晉汾白豬背最長肌肌纖維類型和能量代謝特性分析
2023-09-03 13:49:59安家岐何志強(qiáng)孫迪李嬌趙天枝高鵬飛曹果清郭曉紅蔡春波李步高
山西農(nóng)業(yè)科學(xué) 2023年4期
安家岐 何志強(qiáng) 孫迪 李嬌 趙天枝 高鵬飛 曹果清 郭曉紅 蔡春波 李步高
關(guān)鍵詞:豬;背最長肌;肌纖維類型;能量代謝;UCP3基因
肌纖維是骨骼肌的基本組成單位[1]。豬在妊娠90d時,骨骼肌肌纖維數(shù)量就已固定,出生后肌肉的生長主要是依靠肌纖維增粗和類型轉(zhuǎn)化[2]。根據(jù)肌肉收縮速度與能量代謝的差異,骨骼肌肌纖維可以分為I型、IIa型、IIx型和IIb型,對應(yīng)的標(biāo)記基因分別為肌球蛋白重鏈7(Myosinheavychain7,Myh7)、Myh2、Myh1和Myh4[3]。LI等[4]研究發(fā)現(xiàn),肌纖維能量代謝特性直接影響肌肉品質(zhì),不同類型肌纖維占比是影響肌肉品質(zhì)的關(guān)鍵因素。不同豬種的骨骼肌中4種類型肌纖維組成也有明顯差異,肉質(zhì)品質(zhì)差異也較大。劉凡等[5]研究發(fā)現(xiàn),與瘦肉型商品豬相比,我國地方豬種藏豬骨骼肌中I型肌纖維含量高,IIb型肌纖維含量低,藏豬肉色鮮紅,肉質(zhì)較好。
解偶聯(lián)蛋白3(UCP3)是一類負(fù)責(zé)質(zhì)子轉(zhuǎn)運的載體蛋白,分布于線粒體內(nèi)膜上,可以降低線粒體內(nèi)膜上的質(zhì)子電化學(xué)梯度,使電子傳遞鏈和ATP合成解偶聯(lián),抑制ATP的合成。UCP3在骨骼肌、心肌和棕色脂肪組織中表達(dá)量較高,可以調(diào)控機(jī)體能量代謝[6]。HOEKS等[7]在大鼠不同代謝類型骨骼肌中研究發(fā)現(xiàn),氧化能力較低的肌肉組織中UCP3基因表達(dá)水平較高;RUSSELL等[8]在耐力訓(xùn)練和未訓(xùn)練小鼠組間比較發(fā)現(xiàn),UCP3蛋白主要分布在IIb型肌纖維中,I型肌纖維中的含量較低。進(jìn)一步的研究發(fā)現(xiàn),UCP3可以增加骨骼肌中脂肪酸的氧化代謝活性,進(jìn)而調(diào)節(jié)機(jī)體的能量代謝水平[9]。SENESE等[10]在骨骼肌特異性敲除UCP3的小鼠體內(nèi)研究發(fā)現(xiàn),脂肪酸的氧化能力顯著下降。RODR?GUEZ等[11]在禁食后的大鼠骨骼肌中發(fā)現(xiàn),UCP3的表達(dá)量顯著升高,脂肪酸的代謝活性也明顯增強(qiáng)。SCHRAUWEN等[12]研究發(fā)現(xiàn),酵解型肌纖維中脂肪酸氧化代謝活性明顯高于氧化性肌纖維,這可能是酵解型肌纖維中UCP3基因表達(dá)量較高的重要原因之一。LIU等[13]研究發(fā)現(xiàn),人的UCP3基因的啟動子區(qū)域-55位點的堿基發(fā)生突變(C/T),可以有效降低肥胖癥和糖尿病的發(fā)病率。近年來,UCP3基因的單核苷酸位點多態(tài)性與家畜的肉質(zhì)品質(zhì)、眼肌面積、系水力和大理石花紋等性狀顯著相關(guān)的研究也相繼被報道[14-16],但對地方豬種骨骼肌中UCP3基因表達(dá)模式的研究還相對較少。晉汾白豬是由馬身豬、二花臉豬、大白豬和長白豬經(jīng)過雜交選育而成,具有產(chǎn)仔多、生長速度快的特點,但對其骨骼肌肌纖維特性和類型組成以及能量代謝方式的研究未曾報道[17]。
本研究以晉汾白豬和大白豬為研究對象,通過檢測肌纖維類型和能量代謝相關(guān)基因的表達(dá)模式,旨在探究晉汾白豬肉質(zhì)品質(zhì)的遺傳分子機(jī)制,為晉汾白豬骨骼肌肌纖維特性和不同類型肌纖維組成以及能量代謝方式提供數(shù)據(jù)支撐。
1 材料和方法
1.1 材料
1.1.1 樣品采集 斷奶晉汾白豬與大白豬去勢公豬各6頭,于相同環(huán)境和營養(yǎng)水平下在山西農(nóng)業(yè)大學(xué)動物實驗中心飼養(yǎng)。6月齡屠宰,采集背最長肌組織樣品,一部分組織保存于-80℃冰箱中,用于提取組織RNA;另一部分修剪為1cm×1cm×1cm的組織塊,4%多聚甲醛固定后制作組織切片。
1.1.2 主要試劑及儀器 cDNA反轉(zhuǎn)錄試劑盒(TransScript?UniAll-in-OneFirst-StrandcDNASynthesisSuperMixforqPCR)、qRT-PCR試劑盒(PerfectStartTMGreenqPCRSuperMix)、Trizol等購于北京全式金生物技術(shù)有限公司;HE染色試劑盒購于北京智杰方遠(yuǎn)科技有限公司;石蠟、4%多聚甲醛、無水乙醇、中性樹膠等購于北京索萊寶科技有限公司;UCP3單克隆抗體購自Abcam公司;β-actin單克隆抗體購自武漢三鷹生物技術(shù)有限公司;熒光二抗購自美國LI-COR公司。
攤片機(jī)(KD-PIII,美國);切片機(jī)(KD-2260,美國);智能生物包埋機(jī)(KD-BMIV,美國);烘片機(jī)(ZRX-DH3,中國);高速冷凍離心機(jī)(C5415R,德國);熒光定量PCR儀(CFX,美國),遠(yuǎn)紅外光掃描系統(tǒng)(LI-COR,美國)。
1.2 試驗方法
1.2.1 切片制備 流水沖洗4%多聚甲醛固定24h后的樣品組織塊,沖洗12h后用75%酒精、85%酒精、95%酒精各2h、100%乙醇1h進(jìn)行梯度脫水,然后于二甲苯溶液中透明化處理1h。將組織浸入60℃蠟罐中浸蠟3h,進(jìn)行包埋。使用切片機(jī)將包埋好的樣品切成5μm石蠟切片,47℃條件下攤在載玻片上,然后在45℃烘箱中烘片6h。
1.2.2 HE染色 分別用二甲苯I、二甲苯II浸泡烘干的切片各2min,后依次置于無水乙醇I、無水乙醇II、95%、85%、75%酒精中進(jìn)行逐級復(fù)水。蘇木精染液浸染3min,并用蒸餾水洗90s;返藍(lán)后用1%的鹽酸溶液分化30s,蒸餾水洗2min,伊紅染液染色3min,蒸餾水洗1min;然后依次用75%、85%、95%酒精、無水乙醇II和無水乙醇I各處理2min,逐級脫水,二甲苯I、二甲苯II中各2min進(jìn)行透明,中性樹膠液封片,在顯微鏡下觀察并拍照。
1.2.3 肌纖維直徑和密度的測量與分析 在每張切片上隨機(jī)選擇8個視野拍照,每個視野中包含100根肌纖維以上。用軟件Image-ProPlus6.0進(jìn)行分析,測量肌纖維的短軸和長軸,計算肌纖維的平均直徑;對視野內(nèi)的肌纖維計數(shù),然后計算出單位視野內(nèi)肌纖維數(shù)量的平均值,并換算成個/mm2。
1.2.4 提取總RNA與合成cDNA 采用試劑盒提取總RNA。加入20μLDEPC水溶解總RNA,4℃靜置20min后測定其濃度和純度。采用TransS?cript?UniAll-in-OneFirst-StrandcDNASynthe?sisSuperMixforqPCR試劑盒,按照說明書將每個RNA樣品反轉(zhuǎn)錄成cDNA。第一鏈cDNA合成和gDNA去除:加入gDNARemover1μL,5×Trans?Script?UniAll-in-OneSuperMixforqPCR4μL,總RNA500ng,加RNase-freewater補齊至20μL,輕輕混勻后50℃孵育5min,85℃加熱反應(yīng)5s,失活TransScript?UniRT/RI和gDNARemover。完成后于4℃存放。
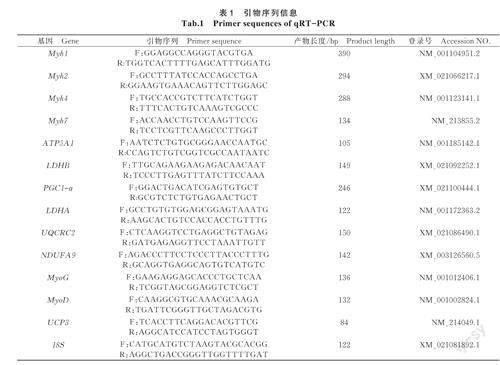
1.2.5 qRT-PCR 用Primer6.0軟件設(shè)計每個基因的特異性引物,內(nèi)參基因為18S,所有引物委托生工生物工程(上海)有限公司合成。引物序列如表1所示。參考實時熒光定量PCR試劑盒說明書,采用20μL體系,包括上下游引物各0.3μL,cDNA4.4μL,2×PerfectStartTMGreenqPCRSuperMix5μL,ddH2O10μL。反應(yīng)程序依照PerfectStartTMGreenqPCRSuperMix說明書進(jìn)行:94℃孵育30s,94℃變性5s,59℃退火反應(yīng)32s,70℃反應(yīng)30s,循環(huán)45次;然后94℃反應(yīng)15s,65℃反應(yīng)30s,95℃反應(yīng)30s。用2-ΔΔCt公式計算各基因相對表達(dá)量。
1.2.6 豬UCP3基因CDS序列的擴(kuò)增 以晉汾白豬背最長肌cDNA為模板,采用PCR技術(shù)擴(kuò)增豬UCP3基因CDS序列。反應(yīng)采用20μL體系,包括上下游引物各1μL,2×EsTaqMasterMix10μL,cDNA2μL,ddH2O6μL。PCR反應(yīng)程序為:95℃孵育5min后95℃變性30s,接著60℃退火反應(yīng)32s,73℃反應(yīng)55s,循環(huán)38次;然后72℃反應(yīng)5min。PCR產(chǎn)物進(jìn)行瓊脂糖凝膠電泳并分析。
1.2.7 蛋白免疫印跡 晉汾白豬背最長肌樣品研磨成粉狀,加入1mLRIPA裂解液,冰上裂解30min,12000r/min離心10min后得到總蛋白,使用BCA蛋白濃度測定試劑盒測定蛋白濃度。蛋白樣品與上樣緩沖液充分混合后100℃變性10min,進(jìn)行凝膠電泳。程序為:80V30min;100V80min。接著進(jìn)行轉(zhuǎn)膜。轉(zhuǎn)膜程序為:100V90min;5%脫脂奶粉于室溫?fù)u床封閉1h;PBS洗滌3次,每次5min;加入適量一抗稀釋液,其中β-actin抗體(1∶2000)和PCNA抗體(anti-Human兔多抗)(1∶1000),4℃過夜;PBS洗滌3次,每次5min;加入二抗稀釋液(1∶20000),室溫孵育1h;PBS洗滌3次,每次5min,然后利用遠(yuǎn)紅外光掃描系統(tǒng)觀察試驗結(jié)果并拍照。
1.3 生物信息學(xué)分析
利用Signal4.0軟件預(yù)測豬UCP3蛋白的信號肽。
1.4 統(tǒng)計分析
試驗采用IBMSPSSStatistics22軟件進(jìn)行單因素方差分析和獨立樣本t檢驗,并分析數(shù)據(jù)差異顯著性。
2 結(jié)果與分析
2.1 豬肌纖維特性分析
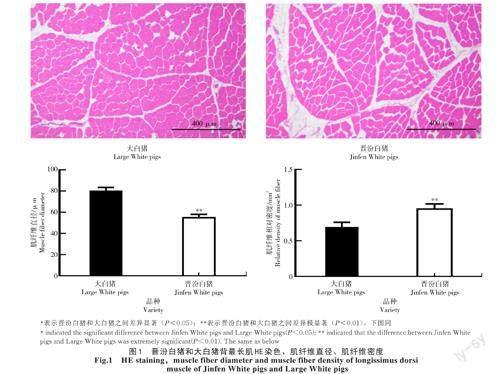
2.1.1 肌纖維形態(tài)分析 HE染色結(jié)果(圖1)顯示,晉汾白豬和大白豬背最長肌細(xì)胞質(zhì)為紅色,染色清晰;細(xì)胞核分布在細(xì)胞邊緣,顯示為藍(lán)色;晉汾白豬骨骼肌肌纖維直徑極顯著低于大白豬(P<0.01),且肌纖維密度極顯著高于大白豬(P<0.01)。
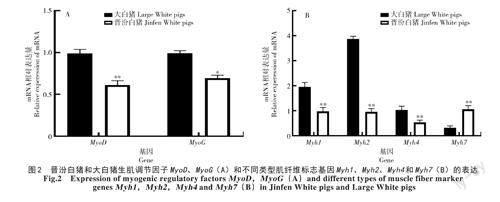
2.1.2 生肌調(diào)節(jié)因子和肌纖維類型標(biāo)志基因表達(dá)量分析 qRT-PCR結(jié)果表明,晉汾白豬中生肌決定因子(MyogenicDeterminingfactor,MyoD)的表達(dá)量極顯著低于大白豬(P<0.01),肌細(xì)胞生成素(Myogenin,MyoG)的表達(dá)量顯著低于大白豬(P<0.05)(圖2-A);晉汾白豬背最長肌中Ⅱ型肌纖維標(biāo)記基因Myh1、Myh2和Myh4的表達(dá)量均極顯著低于大白豬(P<0.01),而Ⅰ型肌纖維標(biāo)記基因Myh7的表達(dá)量極顯著高于大白豬(P<0.01)(圖2-B)。
2.1.3 氧化磷酸化、線粒體和糖酵解活性相關(guān)基因表達(dá)量分析 qRT-PCR結(jié)果顯示,與大白豬相比,晉汾白豬背最長肌中糖酵解代謝標(biāo)志基因乳酸脫氫酶A(LactatedehydrogenaseA,LDHA)和乳酸脫氫酶B(LactatedehydrogenaseB,LDHB)的表達(dá)量極顯著低于大白豬(P<0.01)(圖3-A);氧化磷酸化代謝標(biāo)志基因泛素-細(xì)胞色素c還原酶復(fù)合物核心蛋白2(Ubiquitin-cytochromecreductasecomplexcoreprotein2,UQCRC2)和NADH泛醌氧化還原酶A9亞單位(NADHubiquinoneoxidoreductasecoresubunitA9,NDUFA9)的表達(dá)量極顯著低于大白豬(P<0.01)(圖3-B);且線粒體活性標(biāo)志基因腺嘌呤核苷三磷酸ATP5A1和過氧化物酶體增殖物激活受體γ共同激活劑1α(Peroxisomeproliferatoractivatedreceptor-gammacoactivator1α,PGC1α)的表達(dá)量也極顯著低于大白豬(P<0.01)(圖3-C)。
2.2 晉汾白豬UCP3基因CDS序列擴(kuò)增
利用PCR技術(shù)擴(kuò)增豬UCP3基因的CDS序列,瓊脂糖凝膠電泳結(jié)果顯示,條帶單一且清晰明亮,目的片段長度為927bp(圖4)。分析豬UCP3蛋白與其他物種的序列一致性,按序列一致性從高到低排序依次為狼(93%)、人(90%)、牛(90%)、羊(90%)、大象(90%)、河馬(90%)、大鼠(86%)、小鼠(86%)、雞(74%)、鱖魚(69%)。
2.3 晉汾白豬UCP3蛋白信號肽分析
信號肽預(yù)測顯示(圖5),豬UCP3蛋白Y-Score沒有出現(xiàn)峰值,S-Score為0.186,小于0.5,說明豬UCP3蛋白不存在信號肽。豬UCP3蛋白的D-Score為0.181,小于0.5,表明該蛋白屬于非分泌蛋白。
2.4 豬UCP3的表達(dá)特性分析
2.4.1 晉汾白豬UCP3基因的組織與時序表達(dá)特性分析 qRT-PCR結(jié)果顯示,晉汾白豬UCP3基因在背最長肌和背部皮下脂肪中表達(dá)量較高,其他組織中表達(dá)量較低(圖6-A);而且出生后90d晉汾白豬背最長肌中UCP3基因的表達(dá)量明顯高于出生后1、180d(圖6-B)。
2.4.2 不同豬種UCP3基因表達(dá)分析 qRT-PCR和Westernblot結(jié)果表明,在mRNA水平(圖7-A)和蛋白質(zhì)水平(圖7-C),晉汾白豬背最長肌中UCP3的表達(dá)量均低于大白豬(P<0.05)。
3 結(jié)論與討論
肌纖維是肌肉的基本組成單位,其密度與直徑呈負(fù)相關(guān),且與肉質(zhì)具有顯著的相關(guān)性[18-19]。楊平貴等[20]研究發(fā)現(xiàn),羊的肌纖維直徑越小,肌纖維密度越大,肉質(zhì)也越鮮嫩。姜雪等[21]研究發(fā)現(xiàn),相比于商品豬,長白山野豬骨骼肌中肌纖維的直徑和橫截面積更小,肌纖維密度更大,肉質(zhì)更好。本研究結(jié)果發(fā)現(xiàn),與大白豬相比,晉汾白豬背最長肌中的肌纖維直徑較小,密度較大,表明晉汾白豬肉質(zhì)優(yōu)于大白豬。
MyoD和MyoG是肌肉生成的重要調(diào)節(jié)因子,其憑借特殊的bHLH結(jié)構(gòu)域能夠結(jié)合許多肌肉特異性表達(dá)的基因,從而調(diào)控肌肉的生長發(fā)育。MyoD基因的缺失會導(dǎo)致成肌細(xì)胞無法正常分化,阻礙肌肉正常生長發(fā)育[22];MyoG基因發(fā)生突變或缺失后,小鼠出生后立即死亡,雖然此時小鼠體內(nèi)仍然存在成肌細(xì)胞,但是只有很少的肌纖維,表明MyoG基因在肌肉發(fā)育后期維持肌纖維聚合具有重要作用[23]。HUGHES等[24]研究發(fā)現(xiàn),MyoG基因的過表達(dá)會使得大鼠快肌中氧化酶水平增加,糖酵解酶減少。牛姣艷[25]研究發(fā)現(xiàn),馬身豬背最長肌中MyoD和MyoG在出生后其表達(dá)量先升高后下降,并且其都在出生后90d表達(dá)量最高,而豬在出生后前2個月肌纖維類型會發(fā)生顯著變化,所以,MyoD和MyoG基因在不同類型肌纖維中其表達(dá)有所差異。AGUIAR等[26]對小鼠進(jìn)行短期訓(xùn)練發(fā)現(xiàn),肌纖維直徑增粗與MyoD(r=0.85)、MyoG(r=0.87)之間有顯著正相關(guān)關(guān)系,本試驗結(jié)果與前人研究結(jié)果一致,相較于大白豬,肌纖維直徑較小的晉汾白豬背最長肌中MyoD和MyoG的表達(dá)量均較低,進(jìn)一步表明晉汾白豬和大白豬背最長肌肌纖維類型不同。肌纖維類型與肉質(zhì)品質(zhì)具有顯著相關(guān)性,可以調(diào)控肉色、pH、肌內(nèi)脂肪含量、嫩度等[27]。骨骼肌中肌紅蛋白含量是調(diào)控肉色的重要因子[28-29]。不同類型肌纖維中,肌紅蛋白含量差異較大,I型和IIa型肌纖維中含量最多,IIb型肌纖維中含量相對較低[30]。HU等[31]研究發(fā)現(xiàn),豬骨骼肌中I型和IIa型肌纖維含量越多,肉色越好;IIb型肌纖維所占比例越大,色澤越差。豬骨骼肌中IIb型肌纖維含量與豬肉pH值的下降速度呈顯著正相關(guān);IIb型肌纖維含量較高,肌肉pH值下降速度快,導(dǎo)致肌肉顏色變淺,嚴(yán)重影響肉品質(zhì)[32-33]。HOCQUETTE等[34]研究發(fā)現(xiàn),骨骼肌中I型和IIa型肌纖維含量越多,肌內(nèi)脂肪含量越高,豬肉越鮮嫩多汁,口感較好。本研究發(fā)現(xiàn),晉汾白豬背最長肌中IIb型肌纖維標(biāo)記基因Myh4的表達(dá)量極顯著低于大白豬,I型肌纖維標(biāo)記基因Myh7的表達(dá)量極顯著高于大白豬,表明晉汾白豬的肉質(zhì)優(yōu)于大白豬。
骨骼肌運動所需的ATP主要由氧化磷酸化和糖酵解代謝產(chǎn)生。不同類型肌纖維的氧化磷酸化和糖酵解代謝活性也不相同,I型、IIa型、IIx型和IIb型肌纖維氧化磷酸化代謝活性逐漸降低,糖酵解代謝活性逐漸升高[35]。侯艷茹等[36]研究發(fā)現(xiàn),相比于舍養(yǎng),放牧飼養(yǎng)的羊肌肉中I型和IIa型肌纖維含量較高,IIb型肌纖維含量較低,且氧化磷酸化代謝活性強(qiáng)。GUO等[37]研究發(fā)現(xiàn),相比于6月齡大白豬,馬身豬背最長肌中酵解型肌纖維標(biāo)記基因的表達(dá)量顯著降低,氧化型肌纖維標(biāo)記基因的表達(dá)量顯著升高。李忠秋等[38]研究發(fā)現(xiàn),民豬背最長肌中I型和IIa型肌纖維占比顯著高于大白豬,而IIb型肌纖維含量顯著低于大白豬,LDH活性也顯著低于大白豬。豬骨骼肌中過表達(dá)PGC1α,可以誘導(dǎo)線粒體的生物合成,氧化磷酸化代謝活性明顯增強(qiáng),I型肌纖維含量顯著增加[39]。本研究發(fā)現(xiàn),晉汾白豬背最長肌中氧化磷酸化和糖酵解代謝相關(guān)基因的表達(dá)量都顯著低于大白豬,與以前報道并不一致。晉汾白豬是由二花臉豬、馬身豬、長白豬和大白豬雜交選育的新品豬種,其遺傳變異性較大,這可能是其骨骼肌糖酵解和氧化磷酸化代謝不同于純種豬的主要原因。
UCP3是解偶聯(lián)蛋白家族的成員,在維持機(jī)體產(chǎn)熱和調(diào)控能量代謝方面起著重要作用[40]。骨骼肌中UCP3的表達(dá)量升高,可以促進(jìn)肉毒堿棕櫚酰轉(zhuǎn)移酶1(Carnitinepalmitoyltransferase1,CPT1)的表達(dá),促進(jìn)脂肪酸氧化和脂質(zhì)代謝[41]。UCP3敲除小鼠的體脂沉積顯著增多,而骨骼肌中特異性過表達(dá)UCP3會降低骨骼肌中的脂肪含量[42]。UCP3的表達(dá)特性與肉質(zhì)品質(zhì)也具有顯著的相關(guān)性。胡倩等[43]研究發(fā)現(xiàn),貴州地方糯谷豬和蘿卜豬的UCP3基因外顯子多態(tài)性與肌內(nèi)脂肪含量、嫩度、風(fēng)味等性狀具有顯著的相關(guān)性。任亮[15]研究發(fā)現(xiàn),UCP3基因編碼區(qū)G395A突變與豬胸腰椎間背膘厚和肌內(nèi)脂肪含量顯著相關(guān)。方美英等[44]通過比較中國地方豬和商品豬UCP3基因的序列發(fā)現(xiàn),豬UCP3基因的3′端調(diào)控區(qū)中SNP位點與肌肉和脂肪能量代謝存在相關(guān)性。長白和大白二元雜交豬骨骼肌中UCP3的表達(dá)量較高,ATP產(chǎn)生較少,糖酵解代謝較強(qiáng),肌內(nèi)脂肪含量較低,肌肉嫩度較差[45]。王陽等[46]研究發(fā)現(xiàn),糯谷豬肌細(xì)胞中UCP3的表達(dá)量較低,骨骼肌細(xì)胞中ATP產(chǎn)生效率較高,對脂肪酸供能的依賴性小,肌內(nèi)脂肪含量高,肉品質(zhì)較好。此外,崔悅悅等[47]利用PCR方法成功擴(kuò)增出杜長大UCP3編碼區(qū)序列,得到了927bp的序列信息。本研究同樣利用PCR技術(shù)成功擴(kuò)增出晉汾白豬UCP3基因CDS區(qū)927bp全長序列,并通過蛋白信號肽預(yù)測分析其為非分泌蛋白,與其他學(xué)者研究一致。并且與大白豬比較發(fā)現(xiàn),晉汾白豬中UCP3基因的表達(dá)量較低,表明晉汾白豬背最長肌氧化磷酸化代謝較強(qiáng),肉質(zhì)也相對較好。
相較于大白豬,晉汾白豬背最長肌肌纖維直徑小,密度大,I型肌纖維標(biāo)記基因Myh7的表達(dá)量高,II型肌纖維標(biāo)記基因Myh1、Myh2和Myh4的表達(dá)量低,線粒體活性、氧化磷酸化、糖酵解相關(guān)基因以及UCP3基因的表達(dá)量均較低,表明晉汾白豬骨骼肌中I型肌纖維占比較高,II型肌纖維占比較低,肌纖維氧化磷酸化代謝活性強(qiáng),可能是其肉質(zhì)較好的一個重要原因。
