釀酒葡萄鐵調節轉運蛋白基因VvIRT1的克隆、表達與功能
2023-09-01 10:41:56宋志忠徐維華肖慧琳唐美玲陳景輝管雪強劉萬好1
生物技術通報 2023年8期
關鍵詞:植物
宋志忠 徐維華 肖慧琳 唐美玲, 陳景輝 管雪強 劉萬好1,,
(1. 魯東大學農林工程研究院 山東省高校作物高產抗逆分子模塊育種重點實驗室,煙臺 264025;2. 山東省煙臺市農業科學研究院,煙臺265500;3. 山東省釀酒葡萄與葡萄酒技術創新中心 中糧長城葡萄酒(蓬萊)有限公司,煙臺 264000;4. 劍橋大學植物系,英國劍橋 CB23EA)
鐵(Fe)是植物細胞中含量較為豐富的礦物元素之一,參與植物光合作用、呼吸作用、激素合成、能量代謝和DNA修復等多種代謝途徑和生命過程[1-4]。土壤中的鐵多為Fe3+,植物難以直接吸收和利用,特別是石灰性土壤缺鐵現象更為明顯,土壤缺鐵造成作物嚴重減產或品質降低[5-9]。
Romheld等[10]最早提出了高等植物為適應缺鐵脅迫而進化出的兩種根際鐵吸收機制:吸收機制制I,即通過定位于根細胞膜表面的H-ATP酶向根際分泌H+,降周圍土壤pH,促進Fe3+的溶解,并通過鐵還原酶(ferric reduction oxidase, FRO)將根系周圍的Fe3+被還原為Fe2+,再通過鐵調節轉運蛋白(iron-regulated transporter, IRT)進入根系被植物吸收利用,常見于雙子葉植物和非禾本科單子葉植物[5-6,9-10];吸收機制II,即通過一系列的酶促反應合成和分泌麥根酸類物質,與根際的Fe3+形成螯合物,由一類專一的鐵載體(phytosiderophore, PS)吸收途徑被根系吸收利用,多見于禾本科植物[5-6,9-11]。
特別地,鐵調節轉運蛋白IRT在機制I植物根系轉運Fe2+的過程中發揮重要作用,并能調控Fe3+、Mn2+、Cd2+和Zn2+等金屬離子的轉運,有關植物IRT的生物學功能研究主要集中在擬南芥、水稻等模式植物中[12-17]。Eide等[14]最早從擬南芥(Arabidopsis thaliana)中克隆了在根部外皮層中高量表達的AtIRT1,該基因敲除后,擬南芥植株的生長嚴重受阻,并在缺鐵土壤中很快死亡[15]。酵母異源表達試驗表明AtIRT1具有廣泛的底物特異性,可高效轉運Fe2+、Mn2+、Cd2+和Zn2+離子[14,16]。擬南芥AtIRT2在根表皮中高量表達并受缺鐵脅迫誘導而顯著增加,AtIRT2定位于細胞間囊泡中,通過區室化儲存Fe2+,以防止由AtIRT1吸收過多的Fe2+而造成毒害[17]。水稻(Oryza sativa)是特殊的機制II植物,不能將Fe3+還原成Fe2+,但能直接吸收利用Fe2+。水稻OsIRT1和OsIRT2均定位在根表皮細胞質膜,具有類似機制I植物IRT轉運Fe2+和Cd2+的功能[11-12]。因此,水稻根據其吸收鐵的方式同時存在吸收機制I和吸收機制II[5-6,9-12]。
近年來,IRT1同源蛋白陸續在番茄(Solanum lycopersicum)[18]、小金海棠(Malus xiaojinensis)[19]、蘿卜(Raphanus sativus)[20]、杜梨(Pyrus betulaefolia)[21]等園藝植物中被鑒定,但其生物學功能尚不清楚。
葡萄(Vitis vinifera L.)基因組序列已公布[22],然而,葡萄IRT的生物學功能依然未知。釀酒葡萄(V. vinifera L.)是一種全球重要的主要用于釀造葡萄酒的果樹,本研究從釀酒葡萄‘馬瑟蘭’(V. vinifera cv. Marselan)中克隆并鑒定VvIRT1,明確其在釀酒葡萄不同組織部位的特異性表達模式及其對缺鐵和高鐵毒害的響應,為研究果樹鐵素吸收與高效利用機制提供理論依據。
1 材料與方法
1.1 材料
供試材料為山東省煙臺市農業科學研究院葡萄資源圃中的5年生‘馬瑟蘭’成年樹和組培幼苗[23],酵母菌(Saccharomyces cerevisiae)fet3fet4和表達載體pYH23為魯東大學農林工程研究院實驗室保存。
1.2 方法
1.2.1 脅迫處理 利用‘馬瑟蘭’組培幼苗,以1/2 MS液體培養基為對照,參照張璐等[23]、Song等[24]和崔吉潔等[25]方法進行缺鐵(0 mmol/L)和高鐵毒害(500 μmol/L FeNa-EDTA)處理,3次生物學重復,每次處理12棵幼苗。
1.2.2 VvIRT1的克隆 以擬南芥AtIRT1(AT4G-19690)的氨基酸序列為參考序列[14,16],在葡萄基因組數據庫[22]中檢索可能的VvIRT1同源蛋白。將其結果經Pfam(http://pfam.xfam.org/search)在線服務器預測功能結構域。
從葡萄基因組數據庫獲得VvIRT1的CDS序列(coding sequence),設計上下游引物,提取‘馬瑟蘭’組培幼苗整株的總RNA,通過PrimeScriptTMRT reagent Kit反轉錄試劑盒(TaKaRa,大連,中國)合成第一鏈cDNA為模板,利用Prime STARTMHS DNA聚合酶(TaKaRa,大連,中國)擴增目的基因,并送生工生物工程(上海)股份有限公司進行測序驗證。
1.2.3 系統發育樹建立 利用ClustalX 2.0.13軟件對釀酒葡萄、小金海棠、落花生(Arachis hypgaea)、番茄、擬南芥、水稻、杜梨、柑橘(Citrus sinensis)、桃(Prunus persica)、梅(Prunus mume)、白楊(Populus trichocarpa)、蘿卜、玉米(Zea mays)、馬鈴薯(Solanum tuberosum)和陸地棉(Gossypium hirsutum)15種植物的IRT同源蛋白進行氨基酸序列比對,利用MEGA 7.0中的最大相似法(maximum likelihood)構建系統進化樹,分析遺傳進化關系。
1.2.4 實時熒光定量PCR分析 利用NCBI/Primer-BLAST在線服務器,設計VvIRT1的特異性表達引物(上游引物:5'-GTGTAGGACTACTGAACGCC-3',下游引物:5'-CAGACATGCCACCAGCACCC-3'),以葡萄Ubiquitin(GenBank:MH114011)為內參[23,26-29],運用ABI 7500實時熒光定量PCR儀檢測VvIRT1的組織特異性表達特征及其對不同鐵素供應水平的響應差異。反應程序為95℃ 30 s;95℃ 5 s,58℃ 34 s,40個循環;72℃ 10 s。3次生物學重復,采用2-ΔΔCt法計算相對表達量[23-29]。
1.2.5 酵母異源表達 設計構建重組表達載體引物,上游引物序列:5'-GACGGATCCATGGCAACTTCACC TCTCAAA-3'(下劃線為BamH I限制性酶切位點),下游引物序列:5'-GAGTCTAGATCAAGCCCATTTT GCCATTACA-3'(下劃線為Xba I限制性酶切位點),擴增VvIRT1的CDS目的片段,將其與植物表達載體pYH23(Biovector,北京,中國)連接[12,14,16],獲得重組質粒pYH23-IRT1,并進行雙酶切驗證。
酵母突變菌株DEY1453(MATα/MATα ade2/+canl/canl his3/his3 leu2/leu2 trpl/trpl ura3/ura3 fet3-2::HIS3/fet3-2::HIS3fet4-1::LEU2/fet4-1::LEU2)由于突變了fet3fet4位點,在缺失Fe2+的環境下不能正常生長[12,14-15]。根據Eide等[14]和Nakanishi等[12]的方法,將轉化pYH23和重組質粒pYH23-IRT1的酵母細胞分別于液體YPD培養基(1% yeast extract、2% peptone、2% glucose和10 μmol/L Fe2SO4,pH 4.0)培養至OD600=1.0,經10倍梯度稀釋成10-1、10-2和10-3濃度,分別取4個濃度梯度的菌液5 μL點樣于缺失Fe2+的SD固體培養基(1% yeast nitrogen without amino acied和50 μmol/L bathophenanthroline disulfonic acid(BPDS),pH 5.5)上,30℃黑暗培養60 h,觀察菌落生長情況并拍照。
1.2.6 數據分析 利用SPSS 13.0(SPSS Chicago,Ilinois,美國)對數據進行顯著性分析,釀酒葡萄幼苗在脅迫處理與對照條件2個獨立樣品間進行t-檢驗(** P<0.01)。
2 結果
2.1 VvIRT1的鑒定與克隆
以擬南芥AtIRT1的氨基酸序列為參考,在葡萄基因組數據庫中檢索到1個氨基酸一致性為78.5%的IRT同源蛋白,Pfam預測其含有Zinc/iron transporter(PF02535)功能結構域,編碼典型的鐵調節轉運蛋白。依據葡萄基因組數據庫獲得VvIRT1的CDS序列,設計上下游引物進行擴增,獲得一條約1000 bp條帶,經測序,長度為1047 bp,命名為葡萄VvIRT1。
2.2 植物IRT同源蛋白系統發育樹的構建
氨基酸序列比對結果表明,釀酒葡萄、小金海棠、落花生、番茄、擬南芥、水稻、杜梨、柑橘、桃、梅、白楊、蘿卜、玉米、馬鈴薯和陸地棉15種植物IRT同源蛋白的序列一致性高達65.27%,并在6處Motif的功能域區段高度一致(圖1),表明該蛋白在不同物種進化過程中的功能較為保守。

圖1 不同植物IRT同源蛋白氨基酸序列的一致性分析Fig. 1 Amino acid sequence identity analysis of IRT homologs from different plant species
系統發育樹構建結果表明,15種不同科、屬植物的IRT同源蛋白之間的遺傳進化距離差異較大。其中,4種薔薇科植物(杜梨、小金海棠、桃和梅)的IRT同源蛋白聚在一支,禾本科水稻和玉米IRT同源蛋白傾向于聚在一支,十字花科擬南芥AtIRT1和AtIRT2分別與茄科番茄SlIRT1和十字花科蘿卜RsIRT1緊密聚在一支,楊柳科白楊PtIRT1和豆科落花生AhIRT1緊緊聚在一支,而葡萄科VvIRT1、蕓香科柑橘CsIRT1和錦葵科陸地棉GhIRT1緊密聚在一支,三者之間的遺傳距離最近(圖2)。

圖2 不同植物IRT同源蛋白系統發育樹Fig. 2 Phylogenetic tree of IRT homologs from different plant species
2.3 VvIRT1的表達模式分析
實時熒光定量PCR分析結果(圖3)表明,VvIRT1在5年生‘馬瑟蘭’新生根和組培幼苗根中特異表達,而成年樹體新生葉片、新生韌皮部、花蕾期花朵、盛開期花朵、幼果和成熟果實及組培幼苗葉片和莖部的表達量極低。
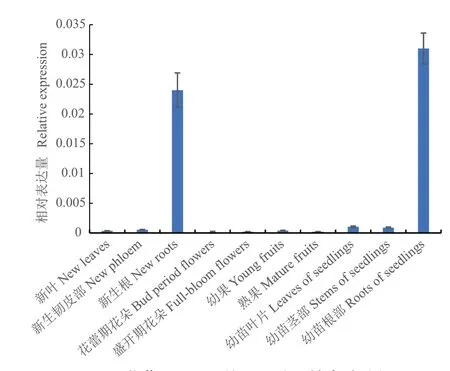
圖3 葡萄VvIRT1的組織特異性表達分析Fig. 3 Tissue-specific expression analysis of VvIRT1 in grape
2.4 VvIRT1對不同鐵素供應的差異響應
以‘馬瑟蘭’組培幼苗為材料,通過進行不同鐵素水平的脅迫,分析VvIRT1的表達模式,結果(圖4)表明,與對照相比,缺鐵處理顯著誘導了VvIRT1在幼苗全部組織(根、莖部和葉片)中的表達,其中,在莖部和葉片中的表達量增加約3倍,而高鐵毒害顯著抑制了VvIRT1在幼苗根中的表達,對莖部和葉片中的表達量沒有影響。
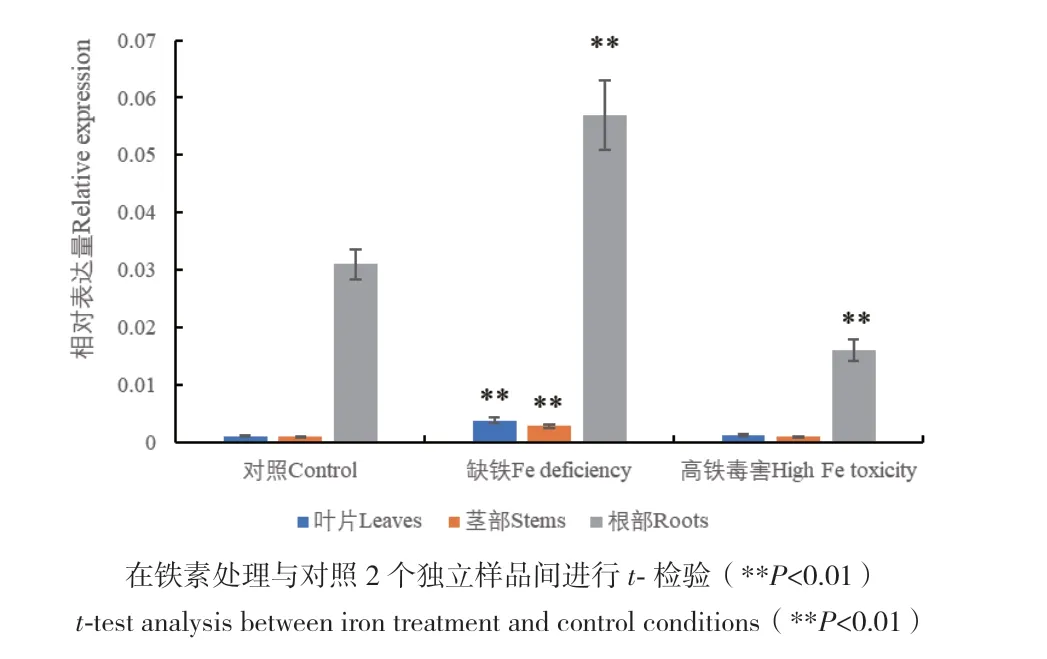
圖4 VvIRT1對不同鐵素供應水平的響應Fig. 4 Responses of VvIRT1 under different Fe supplies
2.5 VvIRT1的功能分析
構建重組表達載體pYH23-IRT1,轉化酵母突變菌株DEY1453,結果(圖5)表明,轉化pYH23空白質粒的DEY1453菌株在缺失Fe2+的SD固體培養基平板上不能正常生長,而轉化pYH23-IRT1重組質粒的DEY1453菌株在缺失Fe2+的SD固體培養基平板上能夠正常生長,表明VvIRT1具有轉運外界Fe2+的功能,進而恢復了DEY1453菌株的生長。

圖5 VvIRT1的酵母功能互補驗證Fig. 5 Verification on the functional complementation of VvIRT1 in yeast
3 討論
植物正常生長所需的鐵濃度為10-9-10-4mol/L,而正常pH土壤中,Fe2+和Fe3+的濃度一般不超過10-15mol/L,遠遠不能滿足植物生長的需求[6,9]。果樹學中,鐵肥施放與果樹生長、花開放、果實品質和產量密切相關,因此,研究果樹鐵素吸收與轉運的分子機制具有重要的科學意義。目前,果樹中鐵吸收與轉運的分子基礎及其調控機制依然未知。葡萄屬于雙子葉植物,其鐵吸收方式屬于機制I[5-6,9-10],但葡萄鐵吸收與轉運的相關分子機制尚未見報道。本研究從釀酒葡萄‘馬瑟蘭’中克隆并鑒定了1個鐵調節轉運蛋白VvIRT1,與已報道植物IRT同源蛋白在氨基酸水平的一致性很高(>64%),暗示VvIRT1可能具有相似的鐵調節轉運蛋白的生物學功能,進一步的酵母功能互補試驗證實VvIRT1具有轉運外界Fe2+的功能,恢復了DEY1453菌株的生長。
本研究所選擇的15種不同科、屬的植物中,VvIRT1與柑橘CsIRT1和陸地棉GhIRT1之間的遺傳距離最近,而同屬于薔薇科的桃、白梨、梅和小金海棠IRT1同源蛋白緊密聚集在一支,禾本科植物水稻和玉米IRT1同源蛋白傾向于聚在一支,而十字花科植物擬南芥和蘿卜IRT同源蛋白緊密聚在一支,這些結果暗示同一科屬或近屬植物IRT同源蛋白在系統發育樹上的遺傳距離可能是更近的,并在長期進化過程中傾向于演化出相似或相近的生物學功能。因此,研究VvIRT1的功能可能為揭示葡萄科植物IRT同源蛋白的生物學功能提供理論支持。
特別地,VvIRT1在成年‘馬瑟蘭’樹體根中和組培幼苗根中特異表達,這一發現與擬南芥AtIRT1和AtIRT2[14-16]、水稻OsIRT1和OsIRT2[11-12]、小金海棠MxIRT1[19]、蘿卜RsIRT1[20]、杜梨PbIRT1[21]、落花生AhIRT1[30]在植物根部高量表達的情況基本一致。已有研究表明,缺鐵脅迫顯著增強了擬南芥AtIRT1[14]、蘿卜RsIRT1[20]、小金海棠MxIRT1[19]、杜梨PbIRT1[21]、落花生AhIRT1[30]在根中的表達水平,與之相一致的是,本研究發現缺鐵處理的確誘導了VvIRT1在幼苗全身不同組織中的表達量,再次暗示植物IRT基因在缺鐵脅迫條件下更傾向于被誘導表達,以便最大限度地發揮轉運Fe2+的活性,進而最大程度地維持釀酒葡萄根部鐵吸收和轉運功能,以保障依賴于鐵的基本生命活動的運行,VvIRT1表達水平的升高可能是釀酒葡萄響應環境缺鐵脅迫的信號之一。此外,本研究發現高鐵毒害顯著抑制了VvIRT1在幼苗根中的表達量,而地上部沒有顯著變化,再次印證VvIRT1表達水平的變化可能是釀酒葡萄適應環境中鐵素供應水平的重要信號之一。
擬南芥AtIRT1具有廣泛的底物運輸特異性,酵母異源表達系統證實,AtIRT1可高效轉運Fe2+、Mn2+、Cd2+和Zn2+離子,AtIRT2不直接參與從土壤中吸收Fe2+,可通過區室化儲存以防止表皮細胞中依賴于AtIRT1吸收過多Fe2+而造成的毒害[14-16]。葡萄與擬南芥同屬于機制I植物,雖然VvIRT1與擬南芥AtIRT1和AtIRT2在系統發育樹上的遺傳距離相對較遠,但其與AtIRT1和AtIRT2在氨基酸水平的一致性分別超過了78%和72%。因此,擬南芥中IRT鐵調節轉運蛋白的生物學功能研究為解析釀酒葡萄IRT同源蛋白提供了技術借鑒。酵母突變菌株DEY1453由于突變了fet3fet4位點,在缺失Fe2+的環境中不能正常生長[12,14,16],本研究借助酵母異源表達系統,將VvIRT1在酵母突變體中異源表達后,直接恢復了酵母突變體的生長情況,初步證實VvIRT1具有轉運Fe2+的能力,具體轉運機制和調控機理值得進一步深入研究。綜上可知,本研究為揭示釀酒葡萄鐵吸收和轉運機制提供了基因資源,并為解析果樹鐵轉調節運蛋白IRT的生物學功能奠定理論依據。
4 結論
從‘馬瑟蘭’中分離并鑒定了1個在根中特異性表達的基因VvIRT1,缺鐵脅迫顯著誘導其在幼苗全身的表達水平,而高鐵毒害顯著抑制了其在幼苗根中的表達水平;VvIRT1具有轉運外界Fe2+的能力,恢復了酵母突變體的生長。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
