西北太平洋公海日本鯖生物學特征研究
2023-09-14 11:45:54趙國慶陳俊霖唐峰華陳云云賀建文
海洋漁業(yè) 2023年4期
關鍵詞:質(zhì)量
趙國慶,陳俊霖,2,張 衡,唐峰華,陳云云,賀建文
(1.中國水產(chǎn)科學研究院東海水產(chǎn)研究所,農(nóng)業(yè)農(nóng)村部東海與遠洋漁業(yè)資源開發(fā)與利用重點實驗室,上海 200090;2.大連海洋大學船舶與工程學院,遼寧大連 116000;3.中國水產(chǎn)舟山海洋漁業(yè)有限公司,浙江舟山 316101)
日本鯖(Scomberjaponicus)為大洋暖水性中上層洄游性魚類,廣泛分布于太平洋、印度洋和大西洋大陸架海域,資源量大且食用價值高[1-3],是中國近海和遠洋重要捕撈對象,主要被圍網(wǎng)和拖網(wǎng)所捕獲[4]。數(shù)十年來,隨著捕撈技術的進步、捕撈力量的增加以及人類對水產(chǎn)品需求量的增大,人類對近海漁業(yè)資源的開發(fā)規(guī)模越來越大,漁業(yè)可持續(xù)性水平發(fā)生了顯著變化[5]。研究發(fā)現(xiàn),世界范圍內(nèi),超過90%的近海生物資源被過度開發(fā)[6]。日本鯖作為中國近海的主要捕撈對象之一,自20世紀80年代即表現(xiàn)出漁獲個體小型化、低齡化的特點,90年代即出現(xiàn)過度捕撈現(xiàn)象[1,7],因此,加強對西北太平洋公海日本鯖資源的探索和開發(fā)對保護近海資源有一定的積極作用。
西北太平洋隸屬于聯(lián)合國糧農(nóng)組織(Food and Agriculture Organization,F(xiàn)AO)61區(qū),該區(qū)域具有十分重要的漁業(yè)地位,不僅是FAO統(tǒng)計區(qū)中的高產(chǎn)區(qū)之一,且區(qū)域內(nèi)的國家如中國、日本和韓國等都具有悠久的漁業(yè)捕撈和漁業(yè)消費歷史[8]。近年來,我國逐漸加強對西北太平洋公海漁業(yè)資源的開發(fā)力度,并積極參加國際上對該海域的漁業(yè)管理工作。2012年2月24日,我國作為締約方之一簽署了北太平洋公海漁業(yè)資源養(yǎng)護管理公約(Convention on the Conservation and Management of High Seas Fisheries Resources in the North Pacific Ocean),其后各締約方成立了北太平洋漁業(yè)委員會(North Pacific Fisheries Commission,NPFC),開始對北太平洋漁業(yè)進行科學管理,以期實現(xiàn)該海域漁業(yè)資源的可持續(xù)利用[9]。日本鯖作為西北太平洋重要捕撈品種之一,是NPFC的重點管理對象。據(jù)NPFC統(tǒng)計,該海域日本鯖2021年的產(chǎn)量超過50萬t,日本捕撈產(chǎn)量占比63.5%,而我國僅占20.2%,具有巨大的發(fā)展空間。
目前,我國對于西北太平洋公海日本鯖的研究內(nèi)容涉及漁場變動[10-11]、資源評估[12]、單位捕撈努力量漁獲量標準化[13]、漁獲組成[14]、攝食生態(tài)[15]等方面。生物學特征是進行諸多深入研究的基礎,目前該海域的相關研究仍有待完善和補充[16-17],基于此,本文以2016—2021年西北太平洋公海捕撈的日本鯖樣品為對象,對其生物學特征進行了研究,以期豐富西北太平洋公海日本鯖生物學內(nèi)容,為進一步研究其資源評估、開發(fā)利用與科學管理提供基礎資料。
1 材料與方法
1.1 數(shù)據(jù)來源及漁場區(qū)域
本文所用日本鯖樣品來源于西北太平洋公海生產(chǎn)漁業(yè)企業(yè),現(xiàn)場隨機取樣并冷凍保存,記錄采樣時間及經(jīng)緯度,樣品采集時間為2016—2021年,共4 417尾(表1),作業(yè)區(qū)域分布范圍為35°~43°N、150°~162°E(圖1),作業(yè)方式為燈光圍網(wǎng)捕撈。由于氣候原因及日本鯖的生長發(fā)育情況,我國在西北太平洋公海的主要作業(yè)時間集中于每年4—12月,季節(jié)劃分為春季(4—5月)、夏季(6—8月)、秋季(9—10月)、冬季(11—12月)。
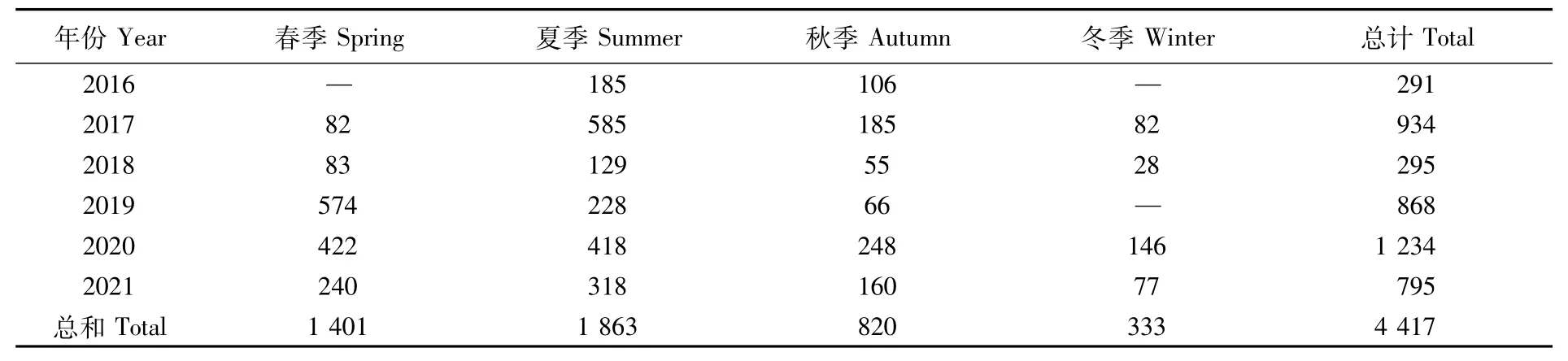
表1 西北太平洋公海日本鯖采樣信息Tab.1 Sampling information of Scomber japonicus in the high seas of the Northwest Pacific(尾)
圖1 西北太平洋公海日本鯖采樣區(qū)域Fig.1 Sampling area of Scomber japonicus in the high seas of the Northwest Pacific
1.2 生物學測定
依據(jù)《海洋調(diào)查規(guī)范》[18]在實驗室對樣品進行生物學測量,測量指標包括性別、性腺成熟度、攝食強度、叉長(FL)和體質(zhì)量(W),叉長與體質(zhì)量分別精確到1 mm和0.1 g。性腺分為6期,分別用Ⅰ~Ⅵ來表示,攝食強度分為6級,用0~5表示,分期和分級標準參照殷名稱[19]。在實驗室進行生物學測量時,對所有樣品的FL和W進行測量,單次樣品不超過30尾時,全部解剖并對其性別、性腺成熟度和攝食強度進行辨認并記錄,超過30尾時,隨機抽取30尾進行解剖,另外,2016年未鑒定樣品性別。
1.3 數(shù)據(jù)處理
1.3.1 叉長、體質(zhì)量分布
利用頻度分布法將體長和體質(zhì)量進行分組并統(tǒng)計分析,體長組距10 mm,體質(zhì)量組距20 g。分別利用K-S檢驗和單因素方差分析(one-way ANOVA)檢測日本鯖雌雄群體叉長分布是否存在統(tǒng)計學差異及顯著性。
1.3.2 叉長-體質(zhì)量關系
利用冪函數(shù)擬合日本鯖叉長-體質(zhì)量關系,其公式表示為:
式中,W為體質(zhì)量(g),F(xiàn)L為叉長(mm);a、b為常數(shù),分別代表生長條件因子和異速生長因子。b值通常在2.5~4.0之間,如果魚類等速生長,則b=3,或接近3;b>3和b<3分別代表正異速生長和負異速生長[20-21]。同時,利用協(xié)方差檢驗性別對叉長體質(zhì)量關系的影響。
1.3.3 性比、性成熟度和初次性成熟叉長
分別統(tǒng)計各年份日本鯖雌雄個體數(shù)量以及性腺成熟度,利用卡方檢驗分析判斷雌雄比例是否符合1∶1。利用多因變量方差檢驗判別叉長和體質(zhì)量與成熟度等級的顯著性關系。一般將種群中50%的個體達到性成熟時的叉長作為該種群初次性成熟叉長,本文即將此時的叉長作為日本鯖的初次性成熟叉長[22-23],計算公式如下:
式中:Pi為日本鯖雌性成熟個體占組內(nèi)樣本比例;Li為各雌性叉長組上限(mm),L50為50%個體初次性成熟叉長(mm),c為模型參數(shù)。
1.3.4 攝食等級和肥滿度
統(tǒng)計分析各年份日本鯖的攝食等級,利用多因變量方差檢驗判別叉長和體質(zhì)量與攝食等級的顯著性關系。采用Fulton狀態(tài)指數(shù)計算日本鯖個體肥滿度及平均肥滿度,肥滿度計算公式如下:
式中:K為肥滿度系數(shù),W為體質(zhì)量(g),F(xiàn)L為叉長(cm)。采用單因素方差分析驗證叉長組與肥滿度的差異顯著性。
本文數(shù)據(jù)分析采用Microsoft Excel 2019和IBM SPSS Statistic 19.0,圖表繪制使用Origin 2018。
2 結果與分析
2.1 叉長、體質(zhì)量組成
日本鯖叉長和體質(zhì)量分布如圖2所示。總體上,4 417尾日本鯖叉長范圍為102~400 mm,分布呈雙峰型,平均叉長236.50 mm,優(yōu)勢叉長組190~230 mm和270~300 mm,占比51.63%;除2019年、2021年和夏季外,其他年份或者季節(jié),其叉長均表現(xiàn)出明顯的雙峰分布。總體上,體質(zhì)量分布呈雙峰型,第二個峰值低于第一個峰值,體質(zhì)量范圍為9.6~898.8 g,平均體質(zhì)量165.0 g,優(yōu)勢體質(zhì)量組50~150 g,占比46.20%;各年份和各季節(jié)體質(zhì)量的分布峰型具有一定差異性,但多數(shù)仍為雙峰型分布。
不同年份和不同季節(jié)的叉長和體質(zhì)量分布表現(xiàn)出一定差異性,體質(zhì)量差異性強于叉長。按年份分析,2020年叉長和體質(zhì)量差異性顯著強于其他年份,2018年叉長和體質(zhì)量均值最高,2016年最低。按季節(jié)分析,夏季的叉長和體質(zhì)量差異性強于其他季節(jié),冬季的均值最高,夏季最低(圖3)。
顯著性分析結果顯示,日本鯖雌雄群體叉長分布不存在統(tǒng)計學差異(P>0.05),而不同年份和不同季節(jié)叉長分布均差異極顯著(P<0.01)。體質(zhì)量分布與年份之間存在極顯著差異(P<0.01),多重比較分析顯示2019年與2020年差異不顯著(P>0.05),其他年份之間均存在顯著性差異(P<0.05)。
2.2 叉長-體質(zhì)量關系
采用冪指數(shù)回歸對日本鯖叉長和體質(zhì)量進行擬合得到曲線,如圖4所示,冪指數(shù)關系式為:
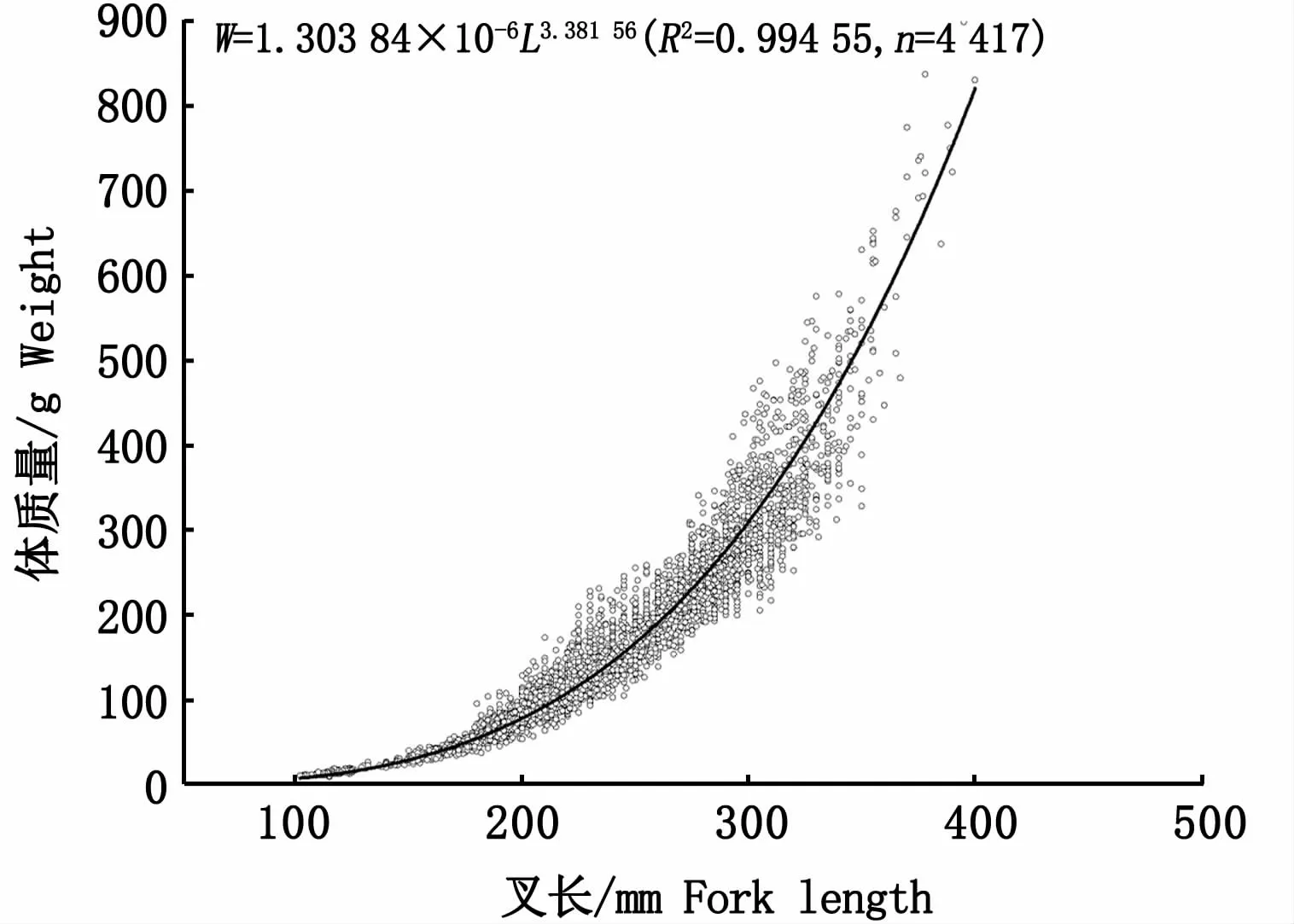
圖4 西北太平洋公海日本鯖叉長-體質(zhì)量關系Fig.4 Relationship between fork length and weight of Scomber japonicus in the high seas of the Northwest Pacific
按不同年份和不同季節(jié)對日本鯖的叉長-體質(zhì)量關系進行分析,方程參數(shù)和擬合曲線分別如表2和圖5所示。按年份分析,a值范圍為7.10×10-7(2016年)~2.56×10-4(2021年),b值范圍為2.44(2021年)~3.49(2016年),2021年變異程度強于其他年份。按季節(jié)分析,a值范圍為1.09×10-6(冬季)~1.17×10-5(夏季),b值范圍為2.99(夏季)~3.42(冬季)。年間和季節(jié)間a值的變化幅度分別為2.55×10-4、1.06×10-5,表明年間日本鯖叉長-體質(zhì)量關系的變化強于季節(jié)。協(xié)方差分析(ANOVA)表明,性別對叉長-體質(zhì)量關系影響不顯著(P>0.05)。
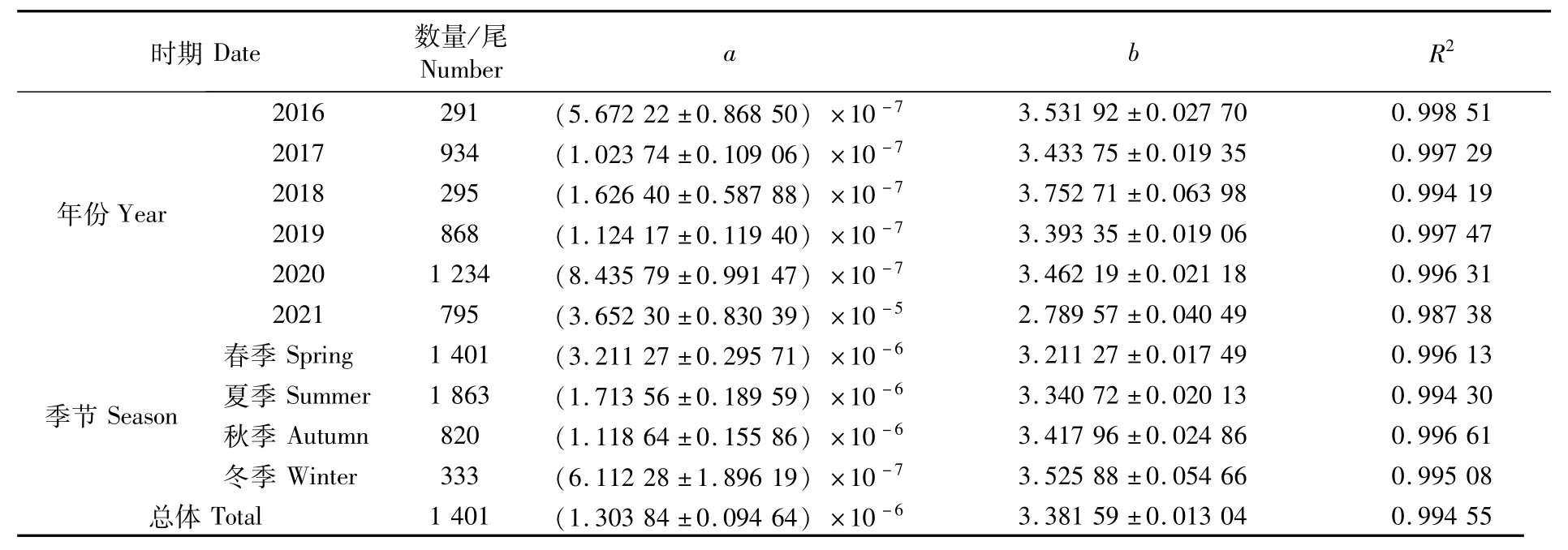
表2 西北太平洋公海日本鯖叉長-體質(zhì)量冪函數(shù)參數(shù)Tab.2 Fork length-weight power function parameters of Scomber japonicus in the high seas of the Northwest Pacific
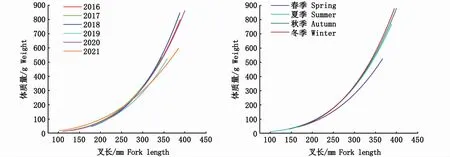
圖5 西北太平洋公海日本鯖各年份和各季節(jié)叉長-體質(zhì)量關系Fig.5 Fork length-weight relationship variations by years,seasons for Scomber japonicus in the high seas of the Northwest Pacific
2.3 性別分析
2.3.1 性比與性成熟度
共鑒定2 317尾日本鯖的性別和性腺成熟度,其中雌性1 472尾,雄性845尾,雌雄比為1.74∶1。卡方檢驗顯示,雌雄比與1∶1差異顯著(P<0.05)。
按照不同年份和不同季節(jié)進行分析,性成熟度均以Ⅱ期占比最高,其次為Ⅲ期,Ⅴ期和Ⅵ期群體均占比較少(圖6);不同年份和不同季節(jié)各叉長組的雌雄比具有一定差異性,大個體中,雌性占比較高,當叉長超過390 mm時,雌性占比達到100%(圖7);隨叉長增大,達到性成熟的個體(Ⅲ和Ⅳ期)也隨之增加,在性成熟個體中,Ⅴ期和Ⅵ期個體占比少(圖8)。多因變量方差分析表明,叉長顯著影響性腺成熟度(P<0.01)。
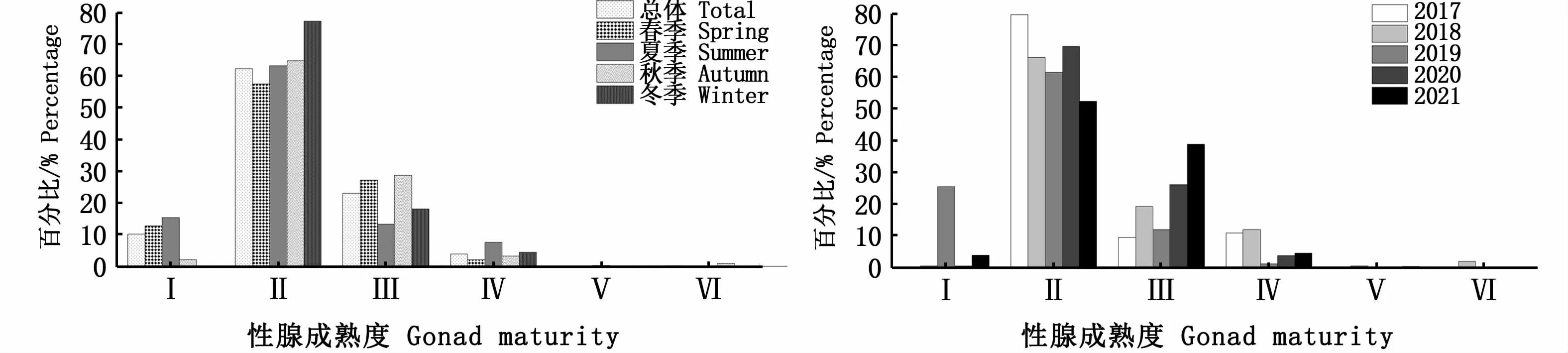
圖6 西北太平洋公海日本鯖各年份和各季節(jié)性腺成熟度Fig.6 Gonad maturity of Scomber japonicus in the high seas of the Northwest Pacific by years and seasons
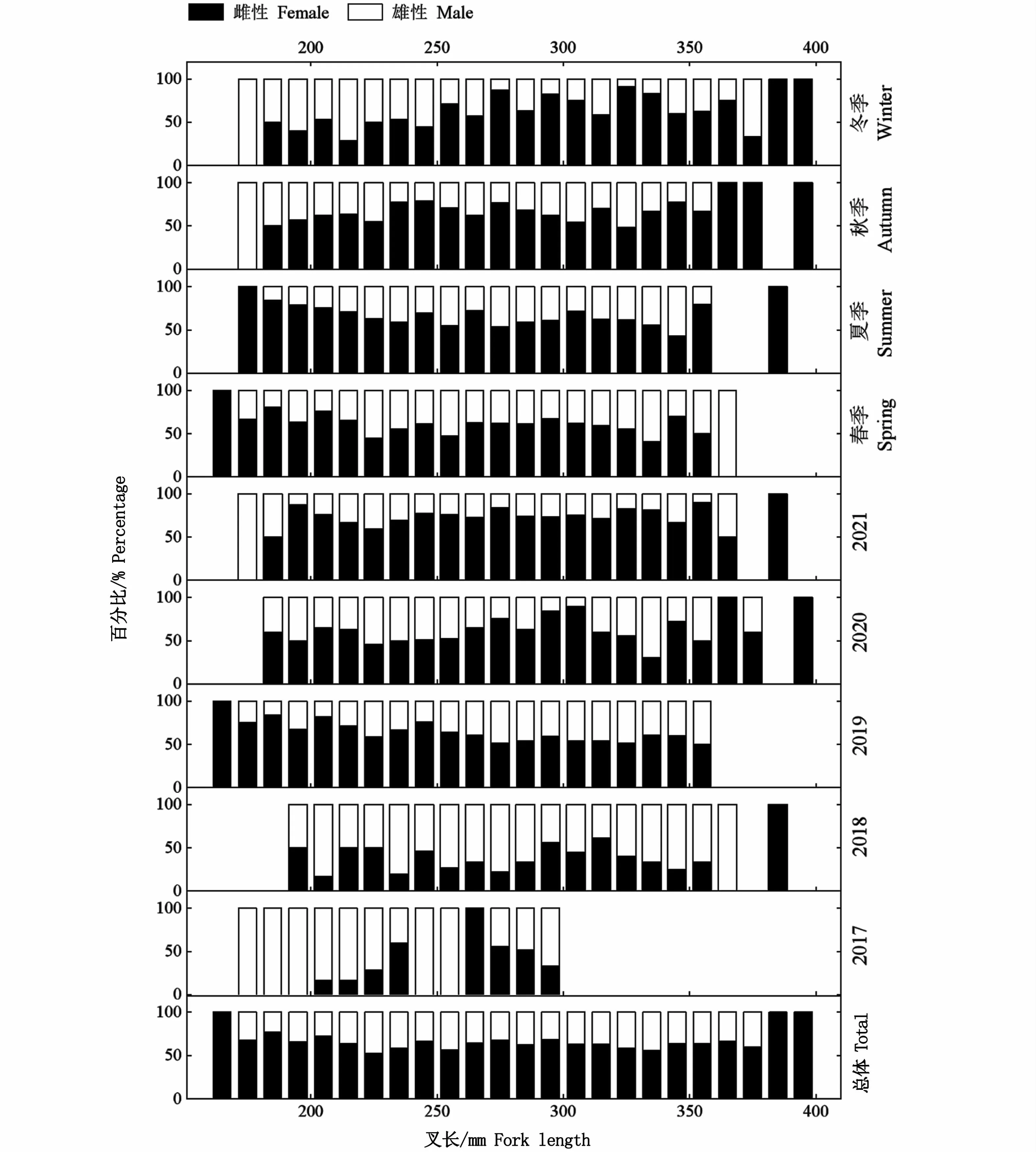
圖7 西北太平洋公海日本鯖各年份和各季節(jié)性比與叉長的關系Fig.7 Relationship between sex ratio and fork length of Scomber japonicus in the high seas of the Northwest Pacific by years and seasons
2.3.2 初次性成熟叉長
根據(jù)不同叉長組中性腺成熟個體的百分比擬合日本鯖邏輯斯蒂曲線(圖9),結果顯示,雌性、雄性個體初次性成熟叉長分別為(298.08±4.32)mm、(287.82±4.26)mm,方程如下:
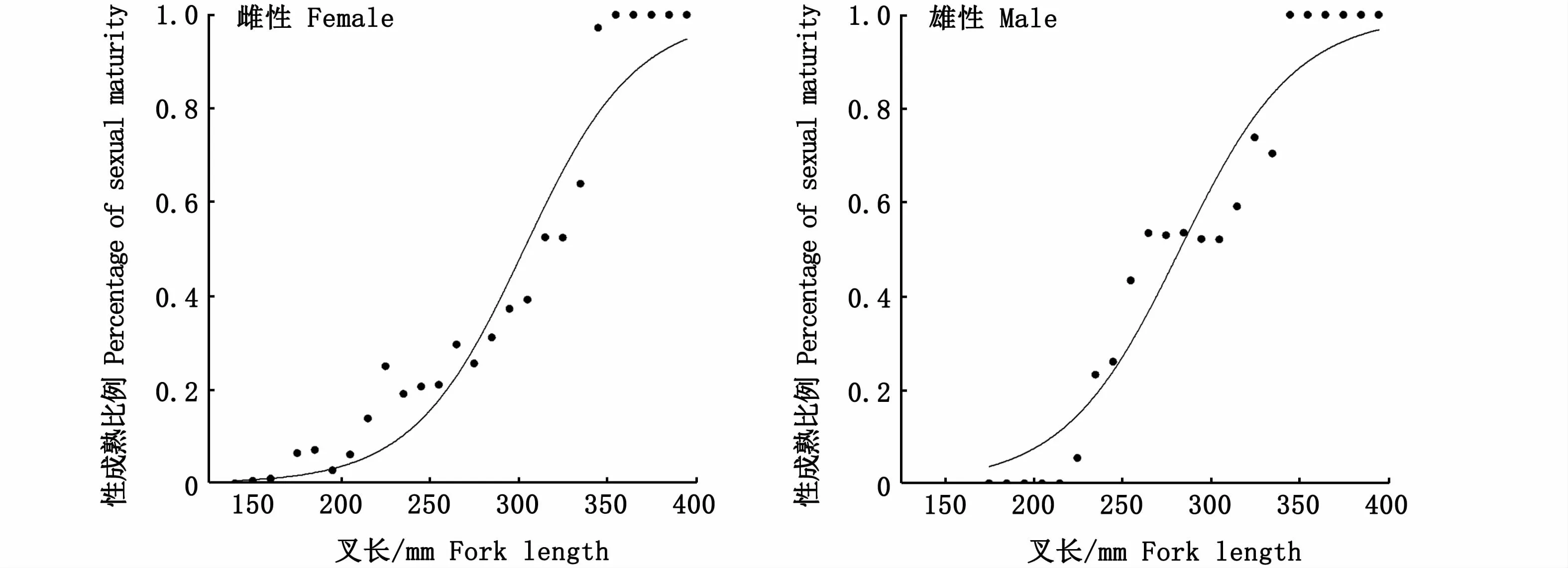
圖9 西北太平洋公海日本鯖初次性成熟叉長Fig.9 Fork length at the first sexualmaturity of Scomber japonicus in the high seas of the Northwest Pacific
2.4 攝食等級和肥滿度
2.4.1 攝食等級
總體上,攝食等級以1~3級為主,占比59.05%,5級占比最少,為0.52%;按不同季節(jié)分析,3~5級各自的占比隨春夏秋冬基本上逐漸增加,年間變化較為隨機,5級個體僅在2021年出現(xiàn)(圖10)。各叉長組所對應的攝食強度組成如圖11所示,空胃以及5級占比均較少,空胃在小于140 mm的小個體中占比高,2級占比最高,為33.55%。攝食強度與叉長顯著相關(P<0.05),與性別的相關性關系不顯著(P>0.05)。
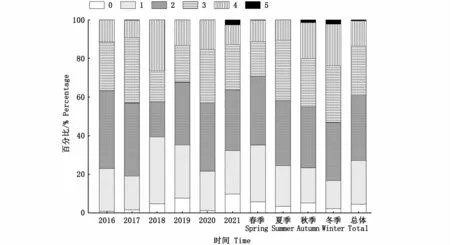
圖10 西北太平洋公海日本鯖各年份和各季節(jié)攝食等級組成Fig.10 Feeding intensity of Scomber japonicus in the high seas of the Northwest Pacific by years and seasons

圖11 西北太平洋公海日本鯖各年份和各季節(jié)不同叉長組與攝食強度百分比組成Fig.11 Percentage of feeding intensity in different fork length groups of Scomber japonicus in the high seas of the Northwest Pacific by years and seasons
2.4.2 肥滿度
總體上,對所有個體分析結果顯示,K值范圍為0.64~2.45,均值1.30。年份和季節(jié)均顯著影響其肥滿度(P<0.05),年間K值變化范圍為0.97(2019年)~1.14(2021年),季節(jié)間按照春夏秋冬依次增大,范圍為0.97~1.19。叉長顯著影響肥滿度(P<0.05),隨著叉長增加,日本鯖肥滿度基本上均隨之增大(圖12)。
情況 3.2.1 C1中的集合都是Y中頂點色集合,4,5,6中至少有2種色同時包含在每個C(ui)中,不妨設4,C(ui), i=1,2,…,10,則C2中的集合都不是X中頂點色集合,且至多有3個不是Y中頂點色集合。
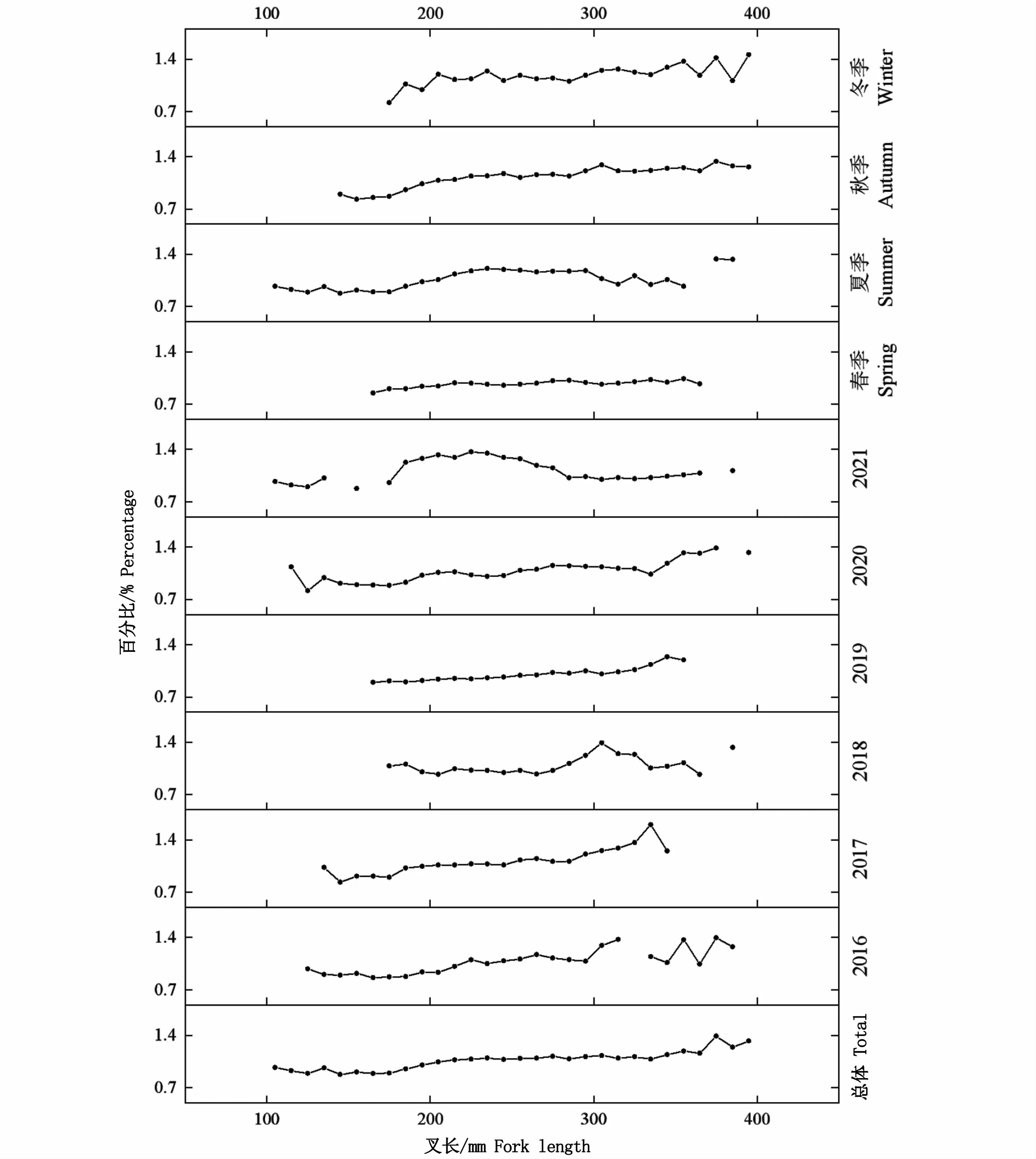
圖12 西北太平洋公海日本鯖肥滿度與叉長的關系Fig.12 Relationship between fork length and fatness index of Scomber japonicus in the high seas of the Northwest Pacific
3 討論
3.1 叉長-體質(zhì)量關系
體長和體質(zhì)量是魚類最基本的生物學特征,通常被換算為資源評估模型的基本參數(shù)用于漁業(yè)科學研究,也是魚類個體低齡化、小型化最直觀的表征[24],兩者之間的函數(shù)關系一直被用于描述魚類的生長特征。其中,冪函數(shù)公式W=aLb是用來描述魚類生長最常用的關系式,式中a和b通過統(tǒng)計分析得出,分別代表條件因子和異速生長因子,是漁業(yè)資源評估的重要參數(shù)[25],能夠反映魚類生長的穩(wěn)定性和適應性[26]。參數(shù)a代表種群生長的環(huán)境條件,易受水文、季節(jié)、棲息地、餌料等因素的影響[27-30];參數(shù)b反映魚類生長狀態(tài),表示魚類生長的不均勻性[30],這種不均勻性是由體質(zhì)量(或體積)和體長的不均勻增長帶來的,如果體長、體高和體寬為等速生長,比重不變,則b=3或接近3[20-21]。b值大小與魚類生長發(fā)育狀態(tài)密切相關,幼魚的b值多小于3,呈強異速性生長,隨魚體生長發(fā)育,異速性減弱,發(fā)育趨于均勻,成魚時b值接近或大于3,其生長狀態(tài)可描述為:異速生長—等速生長—異速生長[27]。
本研究中,叉長-體質(zhì)量關系式W=1.303 84×10-6L3.38156,b值大于3,表明西北太平洋公海日本鯖處于正向異速生長狀態(tài),即體質(zhì)量增長速度快于叉長,能夠較好育肥。受生長階段、餌料水平、攝食效率、性腺發(fā)育程度和產(chǎn)卵頻率等因素的影響[28],參數(shù)a和b的值也一直都處于變化中,本研究的參數(shù)a和b均處于表3所列舉相關報道的a(6.55×10-7~1.66×10-5)和b(2.95~3.59)變化范圍內(nèi)。FROESE[28]認為,b值遠小于或者遠大于3(b<2.5或b>3.5)一般都不具代表性,可能是樣品數(shù)量少或者長度范圍小所致。據(jù)報道,日本鯖叉長-體質(zhì)量關系中參數(shù)b的正常范圍為2.7~3.7[4],而北太平洋日本鯖的b值大多時候均大于3(表3),表明該海域日本鯖的體質(zhì)量增速快于叉長。與以往的記錄相比較(表3),本研究中日本鯖叉長的范圍更廣(102~400 mm),時間跨度更長,樣品量更大,研究所得的日本鯖叉長-體質(zhì)量關系式應更為合理。因此,本研究結果對于西北太平洋公海日本鯖漁業(yè)資源的可持續(xù)利用和保護管理是一個十分有價值的基本參考。但是在日本鯖生物學的諸多研究中[1,4,16,31-35],人們很少捕獲到小于100 mm的樣品,假如小個體缺失,是否會對擬合的叉長-體質(zhì)量關系產(chǎn)生影響?利用這些缺失小個體樣品的叉長、體質(zhì)量或者年齡作為參數(shù)的資源評估結果是否值得信任?這些問題尚需未來進一步研究解決。

表3 不同時期日本鯖叉長-體質(zhì)量關系參數(shù)Tab.3 Parameters of relationship between fork length and weight of Scomber japonicus
在本研究中,叉長-體質(zhì)量關系(圖5)及參數(shù)a和b(表2)各年份間雖有差異,但是并無明顯的變化規(guī)律,然而,其季節(jié)性變化特征十分明顯。參數(shù)b隨季節(jié)變換而變化,最小值和最大值分別為3.21(春季)和3.53(冬季)(表2),當個體叉長一定時,體質(zhì)量由春季到冬季依次增大(圖5),即體質(zhì)量增速與叉長增速的比值隨之逐漸增大。日本鯖具有產(chǎn)卵洄游、索餌洄游、越冬洄游等長距離洄游特性[36],這可能是導致參數(shù)b季節(jié)性變化的重要原因。西北太平洋公海日本鯖的產(chǎn)卵期為每年的3—6月[36],整個春季都存在繁殖個體。魚類在繁殖期攝食強度降低[37],部分肌肉組織在性成熟過程中將會轉(zhuǎn)變?yōu)樾韵伲?8]。因此,魚類體質(zhì)量的增加在春季將會變緩甚至停止,b值降低。此外,漁場重心的季節(jié)性變化可能是影響b值變化的另外一個原因,有研究表明,西北太平洋公海日本鯖漁場重心具有明顯的季節(jié)性變化特征[11]。
3.2 性比及性腺成熟度組成
魚類的雌雄比受多種因素諸如個體大小、海域、年份、季節(jié)等因素制約[38]。在繁殖季初期,日本鯖雌雄個體數(shù)量基本相當,繁殖旺季雌雄比達到最大值,其后逐漸下降,產(chǎn)卵末期雌性個體明顯少于雄性[39]。個體規(guī)格顯著影響雌雄比,小個體雌雄比基本符合1∶1,大個體群體中雌性顯著多于雄性[39],在魚類中這種現(xiàn)象較為普遍,其他魚種諸如東黃海的黃鮟鱇(Lophiuslitulon)[40]和藍點馬鮫(Scomberomorusniphonius)[41]也符合這個規(guī)律。日本鯖雌性個體在產(chǎn)卵期產(chǎn)卵數(shù)次,每次產(chǎn)卵數(shù)量為5萬~9萬粒,據(jù)推測,為了使自身體腔能夠容納更多數(shù)量的卵子,性成熟后雌性個體會保持更高的生長速率,以此來達到維持種群數(shù)量的目的[39]。本研究中,日本鯖雌雄比為1.74,莊之棟等[16]、戴澍蔚[17]得出的雌雄比也均大于1,表明該海域雌性個體數(shù)量多于雄性。在繁殖季節(jié),雌性會通過多次產(chǎn)卵來達到補充群體豐度的目的,因此,據(jù)推測雌性個體多于雄性個體可能是一種維持種群數(shù)量的手段[42]。
魚類性腺發(fā)育具有較強的季節(jié)性,春季是西北太平洋公海日本鯖的產(chǎn)卵期,因此春季性腺達到成熟的個體比例相對較高,但是筆者發(fā)現(xiàn)這種現(xiàn)象并不明顯(圖6)。西北太平洋公海日本鯖具有距離洄游特性,3—6月在日本大陸沿岸海域產(chǎn)卵,隨后向西北進行索餌洄游[43-44],在生殖洄游過程中大部分產(chǎn)卵個體在日本近海被消耗,這可能是造成索餌場內(nèi)性成熟度高的個體數(shù)量少的重要原因之一。本研究中,性成熟度Ⅱ期個體占絕對優(yōu)勢,Ⅳ期、Ⅴ期和Ⅵ期個體數(shù)量少,特別是Ⅴ期和Ⅵ期,占比僅為0.30%。莊之棟等[16]發(fā)現(xiàn),2016年8—10月北太平洋公海日本鯖的性腺成熟度全部為Ⅰ~Ⅱ期。因此,西北太平洋公海海域內(nèi)日本鯖鮮見繁殖行為。本研究中,叉長顯著影響日本鯖性腺發(fā)育程度,隨著叉長增加,Ⅰ期、Ⅱ期占比逐漸下降,Ⅲ期、Ⅳ期占比逐漸升高。
日本鯖初次性成熟時間取決于其叉長而并非年齡,性腺在其叉長達到一定值后即開始發(fā)育成熟[45]。日本鯖初次性成熟的叉長具有較大的時空差異[46-47],本研究發(fā)現(xiàn),西北太平洋公海雌性個體初次性成熟叉長為(298.08±4.32)mm,與20世紀50年代到60年代我國近海結果接近[48-49](表4)。目前,近海海域內(nèi)日本鯖性成熟提前及初次性成熟叉長減小已成普遍現(xiàn)象,但是筆者發(fā)現(xiàn),西北太平洋公海日本鯖初次性成熟叉長與歷史數(shù)據(jù)對比并沒有減小趨勢,可能顯示該海域內(nèi)日本鯖資源并未受到捕撈壓力的顯著影響。
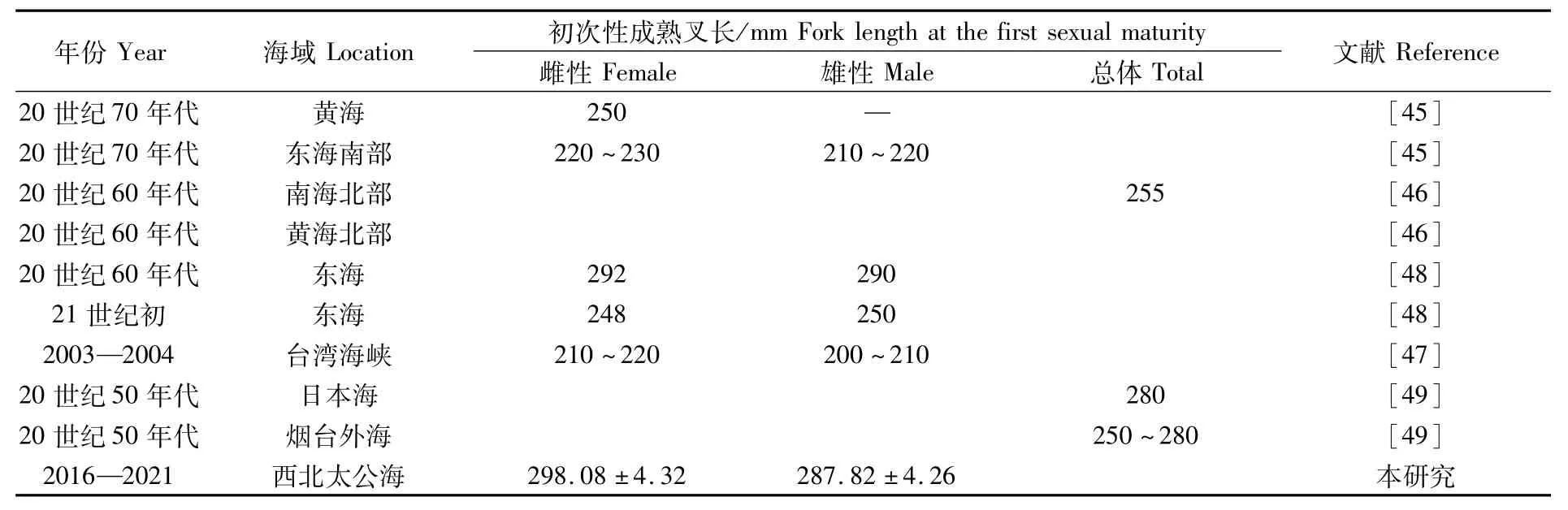
表4 日本鯖初次性成熟叉長Tab.4 Fork length at the first sexualmaturity of Scomber japonicus
3.3 攝食等級與肥滿度
日本鯖的攝食餌料隨個體增大而變化,種類先增加后減少,并由小個體逐漸向大個體轉(zhuǎn)變[15]。這種現(xiàn)象符合“最佳攝食理論”,可以讓攝食者得到最大的食物凈值。在魚類攝食中,攝食強度與叉長存在多種關系:正相關[50]、負相關[51]或沒有顯著差異[52]。日本鯖活動能力強,洄游路線長,其攝食等級和攝食種類具有明顯的季節(jié)性變化。這種變化往往與水溫有直接關系,水溫對于魚類的攝食具有重要影響,攝食強度隨著水溫降低而減小[53],溫度降低時,一部分能量轉(zhuǎn)為脂肪用來保持體溫,此時攝食強度低[15]。但是在本研究中,筆者發(fā)現(xiàn)日本鯖的攝食強度從春季到冬季依次增強,空胃和1級的個體占比逐漸減少,其他攝食等級個體占比逐漸增加,這可能與日本鯖的洄游有關,本研究海域內(nèi)日本鯖漁場重心在每年的8—9月由東北向溫度較高的西南移動[11],說明日本鯖在此范圍內(nèi)洄游。且本研究顯示日本鯖攝食強度以1~4級為主,空胃個體少,此前有研究指出,西北太平洋公海日本鯖的攝食等級以2~4級為主[16],本文結果與其相似,表明西北太平洋公海可以基本滿足日本鯖的餌料需求。肥滿度是用來表征魚類肥瘦程度和生長情況的指標,可用來分析魚類生長速度、成活率和總體增重程度,同時也可用來間接評價生活環(huán)境狀況[54]。本研究發(fā)現(xiàn),叉長顯著影響日本鯖肥滿度,隨叉長增加肥滿度逐漸增大,體質(zhì)量增加快于叉長,表明日本鯖在該海域能夠較好的育肥。結果表明,該海域能夠為日本鯖提供良好的覓食及育肥條件,是其生長發(fā)育的優(yōu)良漁場。
3.4 不足與展望
1)鑒于目前存在關于西北太平洋公海存在多種日本鯖種群的猜想,本研究僅初步鑒別了研究海域的樣品種群,未能結合更多方法做出有效認定,這也是后續(xù)有待深入研究的問題。
2)本研究發(fā)現(xiàn),大個體中雌性占比較高,未來可針對此現(xiàn)象進一步探討原因。
魚類的生物學特征是進行深入研究的基礎,由于我國對西北太平洋公海捕撈的規(guī)范化管理起步較晚,同時科學調(diào)查存在成本高、采樣困難等問題,該海域諸多魚種的生物學特征及其諸如年齡與生長、生活史、種群劃分等方面都有待深入探討。在后續(xù)研究中,筆者將首要明晰西北太平洋公海日本鯖年齡結構組成,進一步地進行資源評估,并分析其生活史,對其生長發(fā)育、繁殖產(chǎn)卵、攝食、洄游等進行深入研究,為合理開發(fā)利用該海域日本鯖資源提供科學指導,并為我國參與NPFC的相關工作提供理論支撐,提高我國在NPFC中的話語權。
猜你喜歡
中學生數(shù)理化·中考版(2022年10期)2022-11-10 09:37:42
中學生數(shù)理化·八年級物理人教版(2022年12期)2022-02-14 07:08:42
中學生數(shù)理化·八年級物理人教版(2021年12期)2021-12-31 03:23:08
中學生數(shù)理化·中考版(2020年10期)2020-11-27 01:59:48
中國生殖健康(2019年2期)2019-08-23 08:12:08
石油化工建設(2018年6期)2018-04-22 03:16:54
產(chǎn)品可靠性報告(2017年7期)2017-09-05 09:49:12
中學生數(shù)理化·八年級物理人教版(2017年12期)2017-04-18 12:59:38
汽車觀察(2016年3期)2016-02-28 13:16:26
民生周刊(2014年7期)2014-03-28 01:30:54
