星豹蛛成蛛感受器的類型及分布
2023-09-03 13:49:59王美趙萌萌王雅麗趙瑞焦麗亞琳牛越張曉晨李銳
山西農業科學 2023年4期
王美 趙萌萌 王雅麗 趙瑞 焦麗亞琳 牛越 張曉晨 李銳
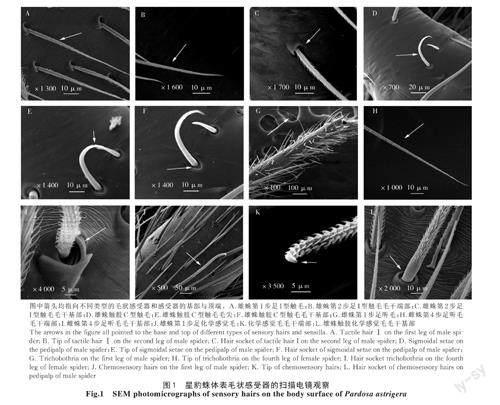
關鍵詞:星豹蛛;感受器;分布規律;形態特征;掃描電鏡
昆蟲會逐漸發展以適應環境的變化[1],與昆蟲同屬于節肢動物門的蜘蛛也會進化發展形成了獨特的行為模式,如捕食、求偶、織網、攜幼等,以及蜘蛛接受種內信息傳遞和蜘蛛與獵物或者天敵之間的種間信息傳遞,這些行為的實現需要感覺系統去識別并感知[2-3]。目前對于蜘蛛體表感受器形態結構和功能的研究較少,最早的研究是1973年FOELIX等[4]通過掃描和透射電子顯微鏡對園蛛科(Araneidae)和狼蛛科(Lycosidae)蜘蛛的觸毛超微結構進行了觀察,為今后進一步研究蜘蛛感受器提供了科研基礎。BARTH[5]通過研究確認,蜘蛛的體表感受器根據形狀的不同分為毛狀感受器(Sensoryhairs)、半球狀感受器(Cellhalf-ball-likemechanosensorysensilla)、裂縫感受器(Slitsensilla)。FOELIX[6]研究發現,蜘蛛的體表分布著成千上萬的體毛;絕大多數體毛是由三合神經支配,進一步證實蜘蛛體毛是重要的感受器。根據體毛接受外界刺激時產生的作用不同,可以將其分為觸毛、聽毛、化學感覺毛、毛狀刺、棘和羽狀毛,其中,聽毛的研究較早且較為深入[7]。劉獻中等[8]采用超薄切片和掃描電鏡對裂縫感受器和半球狀感受器進行了詳細觀察,進一步證實裂縫感受器可以感受其他動物運動以及重力作用產生的刺激,半球狀感受器可以對其他動物運動產生的沖擊起緩沖和保護作用。
星豹蛛(PardosaastrigeraL.Koch)在分類學上屬蜘蛛目(Araneae)狼蛛科(Lycosidae)豹蛛屬(Pardosa)[9-10],是多種農林害蟲的捕食性天敵,具有耐饑餓、壽命長、數量大、成蛛生殖力強、食量大等優點,在長江和黃河流域有廣泛的分布[11-13]。目前關于蜘蛛體表感受器的研究主要集中在機敏異漏斗蛛(Allagelenadifficilis)[14]、園蛛(Argiopebruennichi)[15]和擬環紋豹蛛(Pardosapseudoannulata)[16]。而關于星豹蛛的體表感受器研究僅見JIAO等[17]對其化學感覺毛進行了簡單的描述,其他感受器目前未見相關的研究報道。
本研究利用掃描電子顯微鏡對星豹蛛體表各類型感受器的超微結構、數量及分布進行觀察描述統計,為深入研究星豹蛛種間及種內的信息交流方式提供科研基礎,為開發星豹蛛達到“以蛛治蟲”的生物防治舉措提供充分的理論依據。
1 材料和方法
1.1 試驗材料
供試星豹蛛雌、雄亞成蛛(6齡)均采自于山西農業大學試驗基地(37°25′25″N,112°34′47″E,海拔800m)種植的玉米地和谷子地里,帶回單頭放置在內直徑1.5cm、高8.0cm的指形管中,管底放入濕棉球保濕,置于人工氣候箱中進行飼養,飼養期間溫度保持在(29±1)℃、相對濕度保持在75%±5%、光周期為16L∶8D,以黃粉蟲(Tenebriomolitor)為主要食物,黃粉蟲為市售,每天投喂1次,按時更換指形管保持管內干凈,飼養星豹蛛發育成成蛛供試驗使用。
1.2 試劑與儀器
無水乙醇、叔丁醇,生工生物工程(上海)股份有限公司;2.5%戊二醛、0.1mol/L磷酸鹽緩沖液(pH值7.4),上海源葉生物科技有限公司。
SZX16體視顯微鏡,日本奧林巴斯;MGC-300H人工氣候箱,上海一恒科學儀器有限公司;E-1010型離子濺射儀、JSM-IT300LA型掃描電子顯微鏡,日本JEOL公司;HCP-2臨界點干燥儀,日本HITACHI公司。
1.3 試驗方法
樣品的制備和觀察:取星豹蛛雌、雄成蛛(蛻最后一次皮成為成蟲的第2天)各5頭,首先用70%的乙醇溶液殺死,在體視顯微鏡下將各附肢沿基部切下,用蒸餾水對其進行反復清洗5次;再浸入2.5%的戊二醛溶液中進行固定,4℃冰箱過夜;用0.1mol/L磷酸鹽緩沖液(pH值7.4)清洗星豹蛛標本4次,每次10min;采用梯度脫水法對標本進行脫水,逐級用30%、50%、70%、80%、90%和100%的乙醇溶液浸泡星豹蛛標本,每個濃度梯度保持10min,其中100%的乙醇需要浸泡2次進行脫水;然后用100%的叔丁醇浸泡標本,每次10min,重復3次,充分置換乙醇;采用臨界點干燥法干燥星豹蛛標本;最后通過離子濺射儀噴金,置于掃描電子顯微鏡下觀察星豹蛛體表感受器。
1.4 數據分析
試驗所拍圖片均采用AdobePhotoshopCC2018軟件進行編輯,所得數據運用Excel2019和SPSS22.0軟件進行統計分析,采用Duncan氏新復極差檢驗3種毛狀感受器在體表不同部位分布情況的差異顯著性,同種毛狀感受器在雌、雄間的差異顯著性采用獨立樣本t檢驗分析。
2 結果與分析
2.1 毛狀感受器的類型、形態特征及分布
2.1.1 觸毛形態特征及分布 星豹蛛的毛狀感受器中觸毛根據形狀的不同可以分為2種,分別為I型觸毛(圖1-A)和C型觸毛(圖1-D)。I型觸毛與體表的夾角呈銳角,毛囊呈圓形,一面較凸起,毛干基部與較凸起的那一面毛囊緊貼(圖1-C),觸毛毛干較直,從基部向末端逐漸變細,毛干基部的直徑與末端的直徑差距較大(圖1-B),毛干表面圍繞有許多絨毛狀突起。C型觸毛毛干彎曲呈C型,從基部向末端逐漸變細,毛囊呈橢圓形,毛囊腔窩內壁周圍有明顯的褶皺(圖1-F),C型觸毛毛干較光滑,僅毛干末端有短小的刺狀突起(圖1-E)。
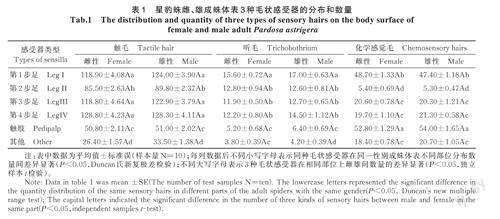
星豹蛛觸毛在雌(F5,54=150.315,P=0.000)、雄(F5,54=173.830,P=0.000)體表的分布均具有顯著性差異,其中,觸毛在第1步足、第3步足和第4步足上分布沒有顯著差異,均顯著高于第2步足、觸肢和其他部位(P<0.05),第2步足顯著高于觸肢和其他部位(P<0.05),其他部位分布數量最少。觸毛在雌、雄成蛛個體之間的數量分布無顯著差異(表1)。
2.1.2 聽毛形態特征及分布 星豹蛛的聽毛細長似絲線(圖1-G),長度大于觸毛和化學感覺毛,聽毛毛干與體表基本呈90°著生,毛干表面著生有大量絨毛,毛干直徑從基部到末端的變化不明顯,與觸毛毛干直徑的變化程度相比較小(圖1-H),毛囊呈橢圓形,毛囊周圍隆起,一側的隆起較高于另一側,隆起的部位較光滑,毛囊窩具有一定深度,聽毛基部從毛囊窩深處伸出與毛囊較隆起的一側間隙較大,毛囊腔窩內壁周圍有多圈明顯的環狀褶皺(圖1-I)。
星豹蛛聽毛在雌(F5,54=44.882,P=0.000)、雄(F5,54=43.334,P=0.000)體表的分布均具有顯著性差異,第1步足分布數量最多,顯著高于第2步足、第3步足、第4步足、觸肢和其他部位(P<0.05),第2步足、第3步足和第4步足之間分布的聽毛數量沒有顯著差異,但均顯著高于觸肢和其他部位(P<0.05),觸肢和其他部位上聽毛數量較少。雌成蛛觸肢和其他部位聽毛分布數量沒有顯著差異,雄成蛛的觸肢上聽毛數量顯著高于其他部位(P<0.05)。聽毛在雌、雄成蛛個體之間的數量分布無顯著差異(表1)。
2.1.3 化學感覺毛形態特征及分布 星豹蛛的化學感覺毛較粗,毛干末端略微彎曲,整個毛干形如弧度較小的S曲線,毛干表面有較濃密的絨毛,但毛干基部光滑無絨毛,化學感覺毛毛干基部與末端的粗細無顯著差別,其直徑小于觸毛、大于聽毛(圖1-J),毛干末端略膨大,形如彎鉤,沒有絨毛著生,在電鏡下可以隱約看見尖端有小的開口(圖1-K)。毛囊呈圓形,化學感覺毛從毛囊的腔窩中伸出,毛囊凸出于體表較光滑,毛囊腔窩內壁有明顯的環狀紋絡(圖1-L)。
星豹蛛化學感覺毛在雌(F5,54=336.979,P=0.000)、雄(F5,54=286.255,P=0.000)體表的分布均具有顯著性差異(P<0.05),在觸肢上分布數量明顯最多,顯著高于步足和其他部位(P<0.05),第1步足上化學感覺毛分布數量顯著高于第2步足、第3步足、第4步足和其他部位;第3步足、第4步足和其他部位分布的化學感覺毛數量沒有顯著差異,均顯著高于第2步足,第2步足上化學感覺毛分布最少。化學感覺毛在雌、雄成蛛個體之間的數量分布無顯著差異(表1)。
2.1.4 其他毛狀體形態特征及分布
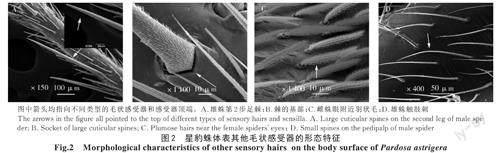
2.1.4.1 棘 星豹蛛的棘長且粗大,零星分布于步足上。棘與體表所成夾角為銳角,毛干從基部向末端逐漸變細,變化程度較明顯,毛干上有許多斜刺狀的絨毛,尖端較光滑沒有絨毛(圖2-A)。棘的毛囊在體表隆起,毛囊呈橢圓形,毛干與體表呈銳角的這一側毛囊表面有短小密集的紋路,另一側毛囊表面較光滑(圖2-B)。
2.1.4.2 羽狀毛 星豹蛛的眼球周圍環繞著生一些長度較短的毛狀體,稱其為羽狀毛。羽狀毛成群分布,毛囊呈圓形,周圍光滑且凸于體表,毛干于毛囊中伸出,與毛囊周圍存在明顯的間隙。毛干與體表呈銳角,毛干基部與端部的直徑無明顯差別,毛干全部向眼的方向伸長靠攏。毛干四周圍繞著生短小的絨毛,絨毛也同樣向眼的方向伸長(圖2-C)。
2.1.4.3 毛狀刺 星豹蛛體表存在許多短小的且聚集著生的毛狀刺,長度較觸毛更短,毛干與體表呈銳角,形狀如拋物線,從基部向末端逐漸變細,毛干上沒有絨毛著生較光滑,毛囊呈圓形凸于體表(圖2-D)。毛狀刺在星豹蛛體表分布數量非常多,大多聚集排列在一起。
2.2 裂縫感受器形態特征及分布
星豹蛛的裂縫感受器根據同一部位數量和形狀的不同,分為單裂縫感受器和多個裂縫感受器形成的豎琴器。單裂縫器形狀圓形稍扁,形似橢圓形兩眼扣(圖3-A),中間有一條明顯的裂縫,裂縫兩側邊緣有輕微凸起且光滑較厚,裂縫頭尾兩端邊緣較薄,與嘴唇形狀有些相似(圖3-B)。單裂縫感受器零星分布于星豹蛛的體毛中,能夠與毛孔很明顯地區分。單裂縫感受器的數量較少,僅觀察到星豹蛛第2步足上分布2個單裂縫感受器。
豎琴器是由多個裂縫感受器成列聚集在一起形成的,豎琴器表面光滑沒有絨毛,其形狀如五六個褶皺平行排列組成,褶皺的長短不相同,其凹槽深淺也有明顯的差異(圖3-C),通常褶皺中最長的一個褶皺其凹槽深度更深。星豹蛛體表豎琴器的分布較單個裂縫感受器的分布較為廣泛,但是數量沒有很多,都著生在附肢的關節處。
2.3 半球狀感受器形態特征及分布
星豹蛛半球狀感受器著生在表面,呈圓形或橢圓形,略微凸起,表面凹凸不平,但是光滑沒有紋路(圖4),集中分布在星豹蛛眼球的周圍。
3 結論與討論
本研究利用掃描電鏡對星豹蛛體表感受器進行觀察,共觀察到毛狀感受器、裂縫感受器和半球狀感受器3種類型。其中,毛狀感受器包含觸毛、聽毛、化學感覺毛、棘、毛狀刺和羽狀毛,裂縫感受器包含單裂縫感受器和豎琴器。
觸毛是蜘蛛體表存在的毛狀感受器中數量最多的一種感受器[18],星豹蛛具有I型觸毛和C型觸毛2種類型觸毛,肖永紅等[19]對北京幽靈蛛(Pholcusbeijingensis)的觸毛進行觀察,本研究結果與其結果相比,星豹蛛的觸毛毛囊窩更深一些,毛干上的絨毛更長更密集,且北京幽靈蛛體表沒有發現C型觸毛;TALARICO等[20]研究認為,C型觸毛可能是機敏異漏斗蛛的接觸化學感受器,能夠同時感受化學和機械刺激。星豹蛛是游獵型蜘蛛,北京幽靈蛛是結網型蜘蛛,觸毛之間的差異可能是由于星豹蛛需要更加靈敏的機械感受器官去接收外界刺激,幫助其順利完成捕食和求偶等行為[21]。
雌雄星豹蛛成蛛的4對步足上聽毛數量均不少于10根,觸肢上觀察到的聽毛數量僅有5~6根。櫛足蛛(Cupienniussalei)與星豹蛛同樣是一種游獵型蜘蛛,但是其步足上分布的聽毛數量在100根以上,觸肢上也不少于50根,它是目前已知蜘蛛類群中聽毛數量最多的種類[22]。蜘蛛聽毛對于氣流的刺激反應靈敏,能夠對物體位置變化產生的空氣振波和某些遠距離物體產生的聲波迅速作出反應[23]。聽毛作為蜘蛛類群里研究最多的一種感受器,在蜘蛛尋找獵物和躲避敵害的方面起著不可忽視的作用,聽毛的數量會由于蜘蛛類群不同而有很大的差異[24]。趙敬釗等[25]對9種不同科的蜘蛛體表聽毛進行觀察,發現聽毛在不同種類的蜘蛛間數量存在明顯差異,同時發現在同種蜘蛛同一步足的不同節上分布的聽毛數量也存在差異,星豹蛛步足的腿節及觸肢的腿節上均未發現聽毛;此外,星豹蛛雌雄成蛛體表分布的聽毛數量有所差異,推測是由于掃描電鏡的過程中聽毛極易脫落,進而對統計結果有輕微影響。
星豹蛛化學感覺毛在步足及觸肢上均有分布,其中,第1步足及觸肢上分布較多,星豹蛛各附肢的不同部位著生的化學感覺毛數量也有很大的差異,在步足及觸肢的腿節均未觀察到化學感覺毛,且每個步足脛節僅有2~3根化學感覺毛零星分布于體毛間,第1步足和觸肢的跗節均密集排列了幾十根化學感覺毛。蜘蛛種類不同化學感覺毛分布及數量也有差異,肖永紅等[26]對5科32種蜘蛛的味覺毛形態、數量及分布進行了觀察,大部分蜘蛛觸肢上未發現化學感覺毛,僅角紅蟹蛛、鼬形微姬蛛和膨脹微蟹蛛觸肢上有數量較少的化學感覺毛存在,熱帶捕獵蛛觸肢末端密布著許多化學感覺毛。化學感覺毛能夠感知同種或異種個體釋放的化學信息物質。目前對于蜘蛛的性信息素化學結構已經有了成熟的研究[27-28],但是對于化學感受器的研究較淺。FOELIX[6]研究發現,蜘蛛化學感覺毛具有2~3種二級神經元,其樹突成束進入圓形腔內。DUMPERT[29]試驗觀察發現,蜘蛛感受氣味的受體能夠產生電位反應。TICHY等[30]通過電生理試驗,觀察到熱帶捕獵蛛雄蛛觸肢上的化學感覺毛在同種雌蛛的性信息素刺激下會產生電生理反應。M?LLER等[15]通過試驗觀察化學感覺毛尖端孔的超微結構,推測蜘蛛化學感覺毛可能具有嗅覺功能。星豹蛛屬于游獵型蜘蛛,其化學感覺毛數量上較其他類型多,推測是因為星豹蛛在尋找獵物、求偶和躲避天敵捕食時,其化學感覺起到的作用較其他類型的蜘蛛發揮的作用更大[31],星豹蛛化學感覺毛末端在掃描電子顯微鏡下可以隱約看見有一開口,后續會采用高倍數精密電鏡進一步觀察。
星豹蛛步足及觸肢上發現存在單裂縫感受器與豎琴器,豎琴器主要分布于附肢的關節處且僅有1~2個,北京幽靈蛛附肢分布有豎琴器不少于10個,且腿節基部具有成群的裂縫感受器分布,數量上較多于星豹蛛[19]。裂縫感受器在節肢動物中存在較為普遍,目前對于裂縫感受器的研究已經有了較為成熟的成果。WITT等[32]研究發現,將蜘蛛的豎琴器蓋住后蜘蛛感受氣味的靈敏度會有明顯的降低,認為蜘蛛嗅覺與豎琴器有關。H?βL等[33]研究表明,成群分布的裂縫感受器和豎琴器可感知到的載荷范圍是單個裂縫感受器的3.5倍左右。陳海峰等[31]進一步證實,北京幽靈蛛雄成蛛在交配過程中的求偶行為主要依靠震動信號。星豹蛛裂縫感受器數量較少,推測星豹蛛在尋覓獵物時主要依靠的不是震動信號,更多的可能是受到視覺、嗅覺、聽覺和化學感覺等多方面的感受來進行判斷的[21]。
在電鏡下觀察到星豹蛛的半球狀感受器成群分布在眼的周圍,此前,有研究發現,在機敏異漏斗蛛的步足上有類似半球狀感受器[14],但星豹蛛步足上尚未發現類似的半球狀感受器。FOELIX[4]研究確認半球狀感受器是蜘蛛體表的壓力感受器,主要分布在頭部,能夠增加接觸面積進而減輕外部對其的沖擊,避免頭部某些器官因為突然的沖擊力而受到傷害。
此外,星豹蛛體表觀察到的棘、羽狀毛及毛狀刺,目前對于其功能尚未清楚。根據感受器表面形態及存在的部位推測棘、羽狀毛和毛狀刺可能對星豹蛛體表或某些器官起到一定的保護和防水作用[14]。
本研究描述了星豹蛛感受器類型、形態特征和分布數量,判斷了部分感受器的作用功能,但是星豹蛛作為農林生態系統中一種重要的捕食性天敵,具有獨特的行為模式,其各種感受器的實際生理功能及在捕食和求偶過程中的作用等,還需進一步借助觸角電位技術(Electroantennograms)和單細胞記錄技術(Singlecellrecordingtechnique)[34]深入探究。
