他水質指標之間的關聯性分析,引入灰色關聯度分析法(Grey Relation Analysis,GRA)。對于具備時間變化屬性的因子,在樣本數量不大的情況下,將數據進行歸一化處理、最值矯正后,通過計算數據之間的關聯度,度量兩組隨時間變化數據的變化走勢是否相近,以此判斷兩個因子之間的關聯性。灰色關聯度的取值范圍為[0,1],值越大的說明灰色關聯度越大[21]。
本研究采用WPS office中WPS表格進行數據的處理和灰色關聯度統計分析;采用Origin 2017進行分析制圖。
2 結果與討論
2.1 水庫水體偏堿污染特征
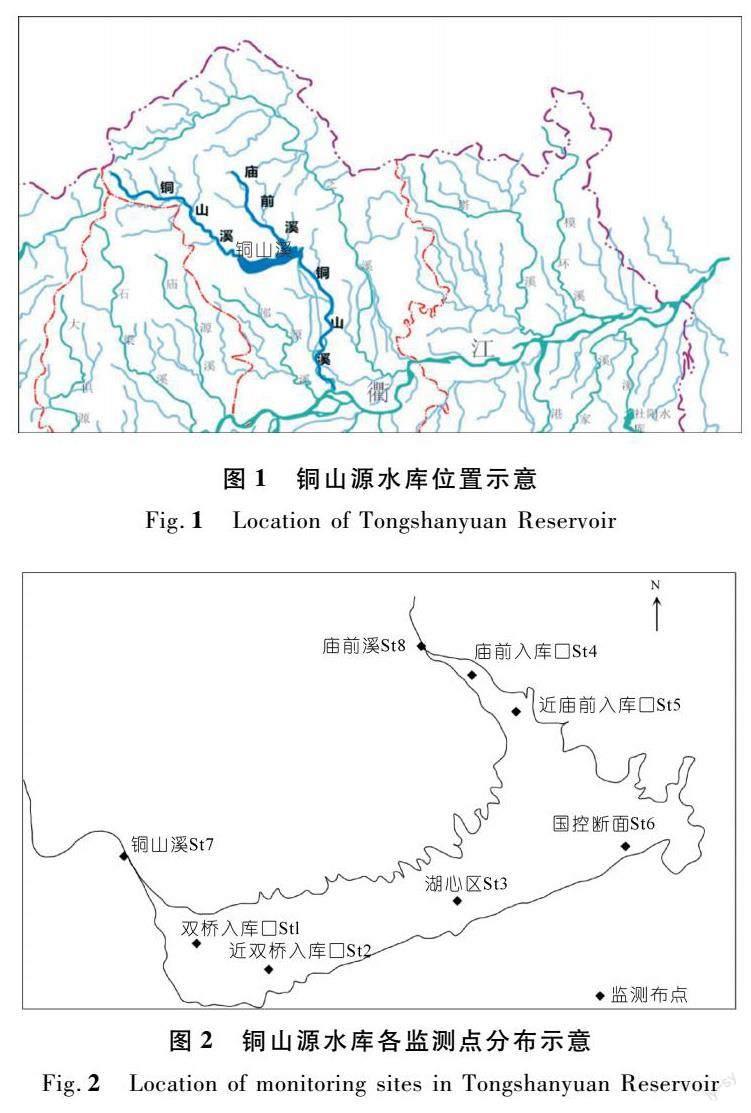
由2016~2020年銅山源水庫水體國控斷面每月pH值變化情況(圖3)可見,銅山源水庫水體偏堿現象在近5 a時間內中長期存在。主要呈現出秋冬季節水體pH值較低而春夏季節較高的特征,尤其每年的4~8月為水庫水體偏堿污染較為嚴重的時期。有調查指出,浙江省內其他水庫,如寧波市東錢湖、臺州市里石門水庫和長潭水庫等,也普遍存在著水體pH值超標現象[11]。
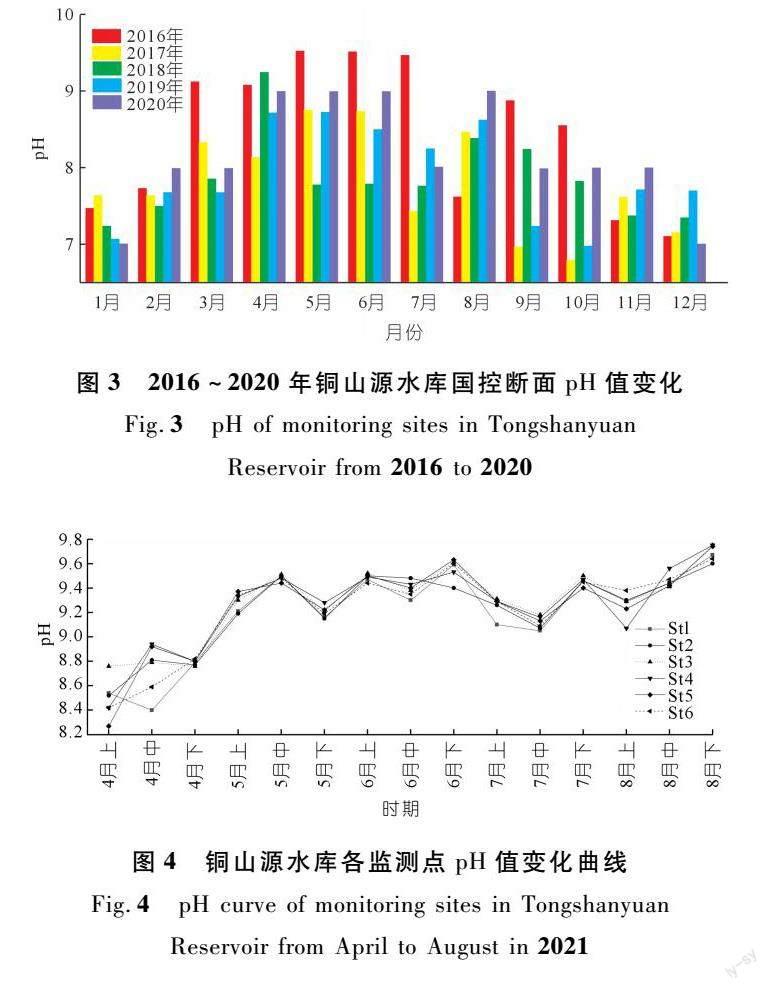
本研究選取銅山源水庫水體偏堿污染現象較為嚴重的4~8月,按旬開展水質加密監測,測得St1~St6各監測點水體pH值變化情況如圖4所示。4~8月銅山源水庫庫區內St1~St6監測點的pH值均呈現偏堿性,數值在8.27~9.75之間。時間尺度上,St1~St6各監測點在4月pH值較低(8.27~8.94),此后水體pH值呈現持續升高的態勢,5月上旬水庫水體pH值已高于地表水質量標準值(9),隨后進入持續性高位波動狀態,特別是6月全月和8月中下旬水體pH值普遍較高,在9.5上下波動,偏堿污染較為嚴重。空間尺度上,St1~St6各監測點的水體pH值變化趨勢基本一致,且同一時間各監測點pH值相差不大。總體而言,銅山源水庫水體在4~8月普遍呈現偏堿性,其中5~8月(夏秋季節)超過地表水環境質量標準,產生偏堿污染,全庫水質pH值的空間差異性較小。
2.2 自然環境本底水質特征
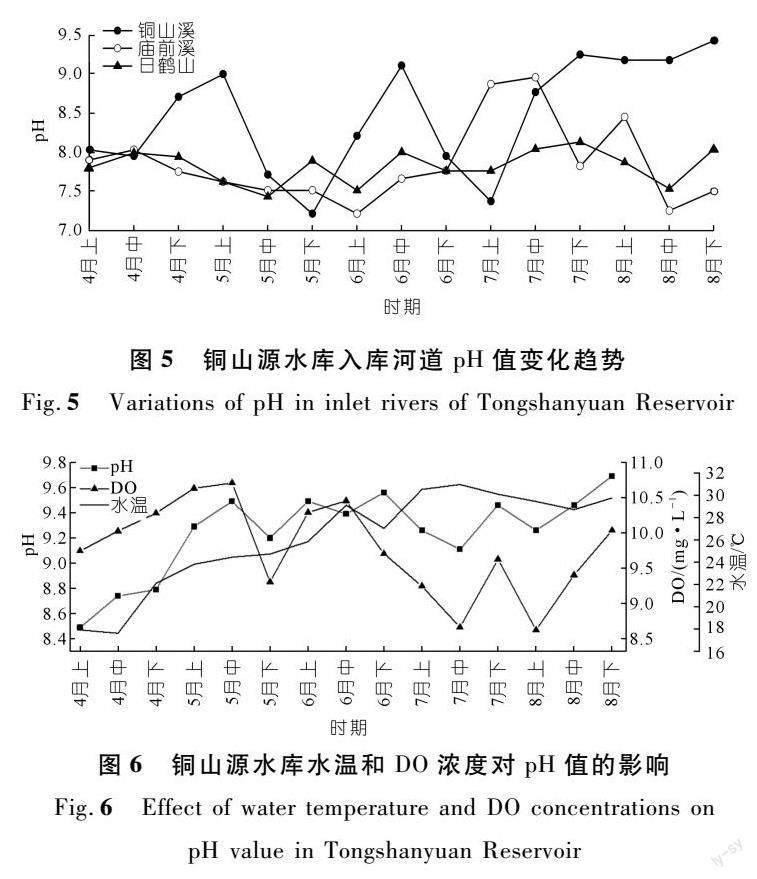
對銅山源水庫的入庫河道水質進行監測,以研究自然環境本底水質對水庫偏堿污染的影響,結果如圖5所示。白鶴山入庫河道(St5監測點附近)水體4~8月的pH值變化范圍在7~8之間,總體變化幅度較小。廟前溪和銅山溪水體的pH值變化幅度較大,其中,廟前溪水體pH值變化在7~9范圍內,銅山溪水體pH值變化在7.0~9.5范圍內;且銅山溪水體pH值在7~8月間總體維持在9以上的高位,已超過地表水水質標準。由此可知,銅山源水庫附近自然河道中流動性本底水質已呈現較為明顯的偏堿性,在7~8月一度超過地表水環境標準值上限。因此,當河道中的水體進入水庫中成為水庫水體的一部分,必然導致水庫水體本底pH值偏高。
2.3 關鍵氣象水文過程對庫區水體偏堿污染的影響
2.3.1 氣象因素特征
氣溫和光照等氣象因素會對地表水體水質產生一定間接影響。一般而言,氣溫的影響主要通過水溫反映,光照主要通過影響綠色植物的光合作用影響水質狀況,而植物光合作用一般采用水體中的溶解氧濃度來表征。因此,本研究主要通過庫區水體的水溫以及溶解氧濃度來分析氣象因素對庫區水體偏堿污染的影響。
由圖6可知,pH值和DO濃度曲線隨著時間變化基本呈現同步升降的趨勢,因此水體pH值和DO濃度兩項指標之間存在較為明顯的正相關性。灰色關聯度分析計算表明,pH值和DO濃度之間的灰色關聯系數為0.89,具有較為顯著的關聯關系,水庫水體pH值受到DO濃度的影響。本次水生態調查結果顯示,銅山源水庫中不存在相當規模的水生植物群落生長跡象,因此水庫水體DO濃度和pH值受到水庫中浮游植物(主要為藻類)生長的影響。
從pH值和水溫曲線的變化趨勢可知,4~8月水體水溫呈現逐步上升趨勢,而同時水體pH值總體呈現波動上升趨勢,說明水溫和pH值具有同向變化的特征。灰色關聯度分析顯示:pH值和水溫之間的灰色關聯系數為0.83,兩者具有較為顯著的正相關關系。有研究指出,水溫變化總體可從3個方面影響水體中pH值變化:① 水溫升高一定程度上促進水生生物的呼吸作用[22],致使水體向偏酸性方向發展;② 在適當范圍內水溫逐步升高可促進增加水體中藻類的生物量和優勢度,促進其光合作用,使水體向偏堿性方向發展[23~25];③ 水溫升高促進CO2氣體分子運動速率加快,容易自水面逸出,致使水體向偏堿性方向發展[26]。由此可知,水溫對于水體pH值的影響主要通過促進水體中藻類生長繁殖,影響光合作用和呼吸作用的強度來實現。
從各特征時間段來看:① 4月至5月中下旬,水體水溫、DO與pH值3項指標逐漸同步升高,表明藻類的光合作用不斷增強、水體pH值也不斷升高;此時期內水溫處于20~25 ℃范圍內,水生態調查發現水庫中優勢藻種依次為4月初的硅藻門小環藻和5月的藍藻門微囊藻,表明隨著水溫的變化,水庫中適應性的藻類群落開始生長演替。② 6月上中旬,水溫由25 ℃逐步升高至30 ℃,適宜藍綠藻生長,同時DO濃度充足,藍綠藻類生長旺盛、光合作用較強,pH值呈現高位波動態勢。③ 7月中旬,水體水溫在30~32 ℃范圍內變動,過高的水溫一定程度上抑止了藻類生長,pH值與DO出現同步降低趨勢。④ 7月中下旬至8月期間,水溫維持在27~30 ℃范圍內,適宜藻類生長繁殖,此時期內水體pH值逐漸出現波動上升趨勢。Reynolds等人研究指出,環境因子對浮游生物類型季節性豐度和多樣性的影響差異顯著,其中,溫度、光照強度等物理因子較為重要[27]。因此,從水溫、DO與水體pH值的變化關系分析可知,水溫和DO濃度通過影響藻類的生長間接影響水體pH值變化趨勢。王斌[28]、吳阿娜[29]等亦指出,湖庫水體葉綠素a與水溫、pH值、溶解氧呈顯著正相關。
2.3.2 關鍵水文過程特征
降雨以及徑流等關鍵水文過程同樣可能對水體水質產生影響。本研究統計了2016~2021年5~8月水庫降雨、出入庫水量情況,如表1所示。由表1可知,2021年5~8月水庫周邊累積降雨量為1 206.90 mm,較2016~2020年同期累計降雨量均值(920.38 mm)多。2021年5~8月累積入庫徑流量14 634.50萬m3,較2016~2020年同期入庫徑流量均值(11 168.61萬m3)大;累積出庫徑流量17 744.32萬m3,與2016~2020年同期出庫徑流量均值(17 709.59萬m3)相當。從凈出庫水量來看,2021年5~8月累積出庫凈流量3 109.82萬m3,較2016~2020年同期累積出庫凈流量(6 540.97萬m3)減少了52.5%。因此,2021年5~8月相比于2016~2020年同期,總體存在著庫區周邊降雨較往年偏多、入庫水量較往年偏多、出庫水量與往年持平、凈出庫水量較往年大幅減少的現象。
多項研究指出,出入庫徑流以及湖庫本身水動力條件顯著影響浮游植物的生長。浮游植物的豐度及結構動態變化與溫度、光照、水體穩定性、水力滯留時間等因素時空變化的結果相關[30-32]。也有研究指出,與銅山源水庫位于相同緯度帶的浙江省新安江水庫,浮游植物群落結構在很大程度上受水溫、氣象條件影響[33]。水力停留時間影響浮游植物生長[1~2],在光合作用強烈的靜水或流速小于0.3 m/s的水體中,光合作用引起的溶解氧變化,必然引起水質pH值變化[34]。銅山源水庫出庫水量偏少的現象容易造成水庫水體流動緩慢,延長了水庫中水流的水力停留時間,適宜的水溫、光照、溶解氧等因素為水體中藻類的生長繁殖提供有利條件,從而促進了藻類的生長和光合作用強度,影響水庫水體的酸堿性,與上述研究結果一致。
2.4 藻類生長狀況及其對水庫水質偏堿污染的影響
從關鍵氣象水文因素分析可知,藻類生長繁殖所引起的光合作用強度遠超水生動植物呼吸作用強度,最終導致水體中DO濃度上升,可能是水庫偏堿污染現象產生的原因之一。2020年,生態環境部太湖流域東海海域生態環境監督管理局對浙閩片的湖庫進行了水環境水生態情況調查,發現在藻類大量生長的4~9月,浙江省部分水庫存在pH值超標的現象[11]。目前,國內外已有多項研究發現,水體中的pH值和藻類生長情況具有顯著相關性[10,24-25,35-36]。有多名學者提出可以采用pH值指標來作為藻類生長和繁殖的指示和預警指標。浮游植物主要受光照、營養物質、溫度、群落結構、生活史、分層或垂直混合和潮汐等因素的影響[37]。本研究擬從藻類生物量、水體營養物質含量以及藻類光合作用強度等方面探究藻類生長對水庫水體偏堿污染的影響。
2.4.1 藻類生物量
4~8月,庫區水體葉綠素a(Chl.a)、藻類生物量以及pH值隨時間變化特征呈現較為明顯的相似性和同步性(圖7)。灰色關聯度分析計算表明,pH值和葉綠素a濃度之間的灰色關聯系數為0.63,具有一定程度的正相關關系,由此可知水體偏堿污染與水體中藻類生長狀況密切相關。4~5月中旬,葉綠素a濃度與pH值同步波動上升,此時藻類生物量為1.59 mg/L;5月下旬pH值局部降低,藻類生物量和葉綠素a同步降低為0.84 mg/L和5.5 μg/L;6~7月,水溫長期保持在25~30 ℃,藻類生物量增至3.06~3.91 mg/L,葉綠素a濃度增至7~13 μg/L,此時pH值達到9.0~9.6;8月中下旬,pH值上升至9.4~9.8,此時水體中葉綠素a濃度也同步升至12~18 μg/L,上升幅度較大。因此,銅山源水庫中藻類群落的生物量對水體pH值產生影響。
此外,監測了3條入庫河道中葉綠素a濃度變化趨勢,見圖8。由圖可知,白鶴山入庫河道葉綠素a濃度在監測期內一直極小;廟前溪和銅山溪水體中葉綠素a濃度在7~8月顯著升高,尤其是銅山溪水體中葉綠素a濃度在8月末達到最高值23 μg/L,說明7~8月份銅山溪和廟前溪河道水體出現藻類快速繁殖現象,在一定程度上,可能對水庫水體中藻類生長繁殖產生一定程度的累加作用。
從銅山源水庫藻類群落演替過程來看,4月硅藻為優勢種,5月水庫中藍藻為優勢種,6月藍藻和綠藻相當,7月開始則是綠藻為優勢種。由此可知,水庫中的綠藻在6月初步開始繁殖,至7~8月已成為能夠提供大量葉綠素a用于光合作用的藻類群體,因此也極大地促進了7~8月份水體pH值的堿性發展。
2.4.2 營養物質
一般而言,增加的營養負荷被認為是藻類尤其是藍藻優勢的驅動因素[38],一定條件下營養物質相比于溫度等氣象因素,對藻類生長產生的影響更大[39]。藻類在湖泊中大量繁殖,導致水體環境中光合作用強度大于呼吸作用,促進水體偏堿發展。4~8月,TP和TN濃度與pH值變化關系曲線如圖9所示。
由圖9可知,pH值和TP濃度隨時間的變化曲線具有較為明顯的升降趨勢同步性,同時灰色關聯度計算顯示,pH值和TP、TN濃度之間的灰色關聯系數分別為0.67和0.69,具有一定程度的正向相關關系。可推斷總磷為水庫中的浮游植物尤其是藻類提供營養物質,間接對水庫水體的酸堿性產生影響。同時,庫區水體的TP濃度較低,總體維持在Ⅱ類水以內,僅有局部監測時段在Ⅱ~Ⅲ類水之間。由此可知,水庫中TP濃度并未完全滿足藻類生長的需求,隨著TP濃度的升降,藻類生長(葉綠素a指標)表現出極其同步的升降變化,TP指標可認為是銅山源水庫中藻類生長的限制性指標。
由圖9可知,pH值和TN濃度隨時間的變化曲線之間并未產生明顯相關性。王斌等人[28]亦指出,葉綠素濃度與TN濃度相關性較小。同時,水庫的TN濃度在Ⅲ類標準以內(相比TP指標,TN含量較高),可知TN濃度對于此階段內的藻類生長尚未產生限制作用,充足的氮源為藻類生長提供了豐富的養料。
研究指出,降雨和徑流中的養分對湖泊、水庫的養分負荷產生一定影響[40~42]。銅山源水庫上游地區的外源污染主要通過入庫河道匯入,監測3條入庫河道總磷和總氮濃度變化趨勢如圖10所示。由圖可知,廟前溪水體TP濃度遠大于銅山溪和白鶴山入庫河道。對于總氮指標來說,銅山溪和廟前溪總體呈現波動趨勢,其濃度值變動范圍在0~1.43 mg/L范圍內(Ⅰ~Ⅳ類),白鶴山入庫河道的總氮濃度在監測時段內普遍較高,最高時可達2.36 mg/L,屬劣Ⅴ類水質。因此,從營養性物質上來看,3條入庫河道中白鶴山的總氮污染較嚴重;廟前溪的總磷污染較嚴重。氮磷類營養物質進入庫區,會為水庫中藻類提供大量營養,促進其生長和繁殖,從而導致水體偏堿。
2.4.3 藻類光合作用強度
有研究指出,在富營養化的地表水中,由于光合作用產生的O2量遠遠大于呼吸作用所需要的O2量,DO主要來源于藻類的光合作用,而光合作用引起的DO變化,必然引起水體pH值的變化[34]。有研究指出,藻類光合作用可快速減少CO2,使HCO-3的解離平衡偏移,促使水中HCO-3分解,OH-濃度或活度增加,pH值升高[4]。Trivedi[43]研究指出,湖庫水體高pH值可能是由于微觀和宏觀植被的高光合作用,使平衡向堿性方向移動。由前文分析可知,水庫水體pH值和DO總體呈現出消長同步的變化特征,兩者存在較為明顯的正相關關系。一般而言,在藻類生長旺盛的水體中,pH值和溶解氧因主要受藻類晝夜間交替進行的光合作用與呼吸作用的影響而呈現明顯的日變化規律,即pH值和溶解氧在早晨時最低,日出后整個白天內,從最小值逐漸增加,至日落前達到最大[17-18]。因此,選取水庫中3~4個監測點開展了pH值和溶解氧兩個指標的晝夜變化監測,探究水庫中藻類光合作用對水體偏堿污染的影響。
7月水庫各監測點的pH值和溶解氧指標晝夜變化曲線如圖11所示。雙橋入庫口(St1)、湖心區(St3)、廟前入庫口(St4)以及國控斷面(St6)的水體pH值和溶解氧指標隨時間均呈現類正態分布的變化趨勢,且兩指標的消長基本同步。其中雙橋入庫口附近水體在夜間20:00左右pH值和溶解氧指標達到最高值,其余位置水體在傍晚16:00和17:00間達到最高,與藻類光合與呼吸作用晝夜變化的時間特征吻合程度較高。
8月水庫各監測點的pH值和溶解氧指標晝夜變化曲線如圖11所示。由圖可知,8月份的pH值和溶解氧指標晝夜變化情況與7月份極為相似,兩者都呈現較為明顯的先升后降情況,與藻類光合與呼吸作用晝夜變化的時間特征吻合程度較高。
2.5 銅山源水庫水體偏堿污染形成機制
通過對銅山源水庫的水文氣象條件、水體pH值指標與其他水質指標之間的相關性分析可知,銅山源水庫水體偏堿問題是在多種因素共同影響下形成的一種綜合外在表現。首先,銅山源所在地區的大環境本底水質呈現偏堿的現象,尤其是入庫河道銅山溪水質呈現明顯超標現象(pH>9),根據水庫多年統計數據,銅山溪的年入庫徑流量在水庫總入流量中所占比重高達60%~70%,因此銅山溪河道水質偏堿是水庫水體偏堿污染的原因之一。從2.3節和2.4節分析可知,銅山源水庫水體pH值與水溫、DO、葉綠素a、藻類生物量以及TP濃度等指標均有著良好的正相關關系。有研究指出,水體中CO2含量會受到藻類光合作用、水生生物呼吸作用等的影響[17]。一般來說,水生生物呼吸作用產生CO2,而光合作用消耗水體中的CO2。光合作用所需的CO2一部分來自于大氣,一部分來自于水中HCO-3的解離(HCO-3=OH-+CO2),HCO-3在磷酸酐酶的催化作用下變化為H2CO3,然后經脫水反應生成CO2后被利用,在HCO-3變成H2CO3的過程中,電離出OH-。因此,光合作用使水體偏堿發展,而呼吸作用的結果則使水體向偏酸方向發展。一旦光合作用的效應大于呼吸作用,則水體中堿性物質的產生速率大于酸性物質的產生速率,水體即發生偏堿污染現象。4~8月間銅山源水庫水體pH值、水溫、DO、TP濃度等指標均與葉綠素a、藻類生物量呈現正相關關系,而且水庫中存在著較為穩定的水動力條件,說明水庫中藻類生長繁殖較為旺盛,光合作用較強,影響了水體中pH值、DO等物化指標的變化。這與游亮等人的研究成果一致,當藻類數量增加到一定數量級時,其數量的多少和生命活動的旺盛程度對水體的pH值起主導作用[44]。監測所得的7~8月pH值、溶解氧晝夜變化關系也恰好證明了這一點。而同時較高的pH值有利于水華藻類的生長,大量藻類繁殖又進一步提高pH值[45],形成循環反饋的機制,在一定程度上加重水質偏堿污染。
總體來說,銅山源水庫水體偏堿污染形成機制為:上游入庫河流本底水質偏堿,同時夏季藻類生長、光合作用速度遠遠大于水生動植物的呼吸作用,導致水體pH值在夏秋季節進一步上升,使水體偏堿污染嚴重。
3 結 論
本文通過開展實地水質監測和水生態調查,揭示了自然環境本底水質、關鍵氣象水文過程、外源污染以及藻類生長等對銅山源水庫水質偏堿污染的影響機制,得出水庫水質偏堿現象成因主要有兩個方面。
(1) 銅山源水庫周邊自然環境內的水體均呈現不同程度的堿性,本底水質偏堿是造成水庫水質偏堿的重要外因。
(2) 藻類生長和繁殖所產生的光合作用效應大于水體中水生動植物的呼吸作用,是造成水庫水質偏堿的內部原因。其中,春夏季節的水溫、光照、溶解氧、氮磷類營養物質以及相對穩定的水流條件等客觀因素,為藻類生長提供了穩定的生長環境和充分的光合作用原料,有力促進了藻類生長繁殖,間接促進水庫水體水質朝著偏堿的方向發展。
近年來,中國地表湖庫出現不同程度的水體偏堿污染現象。本研究揭示的地表湖庫水體偏堿問題成因,可為出現此類污染現象的湖庫治理提供理論支撐。
參考文獻:
[1] 馬沛明,施練東,張俊芳,等.浙江湯浦水庫浮游植物季節演替及其影響因子分析[J].環境科學,2016,37(12):4560-4569.
[2] 朱海燕,戴學穎,王可玉.于橋水庫pH值變化原因分析及可調控措施.[J].水科學工程與技術,2010(1):40-43.
[3] 秦保平,孫韌,王德龍,等.引灤河道中水質偏堿的起因研究[J].環境科學研究,1999,12(5):38-42.
[4] 林影,徐家明,張廣譜,等.煙臺市夾河流域pH偏高的原因[J].山東環境,1995,2:12-13,45.
[5] 施章薇.東風西沙水庫原水pH值升高及出廠水鋁控制方法研究[J].建材與裝飾,2019(4):33-34.
[6] 邊歸國.東牙溪水庫偏堿性污染的成因研究[J].福建環境,2003,20(6):15-17.
[7] 李建民,毛小英.大王灘水庫溶解氧及pH變化規律與原因分析[J].廣東水利水電,2012(12):37-40.
[8] 申開旭.漁洞水庫pH值堿化成因分析及控制[J].環境科學導刊,2011,30(1):47-49.
[9] 王蘇民,竇鴻身.中國湖泊志[M].北京:科學出版社,1998.
[10] 米瑋潔,俞建軍,陳暉,等.浙江飲用水源地水環境狀況及存在問題分析[J].人民長江,2012,43(13):95-98.
[11] 生態環境部太湖流域東海海域生態環境監督管理局.浙閩片主要湖庫基本情況調查與分析報告[R].上海:生態環境部太湖流域東海海域生態環境監督管理局,2020.
[12] 鐘衛鴻,單劍鋒,薛浚,等.氮和磷對銅山源水庫優勢藻生長影響實驗研究[J].環境污染與防治,2003,25(1):20-22.
[13] 李洪遠.生態恢復的原理與實踐(第一版)[M].北京:化學工業出版社,2005:15-18.
[14] 李錦秀,杜斌,孫以三.水動力條件對富營養化影響規律探討[J].水利水電技術,2005,36(5):15-18.
[15] 王志紅,崔福義,安全,等.pH與水庫水富營養化進程的相關性研究[J].給水排水,2004,30(5):37-41.
[16] 胡艷娟.水庫藻類光合作用對pH和溶解氧的影響[J].黑龍江水利科技,2011,39(2):17-18.
[17] 黃歲樑,臧常娟,杜勝藍,等.pH、溶解氧、葉綠素a之間相關性研究Ⅰ:養殖水體[J].環境工程學報,2011,5(6):1201-1208.
[18] 黃歲樑,臧常娟,杜勝藍,等.pH、溶解氧、葉綠素a之間相關性研究Ⅱ:非養殖水體[J].環境工程學報,2011,5(8):1681-1688.
[19] 國務院.水污染防治行動計劃[Z].北京:國務院,2015.
[20] 國家環境保護總局,水和廢水監測分析方法編委會.水和廢水監測分析方法(第四版)[M].北京:中國環境科學出版社,2002,223-226,243-257,276-285.
[21] 譚學瑞,鄧聚龍.灰色關聯分析:多因素統計分析新方法[J].統計研究,1995,12(3):46-48.
[22] 祁峰,李曉東,趙艷紅,等.鹽度、光照度和溫度對漂浮剛毛藻光合作用的影響[J].大連水產學院學報,2008,23(5):382-386.
[23] PEPERZAK L.Climate change and harmful algal blooms in the North Sea[J].Acta Oecologica-International Journal of Ecology,2003,24(增1):139-144.
[24] PAERL H W,HUISMAN J.Blooms like it hot[J].Science,2008,320:57-58.
[25] O′NEIL J,DAVIS T,BURFORD M,et al.The rise of harmful cyanobacteria blooms:the potential roles of eutrophication and climate change[J].Harmful Algae,2012,14:312-334.
[26] 劉輝,姬泓巍,辛梅.膠州灣水體中的二氧化碳體系[J].海洋科學,1998,1(6):44-47.
[27] REYNOLDS,C S.Phytoplankton periodicity:The interactions of form,function and environmental variability[J].Freshwater Biology,1984,14(2):111-142.
[28] 王斌,馬健,王銀亞,等.天山天池夏季葉綠素a的分布及富營養化特征研究[J].環境科學,2015,36(7):2466-2471.
[29] 吳阿娜,朱夢杰,湯琳,等.淀山湖藍藻水華高發期葉綠素a動態及相關環境因子分析[J].湖泊科學,2011,23(1):67-72.
[30] BECKER V,CAPUTO L,ORDóNEZ J,et al.Driving factors of the phytoplankton functional groups in a deep Mediterranean reservoir[J].Water Research,2010,44(11):3345-3354.
[31] VOGT R J,SHARMA S,LEAVITT P R.Decadal regulation of phytoplankton abundance and water clarity in a large continental reservoir by climatic,hydrologic and trophic processes[J].Journal of Great Lakes Research,2015,41:81-90.
[32] INEKE V G,JEROEN V W,KATLEEN V G,et al.Covariation between zooplankton community composition and cyanobacterial community dynamics in Lake Blaarmeersen(Belgium)[J].FEMS Microbiology Ecology,2008,63(2):222-237.
[33] 笪文怡,朱廣偉,吳志旭,等.2002~2017年千島湖浮游植物群落結構變化及其影響因素[J].湖泊科學,2019,31(5):1320-1333.
[34] 張澎浪,孫承軍.地表水體中藻類的生長對pH值及溶解氧含量的影響[J].中國環境監測,2004,20(4):49-50.
[35] 徐恩兵,孫健,韋雷.黑臭河道中的藻類光合作用對pH和溶解氧的影響.[J].環境工程,2018,36:78-79.
[36] 許霄霄,王靖偉.四川省邛海水體富營養化評價及建議[J].水利水電快報,2021,42(8):83-87.
[37] ALVAREZ-GNGORA C,HERRERA-SILVEIRA J A.Variations of phytoplankton community structure related to water quality trends in a tropical karstic coastal zone[J].Marine Pollution Bulletin,2005,52(1):48-60.
[38] BROOKES J D,CAREY C C.Resilience to blooms[J].Science,2011,334:46-47.
[39] THACKERAY S,JONES I,MABERLY S.Long-term change in the phenology of spring phytoplankton:species-specific responses to nutrient enrichment and climatic change[J].Journal of Ecology,2008,96:523-535.
[40] JEPPESEN E,KRONVANG B,MEERHOFF M,et al.Climate change effects on runoff,catchment phosphorus loading and lake ecological state,and potential adaptations[J].Journal of Environmental Quality,2009,38:1930-1941.
[41] JEPPESEN E,KRONVANG B,OLESEN J E,et al.Climate change effects on nitrogen loading from cultivated atchments in Europe:implications for nitrogen retention,ecological state of lakes and adaptation[J].Hydrobiologia,2011,663:1-21.
[42] MOSS B,KOSTEN S,MEERHOF M,et al.Allied attack:climate change and eutrophication[J].Inland Waters,2011(1):101-105.
[43] TRIVEDI,R.K.Limnology of three fresh water ponds in Manglore[C]∥National Symposium on Advances in limnology conservation of endangered fish species.Srinagar,Garhwal:Narendra Pub.House,1989,23-25.
[44] 游亮,崔莉鳳,劉載文,等.藻類生長過程中DO、pH與葉綠素的相關性分析[J].環境科學與技術,2007,30(9):42-44.
[45] 申開旭,申時斌,宋大恩.漁洞水庫pH值異常成因分析及水資源保護建議[J].環保科技,2014,20(1):34-38,45.
(編輯:李 慧)
Abstract:
In recent years,reservoirs in the hilly areas of southeastern China have been polluted by meta-alkalization in varying degrees,and the pH values of some reservoirs have frequently exceeded the environmental quality standards of surface water during some periods of time.Taking Tongshanyuan Reservoir,a large (second) type reservoir in Quzhou City,Zhejiang Province as an example,water quality monitoring and water ecology investigation were conducted,and the correlation analysis among hydrology,meteorology,water quality and water ecology indicators were combined to reveal the influence mechanism of background water quality of the natural environment,the key meteorological and hydrological processes,the exogenous pollution and the growth of algae on the meta-alkalization pollution of the reservoir.The results showed that the meta-alkalization pollution in Tongshanyuan Reservoir had lasted for a long time and was more serious from April to August every year.The background water of the natural environment around the reservoir was alkaline.The pH values of the inflow rivers were as high as 802 to 847,which was higher than the surface water standard in some time.The pH values of water in reservoir showed a relatively positive correlation to water temperature,DO,nutrients(TP、TN),chlorophyll-a to a certain extent,which rose and fell in synchronization with the change of time.Changes in the pH value of the water body were influenced by a variety of factors,which were mainly manifested in the direct or indirect enhancement for algal photosynthesis,leading to a greater effect of photosynthesis than respiration of aquatic organisms,which resulted in meta-alkalization and pollution of the reservoir water body.
Key words:
reservoir pollution;pH;algae;DO;chlorophyll-a;Tongshanyuan Reservoir
猜你喜歡
環境(2023年5期)2023-06-30 01:20:01
小學科學(學生版)(2021年7期)2021-07-28 06:44:32
小學科學(學生版)(2020年6期)2020-08-13 07:47:12
當代水產(2019年1期)2019-05-16 02:42:04
當代水產(2019年3期)2019-05-14 05:42:48
當代陜西(2019年7期)2019-04-25 00:22:18
領導決策信息(2018年26期)2018-10-12 02:18:26
電子制作(2018年14期)2018-08-21 01:38:16
水利規劃與設計(2016年7期)2016-02-28 15:06:27
都市麗人(2015年5期)2015-03-20 13:33:49
