左聚糖蔗糖酶及其在酶法合成左聚糖中的應用研究
2023-09-11 02:30:28趙紫琰陳倩倩徐海洋徐淋香
中國調味品 2023年9期
趙紫琰,陳倩倩,徐海洋,徐淋香
(1.江蘇省海洋生物資源與環境重點實驗室,江蘇 連云港 222005;2.江蘇海洋大學 海洋食品與生物工程學院,江蘇 連云港 222005;3.江蘇省海洋生物產業技術協同創新中心,江蘇連云港 222005;4.江蘇省海洋資源開發研究院,江蘇 連云港 222005)
Levan果聚糖是一種微生物功能性多糖,其主鏈由β-(2,6)果糖苷鍵連接而成,少量支鏈由β-(2,1)果糖苷鍵連接而成,具有抗氧化、抗炎、抗腫瘤、保濕及益生功能,廣泛應用于食品、化妝品及醫療領域[1]。Levan果聚糖在食品中充當具有益生功能的食品添加劑,在化妝品行業中用作保濕劑,在醫療領域中用作各種大分子藥物的載體[2-4]。
Levan果聚糖可通過化學、微生物發酵以及酶法合成。化學方法合成levan多糖過程復雜,產物難以分離純化且產物種類較單一。微生物發酵法產levan多糖是一種通過產糖菌株在含有高濃度蔗糖的培養基中發酵,經過醇沉、離心、有機溶劑去蛋白等多種工藝處理后得到粗多糖產品的方法,該方法操作較簡便,但仍有許多不足之處,如發酵液容易污染、產物純度及產量較低等。近年來的研究發現,酶法合成在生產levan方面具有更高的效率和產量,因此成為如今最有前景的技術[5]。酶法合成主要是指LSase在適當條件下以高濃度蔗糖溶液為底物,催化形成levan果聚糖的方法。不同溫度、pH、底物濃度、酶液濃度等因素都會影響levan果聚糖的產量和分子量[6]。目前已有的LSase的活性和穩定性無法滿足酶法合成levan的大規模生產,所以尋找活性更強、效率更高的LSase對levan的產業化和商業化至關重要。
1 左聚糖蔗糖酶的分子結構
左聚糖結構式見圖1,左聚糖蔗糖酶分子結構見圖2。
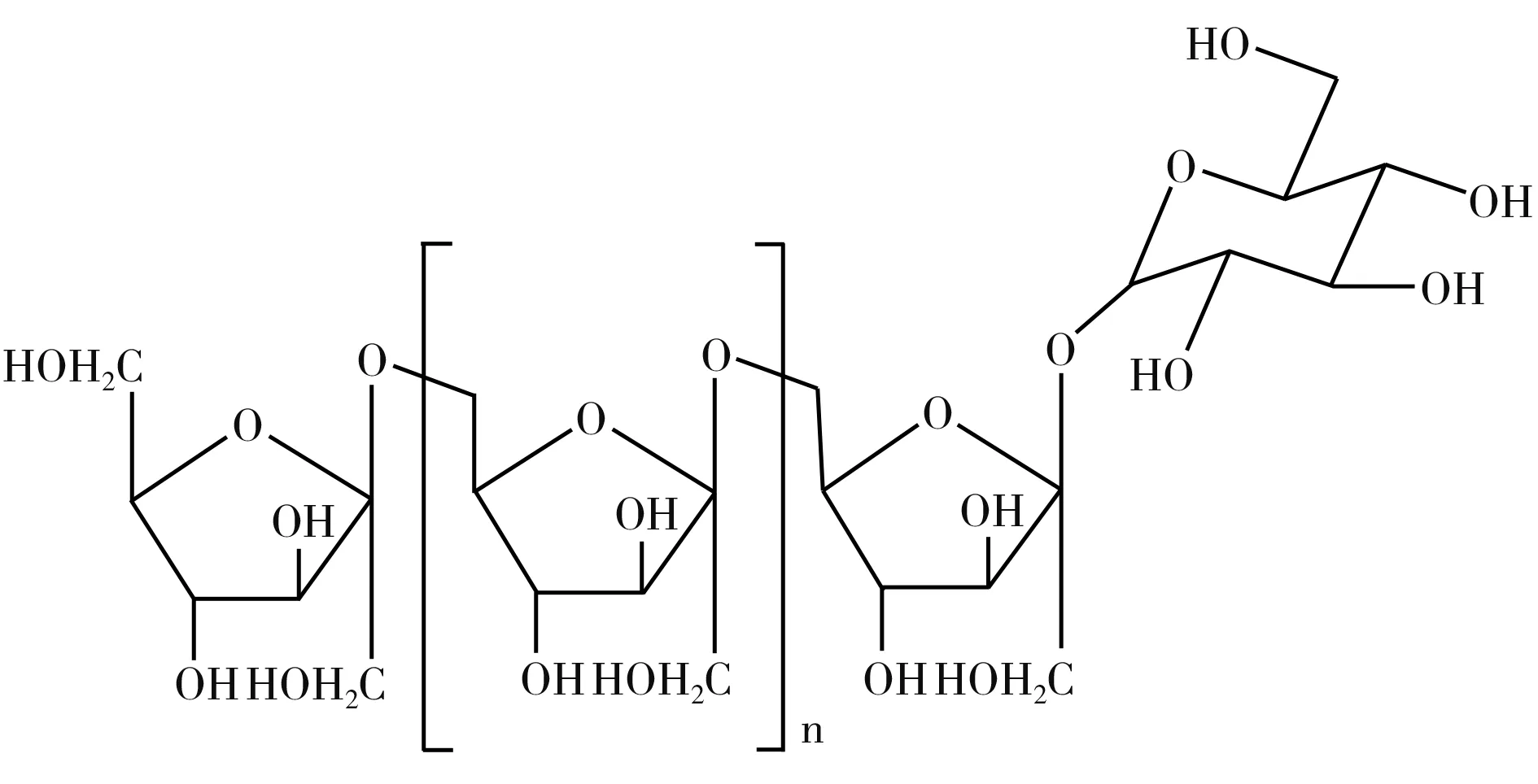
圖1 左聚糖結構式[7]Fig.1 Structural formula of levan
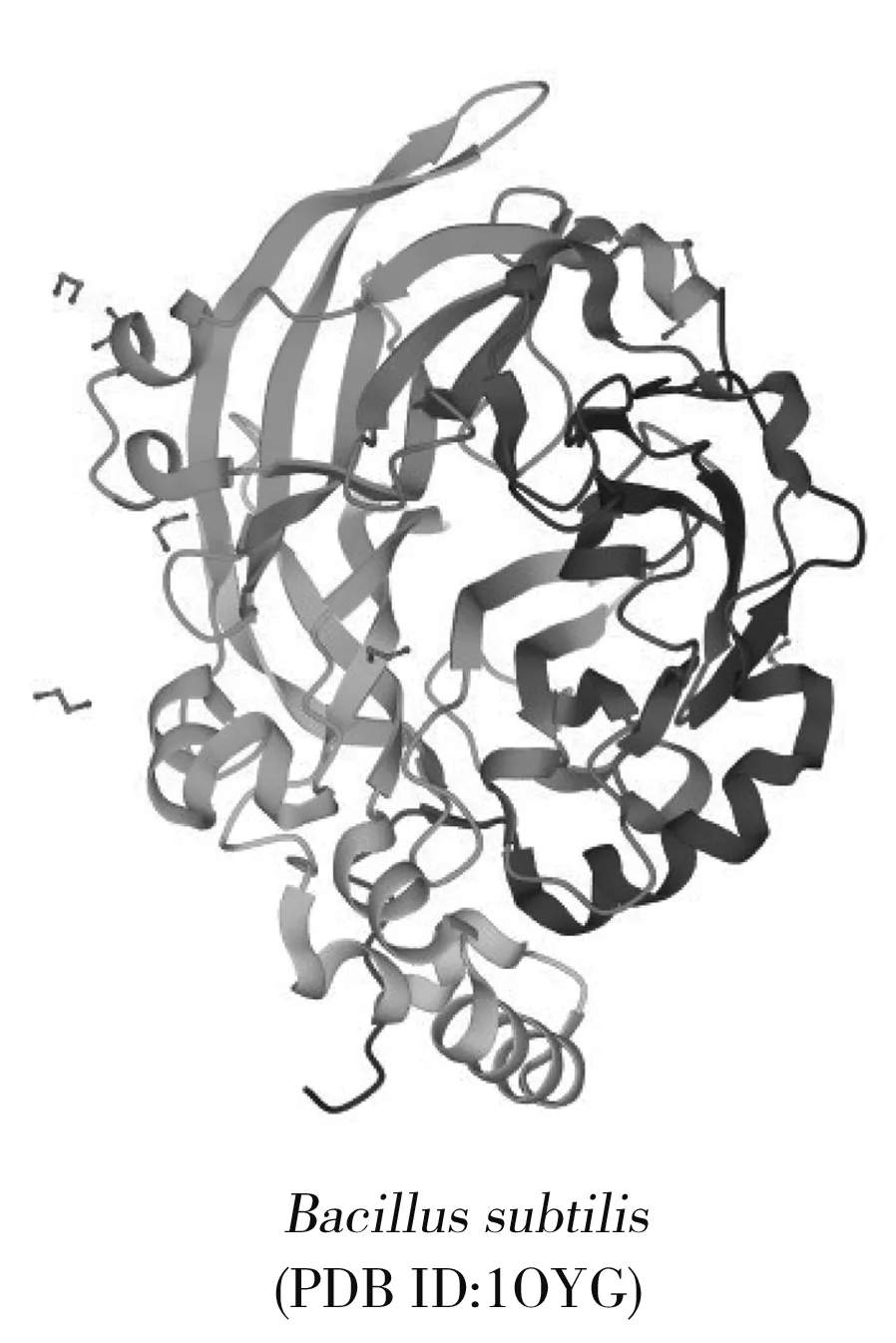
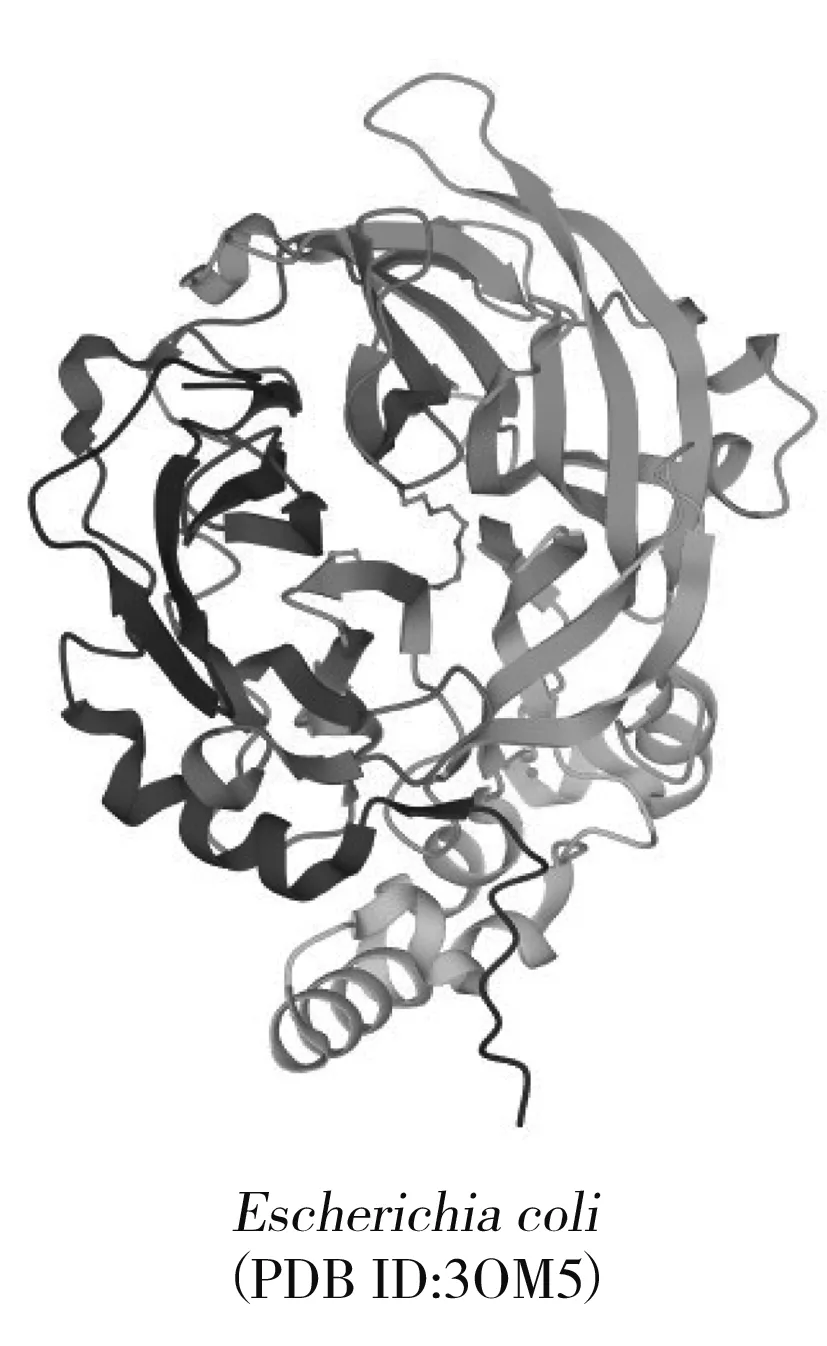
圖2 左聚糖蔗糖酶分子結構Fig.2 Molecular structure of levansucrase
LSase(levansucrase)屬于糖苷酶GH68家族的一類果糖基轉移酶,具有水解和轉移果糖基雙重活性,后者可生成高聚合度的levan果聚糖[5]。
LSase是一種在細胞外起作用的酶,其蛋白質分子結構中通常包括5個重要結構區域:信號肽、N-末端結構域、催化結構域、細胞壁結合區域和C-末端結構域[8]。GH68家族叉頭盒轉錄因子(forkhead box transcription factors,FTFs)都有3個高度保守的氨基酸序列,分別為共價催化基團、廣義酸/堿催化基團和過渡態穩定基團,形成催化三聯體[9]。LSase的晶體結構大多相似,催化結構域均由5片折疊的β-螺旋拓撲結構組成,形成一個帶負電荷、中心凹陷的袋狀結構[10],每個β鏈都由4股扭曲的鏈組成,外層β股幾乎垂直于內股。
2 左聚糖蔗糖酶的反應機制
果糖基轉移酶是一種多功能酶,它能夠直接將蔗糖轉化為高度聚合的果聚糖。蔗糖可同時作為果糖基的供體和受體進行反應,果糖、葡萄糖、低聚果糖及水分子都可作為受體結合由蔗糖產生的果糖基,所以該酶可催化聚合、轉移及水解3種不同的反應,見圖3。該酶的合成產物為菊粉時被命名為菊糖蔗糖酶,聚合產物為levan時被命名為LSase,LSase也可以合成由β-(2,1)糖苷鍵連接的菊粉型低聚果糖(FOS),或形成連接聚合物β-(2,6)鏈的分支點。
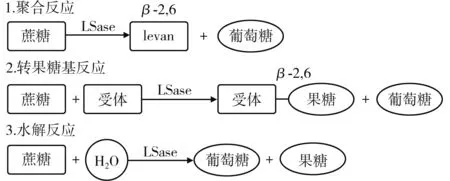
圖3 左聚糖蔗糖酶的3種反應Fig.3 Three reactions of levansucrase
所有的LSase都能夠催化從蔗糖到各種受體分子的果糖殘基轉移,如蔗糖、合成的β-(2,6)糖苷鍵連接的低聚果糖以及分解出的單糖葡萄糖和果糖。根據酶的來源不同,在反應過程中其與受體的親和力有顯著差異,來自革蘭氏陽性菌的LSase產生的低聚糖不會積累中間產物,如枯草芽孢桿菌、巨大芽孢桿菌和唾液鏈球菌[11-13];而來自革蘭氏陰性菌的LSase產生高水平的低聚果糖,導致levan的產量較低,比如運動發酵單胞菌、淀粉芽孢桿菌和丁香假單胞菌等[14-17]。來自革蘭氏陽性菌的β-呋喃果糖苷酶Ms FFase似乎是一個特例,因為該酶即使在高濃度蔗糖的環境下,也主要起水解作用,在蔗糖和乳糖混合物的情況下主要產生乳糖葡萄糖[18]。
雖然所有的LSase都產生高分子量的多糖,但其大小分布會有一定的差異。隨著酶與底物的反應,果聚糖鏈也在不斷增長[19],當原始底物蔗糖耗盡時,合成的聚合物用來充當反應供體。然后,具有轉移酶活性的LSase以外部切割的方式切割levan鏈的β-(2,6)糖苷鍵,導致末端果糖的連續釋放,直到反應在分支點停止[20]。
LSase具有水解和轉移酶活的雙重特性[13],且水解度和轉移率在很大程度上取決于初始反應條件,但在反應過程中會有所不同。蔗糖最適宜的水解溫度為50~60 ℃,在較低溫度(10~40 ℃)下或初始蔗糖濃度較高時水解速率會降低,在蔗糖將近消化完全時,水解會成為主要反應。在反應過程中加入氯化鈉或有機溶劑會促進轉移酶活性[21-23]。LSase的最適pH為5~7,但在大多數情況下,pH變化不影響水解酶活及轉移酶活[24]。來自運動發酵單胞菌的LSase在pH低于6時幾乎只以不溶性微纖維的形式催化合成的高聚合度levan,但在pH高于7時主要以可溶性二聚體的形式催化合成高聚合度levan[25]。
3 左聚糖蔗糖酶的分子改造
3.1 基于產物鏈長調控的分子改造
微生物來源的LSase在以蔗糖為底物合成levan時,具有一定的規律性。即革蘭氏陽性菌來源的LSase天然合成的產物大多是高分子量的levan多糖,而革蘭氏陰性菌來源的LSase合成的產物多為低分子量levan。在不同微生物來源的LSase間存在5個高度保守的氨基酸殘基,分別為“-VWD-”、“-EWSG-”、“-RDP-”、“-DQIER-”和“-TISH-”。經比對發現,兩種來源的LSase在殘基“-VWD-”上存在一定的差異,絕大部分的革蘭氏陽性菌 LSase 在該位置上表現為絲氨酸,而革蘭氏陰性菌LSase在該位置上替換成了丙氨酸。經過序列比對發現,在殘基“-TISH-”上也存在著差異,所有的革蘭氏陽性菌來源的 LSase 在該位置上表現為精氨酸,而革蘭氏陰性菌則表現為組氨酸。
將革蘭氏陰性菌來源的Brsp-LSase中的兩個特殊殘基位點替換成革蘭氏陽性菌來源的LSase中的絲氨酸和精氨酸。結果表明,改變第一個特殊位點的LSase轉糖苷活力和水解活力明顯下降;而在第二個特殊位點改變的LSase在以蔗糖為底物進行反應時不再合成長鏈和高分子量的levan,只能合成蔗果三糖,并且以6-蔗果三糖為主[26]。
3.2 酶的熱穩定性改造
徐煒[27]通過蛋白序列比對、蛋白結構模擬、基因的定點飽和突變等方法,實現了Brsp-LSase的熱穩定性改造。通過與GenBank登錄號的所有LSase相比對,發現在404位氨基酸附近出現了一個“-TEXP-”的氨基酸保守區域(其中X代表可變殘基),且在該氨基酸中除了可變殘基“X”外,其余3個氨基酸殘基均呈現出高度的保守性,于是推斷404位氨基酸殘基對LSase的熱穩定性存在影響。模擬出分子結構后取合適的B-factor(溫度因子或原子位移參數)值較高的區域,經PCR處理得到突變體,之后引入疏水氨基酸,發生了β結構的穩定性增強、活性口袋附近疏水環境的改善以及蛋白結構N端和C端共同形成的“夾具”效應,從而增強了Brsp-LSase的整體結構穩定性。
Yu等[28]針對鏈霉菌Streptomycesdavawensis來源的菊糖果糖基轉移酶進行熱穩定性改造,通過增強酶催化活性口袋附近的分子間相互作用力,最終顯著提升了該酶的熱穩定性,增強了該酶的實用性。
Zhang等[29]發現通過在SacB-T305A中添加TPS-SacB-T305A的額外C端結構域來首次構建融合LSaseTPS-SacB-T305A,該結構域涉及酶的熱穩定性,將融合酶與游離SacB和SacB-T305A相比,融合酶產生的levan分子量分布表現出良好的改善效果。結果表明,在levan聚合時,熔融TPS-SacB-T305A的最佳溫度比游離的SacB-T305A高15 ℃,融合酶在40 ℃下的半衰期增加到4倍以上。
4 左聚糖蔗糖酶催化不同分子量的levan果聚糖合成
反應條件不僅會影響多糖的產率,而且會影響產物levan的平均分子量和支鏈數,雙峰levan的合成是芽孢桿菌LSase的特殊性質[30-32]。影響聚合物分子量的因素主要有酶濃度、溫度、離子濃度和可溶性有機溶劑。低濃度的枯草芽孢桿菌LSase通過促進果聚糖鏈的延長有利于高分子量levan的合成,而初始底物濃度的影響可忽略不計[33-34]。有文獻表明,乙醇、聚乙二醇或乙腈的存在有利于高分子量levan的合成,而離子作用力的增加導致產物從雙峰型levan轉變為單一低分子量levan[35]。在高溫(50 ℃)下,地衣芽孢桿菌RN-01來源的LSase主要合成高分子量levan(612 kDa),但當溫度降至30 ℃或加入氯化鈉時,主要合成低分子量 levan(11 kDa)[36]。
近年來有研究報道,從枯草芽孢桿菌中表達出的SacB-T305A酶與海藻糖-6-磷酸合酶的C末端融合后產生一種新型酶——TPS-SacB-T305A,它可以將產物低分子量levan逆轉為高分子量levan[29]。重組后的酶被證明發生了糖基化反應,有助于緊密維持酶與糖鏈的結合,防止其脫落,以實現糖鏈的延伸。高分子量levan(約2 000 kDa)占總多糖的比例從約4%(由游離SacB-T305A催化)大幅提高到約91%(由TPS-SacB-T305A催化)。
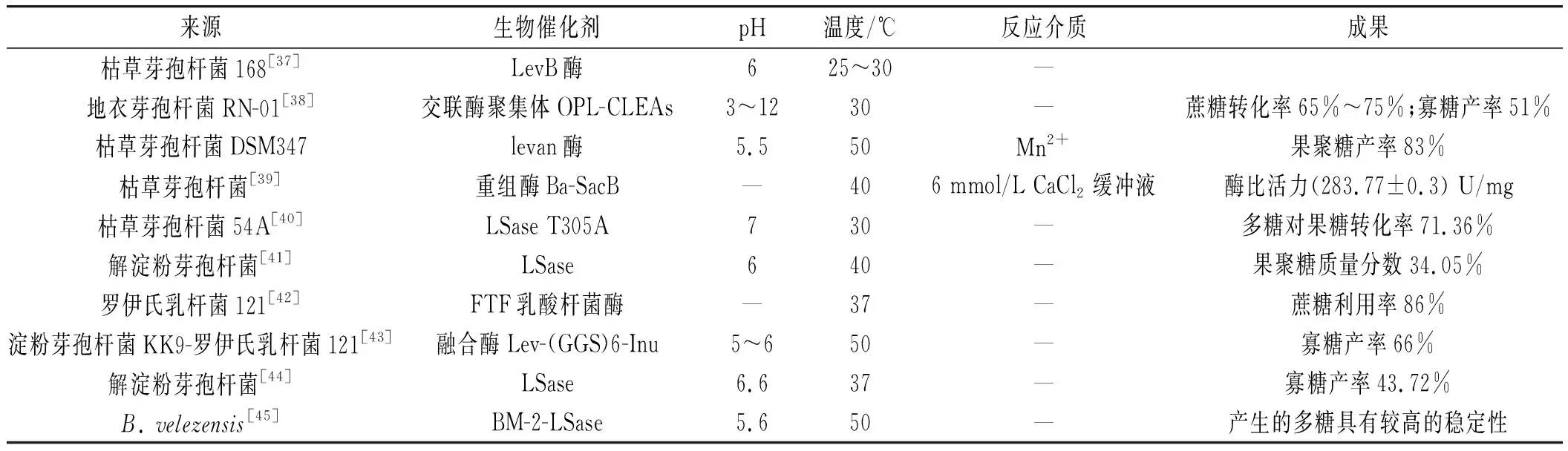
表1 不同微生物來源LSase催化蔗糖產生levan多糖的研究Table 1 Study on the production of levan polysaccharides catalyzed by LSase from different microbial sources
5 左聚糖蔗糖酶的應用
5.1 促進levan果聚糖的生產
周穎[39]運用水熱法制備出磁性納米粒子Fe3O4MNPs,驗證結果表明該物質具有良好的磁吸附性,被EDC-NHS共修飾后可與重組酶Ba-SacB共價結合成固定化酶Ba-SacB@Fe3O4MNPs,該固定化酶具有較好的穩定性,實驗結果表明該酶溫度和pH的穩定性均高于游離酶,并且可以多次重復使用。實驗測得在最適條件下,該固定化酶以蔗糖為底物的反應,蔗糖轉化率為63.42%,levan果聚糖的產率為37.81%。
5.2 生產低聚糖
在GH68家族里存在β-呋喃果糖苷酶,該酶在以蔗糖為底物時能夠專一形成低聚糖[46],因為6-蔗果三糖較珍貴,Abreu等[47]曾定性改造Schwanniomycesoccidentalis來源的β-Ffase合成6-蔗果三糖的能力,以產出更多的6-蔗果三糖。通過鏈長調控的手段控制產物的鏈長,使LSase失去形成長鏈levan的能力,可以有效提高其生產低聚果糖的能力。而對大腸桿菌重組酶Brsp-LSase的第327位基因進行突變后得到突變體H327A,該酶在進行反應時產生以6-蔗果三糖為主的蔗果三糖,其中 6-蔗果三糖的產量為 18.4 g/L,轉化率為9.2%。
5.3 調控不同分子量levan果聚糖的合成
Levan果聚糖的分子量和尺寸分布通常會涉及到蛋白質和反應介質,早于1988年人們就將定點誘變技術應用于LSase的改造[48],通過突變革蘭氏陽性菌來源及革蘭氏陰性菌來源的LSase中的精氨酸或組氨酸,來降低共價中間產物果糖基的形成速率,使水解活性不受影響[49]。Bs-SacB中的殘基K373構成一個結合位點,當被丙氨酸突變時,能夠形成DP>6的低聚果糖;當被精氨酸突變時,其形成的產物DP>8。此外,在Y247位置上必須使用芳香族殘基,以確保可以合成DP>10的產物。
6 總結及展望
微生物來源的左聚糖具有抗氧化、抗炎、抗腫瘤、保濕及益生的作用,LSase是以蔗糖為底物,經過水解、轉移果糖基及聚合3種反應形成不同聚合度的左聚糖。相較于化學合成及微生物發酵合成得到的左聚糖,酶法合成的左聚糖純度及產量更高,目前在最優條件下,LSase的催化反應能從500 g/L蔗糖中提取出(184±3) g/L的levan果聚糖。在此條件下,未來還需解決酶的大量獲取、酶的重復利用及如何高效快速獲得純凈的產物3個問題,如優化產酶條件及提高酶活力來適應市場對于該酶的需求;利用酶的固定化來實現酶的重復使用,提高生產效率,節約成本;目前大多使用乙醇醇沉的方法沉淀多糖,該方法提取率較低,亟待找尋更高效的產物提取純化的方法。盡管目前還存在著許多挑戰,但科學技術的發展必將促進該方法的完善,酶法產左聚糖也會在未來成為一種新的趨勢。
