天山北麓4個山葡萄品種枝條抗寒性綜合評價
2023-09-21 13:01:30潘越劉珩王明韓政偉郭靖張浩王季姣馬勇杜林峰
中外葡萄與葡萄酒 2023年5期
潘越,劉珩,王明,韓政偉,2,郭靖,2,張浩,2,王季姣,4,馬勇,4,杜林峰
(1. 新疆林業科學院,新疆烏魯木齊 830063;2. 新疆林業科學院佳木國家重點林木良種基地,新疆溫宿 843100;3. 新疆阿克蘇森林生態系統國家定位觀測研究站,新疆溫宿 843100;4. 新疆農業大學林學與風景園林學院,新疆烏魯木齊 830052;5. 昌吉州林業技術推廣中心,新疆昌吉 831100)
山葡萄是葡萄屬中最抗寒的一個種[1],新疆獨特的區位地理及環境因素,使山葡萄具備很大發展潛力。自2019年以來,新疆林科院陸續從吉林地區引進抗寒性狀表現優異的10余個山葡萄品種, 經馴化、區域栽培試驗,從中篩選出4個適應性良好的品種,目前栽培面積穩中有升,產量持續增加。沿天山北麓地帶光照充裕,空氣干燥,晝夜溫差大,有利于糖分累積,但受春季回溫加速和冷暖交織加劇影響,葡萄出土后極易遭受霜凍災害[2-3],影響釀酒葡萄產業的健康發展,為此篩選出高抗寒的品種對該區域的釀酒葡萄產業發展具有重大意義。
低溫脅迫是一種限制植被生長與分布的非生物脅迫因素。關于植物抗寒性的研究多集中在通過測定相對電導率和Logistic方程推算半致死溫度的方法,對葡萄抗寒性的研究也有很多,如賈金輝等[4]以8個釀酒葡萄為試材,通過測量一年生枝條相對電導率,基于隸屬函數值和半致死溫度綜合排序,得出‘雙豐’抗寒性最強;劉鈺璽等[5]基于主成分分析對葡萄抗寒性綜合評價時認為,評價結果與半致死溫度LT50存在極顯著相關性,通過計算半致死溫度,可有效判別不同品種間的抗寒性。同時,通過探究滲透調節物質丙二醛[6]、游離脯氨酸[7]和保護酶系統[8]等與植被抗寒能力的差異性, 亦可間接反映植被的抗寒水平。目前,關于葡萄的抗寒性及品種的生物學特性研究很多,但是對天山北麓引進的山葡萄抗寒生理評價方面的研究報道較少。
為進一步探索山葡萄在天山北麓生態區域的抗寒生理特性,試驗以‘北冰紅’等4個適應性良好的品種為試材,對一年生休眠枝條的抗寒生理指標及萌芽率進行調查,綜合評價各個品種的抗寒性,為天山北麓生態區域篩選自然越冬的高抗寒山葡萄品種提供理論依據。
1 材料與方法
供試的4個山葡萄品種(北冰紅、雙紅、左優紅、雪蘭紅)一年生苗于2019年秋從中國農科院特產所(左家)引進,窖內保存, 2020年3月于溫室內煉苗,5月種植在新疆烏魯木齊三坪農場山葡萄資源匯集圃。于2021年12月中旬,每個品種隨機選取5株樹,采集充分成熟、無病蟲害、長勢一致的一年生枝條。所有枝條的朝向及位置盡量保持一致,使用凡士林涂抹剪口,根據品種分別裝入聚乙烯保存袋中,于0~4 ℃冰箱內保存。
1.1 試驗處理
采回的一年生枝條分段,剪成10 cm的長度,自來水沖洗干凈,再用離子水沖刷3次,吸干枝條表面水分,每個品種分為5份,紗布包裹后置入超低溫冰箱進行低溫處理。共設置0、-10、-20、-30、-40 ℃ 5個梯度,冷凍降溫幅度及解凍升溫幅度均保持5 ℃·h-1,到達預設溫度時維持12 h后解凍,測量相關生理指標,每組處理3次重復。
1.2 試驗方法
1.2.1 生理指標的測定
相對電導率的測定。將低溫處理后的枝條剪成2~3 cm的小段,避開芽眼,稱取1 g置入三角瓶內,加入20 mL重蒸餾水,放置搖床室溫環境浸提10 h,用電導儀(型號:DDS-11A)測定此時電導率(R)。用保鮮膜將三角瓶瓶口封好置入沸水浴中蒸煮30 min后浸提10 h,測量此時電導率R0。每個樣品設置3次重復,計算公式如下:
游離脯氨酸(Pro)采用酸性茚三酮法測定[9];丙二醛(MDA)采用硫代巴比妥酸法測定[10],在450、532、600 nm分別測量吸光度值,計算含量;可溶性糖采用蒽酮乙酸乙酯法測定[11];超氧化物歧化酶(SOD)活性采用氮藍四唑(NBT)還原法測定[11]。
1.2.2 萌芽率的測定
枝條經不同低溫梯度處理12 h后扦插于溫室(25~28 ℃)營養缽內,每個營養缽扦插6根枝條,以體積比2∶1的草炭土和鋸末作為基質,每組處理重復3次,1個月后統計枝條芽體萌發情況,以破磷露白為標志,萌芽率(%)=萌發芽眼數/扦插枝條芽眼總數×100。
1.3 數據分析
1.3.1 半致死溫度的計算[12]
各品種半致死溫度LT50擬合曲線方程為:
其中,Y為相對電導率,a、b是方程參數,K是常數,此處取值100。LT50為方程拐點,計算公式如下:
1.3.2 主成分分析
主成分分析是將多項復雜指標簡化為少數綜合指標的統計學方法,在保證原始信息損失少且減少變量數目的前提下,采用少數變量反映原始信息[13-15]。
在對山葡萄枝條抗寒能力進行綜合評價之前,考慮到各指標數量綱不一致,其中MDA為達到峰值時的溫度,其值越低越好,萌芽率、SOD、游離Pro、可溶性糖和半致死溫度均為值越高越好,為此在進行主成分分析前,基于隸屬函數法對原始數據進行標準化處理。
設X=(X1,X2,…Xn)',設n為隨機向量,其線性關系式如下:
將PC1代替原來p個變量X1,X2,…,Xp,PC1會盡可能多的反映原始變量的信息,如若PC1不足以反映原始的絕大多數信息,則考慮引入PC2,據此類推。主成分分析成分個數m需依據各個成分的累計方差貢獻率最終確定。
式中,λ為各成分所對應的特征值;k為選定的成分數;i為全部成分數。
主成分分析,通過主成分載荷矩陣Ai和特征向量λ計算各成分系數矩陣Ti,計算公式如下:
其中,A為不同山葡萄品種的成分值,C為各成分的貢獻率(i=1,2,…)。主成分分析采用SPSS 21.0進行。
2 結果與分析
2.1 不同低溫處理下山葡萄相對電導率的變化
由圖1可見,經不同梯度低溫處理后,4個山葡萄品種的相對電導率與處理溫度呈“S”型上升趨勢,各品種的相對電導率在0~-20 ℃緩慢上升,其中‘左優紅’上升幅度最小,‘雙紅’次之,‘北冰紅’上升幅度最大;在-20~-30 ℃表現為急速上升狀態,在-30 ℃時,‘雙紅’相對電導率顯著低于其他品種;在-30~-40 ℃,各品種上升幅度趨于平緩。4個山葡萄品種在相同低溫處理下的相對電導率差異較大,伴隨溫度降低,相對電導率上升幅度不一,增幅在56.95%~65.94%,這主要與品種間抗寒能力的差異有關。
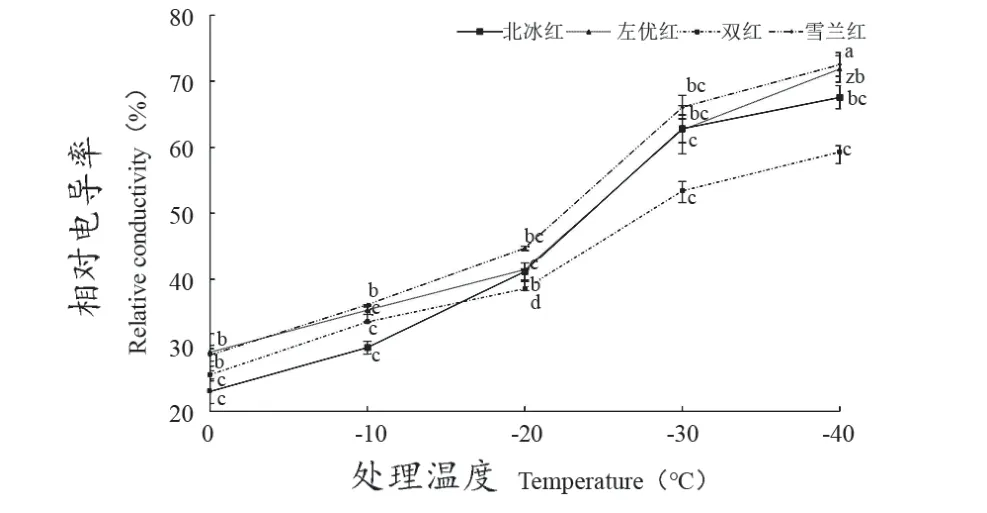
圖1 不同低溫處理后4個山葡萄品種枝條相對電導率的變化Figure 1 Changes in relative conductivity of four Vitis amurensis Rupr. branches with different temperatures treatments
2.2 半致死溫度的比較
根據相對電導率,基于Logistic方程擬合得出不同山葡萄品種的半致死溫度。不同低溫梯度處理下材料的Logistic方程、擬合度及半致死溫度見表1。4個山葡萄品種的半致死溫度在-29.193~-20.236 ℃,擬合度均大于0.900,達到極顯著水平。由于半致死溫度與植物抗寒能力呈負相關,抗寒性越強,半致死溫度越低[16]。由此得出抗寒性由高到低排序依次是:雙紅>北冰紅>左優紅>雪蘭紅。
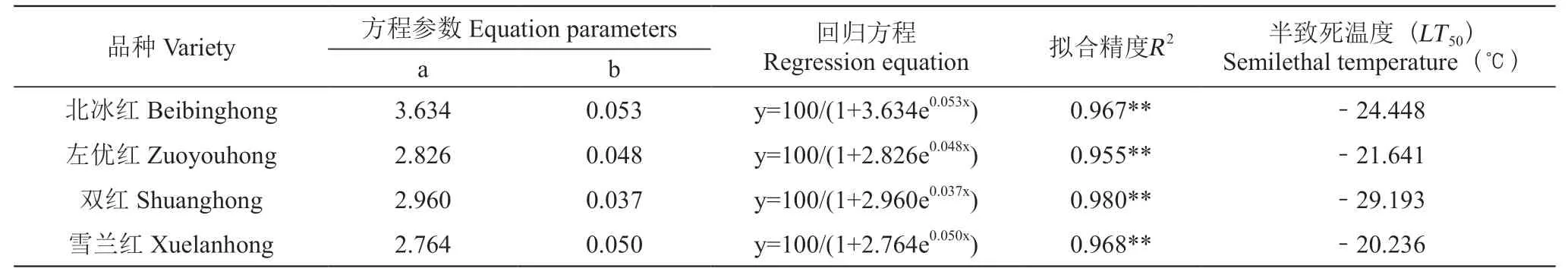
表1 山葡萄枝條Logistic方程及半致死溫度Table 1 The relative conductivity Logistic equation parameter and semilethal temperature (LT50) of Vitis amurensis Rupr. shoots
2.3 低溫脅迫下各品種枝條抗寒生理指標變化
表2可以看出,從0~-30 ℃,MDA、游離Pro、可溶性糖含量和SOD活性均隨著溫度的降低而增加,枝條萌芽率則與之相反。‘左優紅’在-20、-30、-40 ℃時, MDA含量顯著高于其他3個品種;‘北冰紅’‘左優紅’‘雙紅’和‘雪蘭紅’游離Pro的最大值分別是55.46、45.38、53.23、49.47 μg·g-1。多重比較可知,‘北冰紅’與‘雙紅’差異不顯著,但均顯著高于‘左優紅’;4個品種的SOD活性均在-30 ℃達到最高,由高到低排序依次是北冰紅>雙紅>左優紅>雪蘭紅;4個品種可溶性糖含量均伴隨脅迫溫度的降低而增加,均在-40 ℃達到最大值;枝條萌芽率方面, ‘北冰紅’和‘雙紅’在-10~-30 ℃區間內萌芽率下降幅度較小,‘左優紅’和‘雪蘭紅’下降幅度較大,‘左優紅’和‘雪蘭紅’在-30 ℃時的萌芽率顯著低于-10 ℃和-20 ℃,4個品種均在-40 ℃時萌芽率最低。
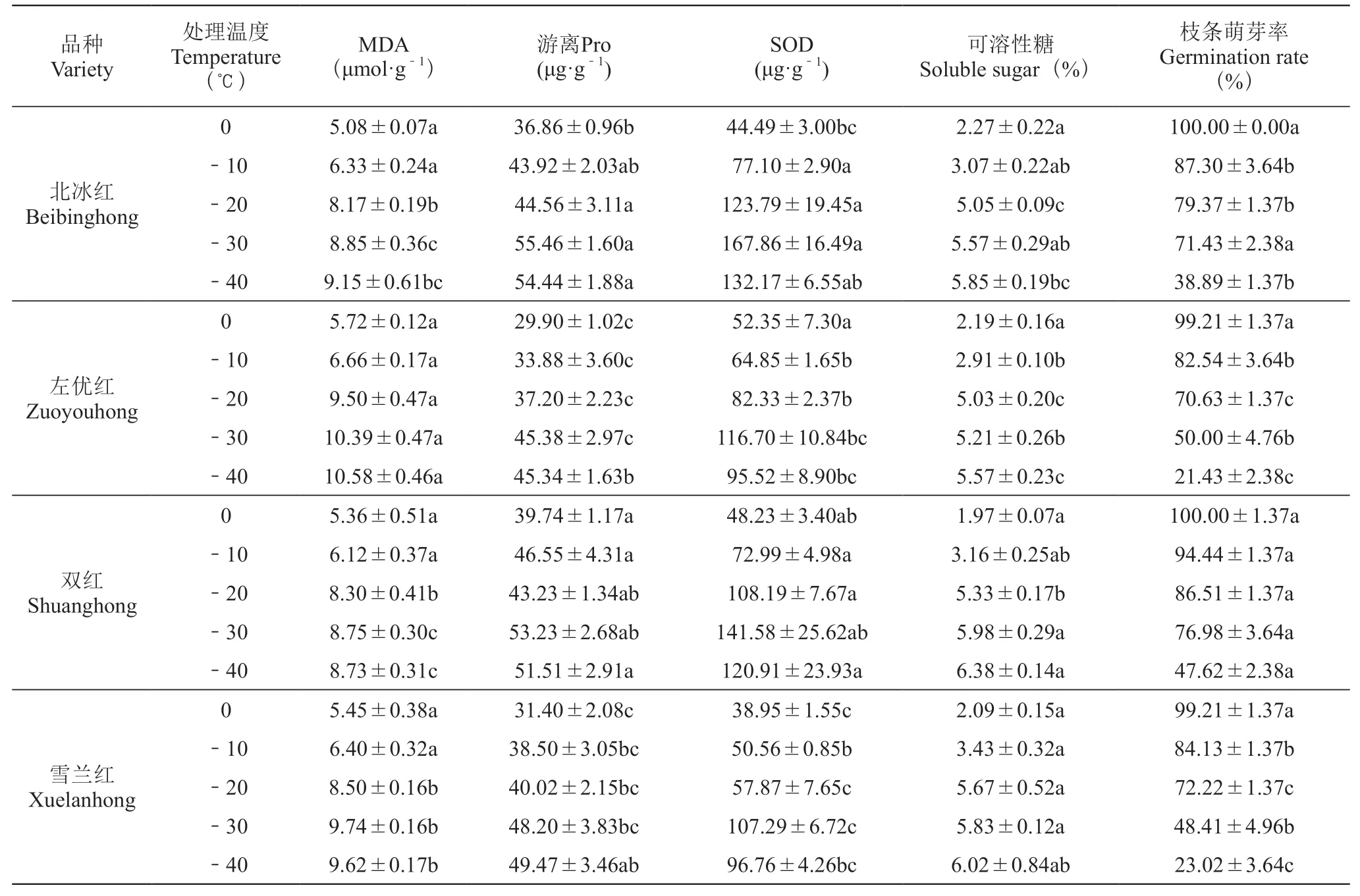
表2 低溫條件對山葡萄枝條抗寒生理指標及萌芽率的影響Table 2 Changes of MDA content、Pro content、 SOD activity 、Soluble sugar and Germination rate in branches from different Vitis amurensis Rupr. under low temperature stress
2.4 山葡萄枝條各項生理指標的主成分分析
從表3中看出,前兩個主成分的特征根>1,且累計解釋方差達95.072%,包含了原始數據的絕大部分信息,為此提取前兩個主成分進行分析。
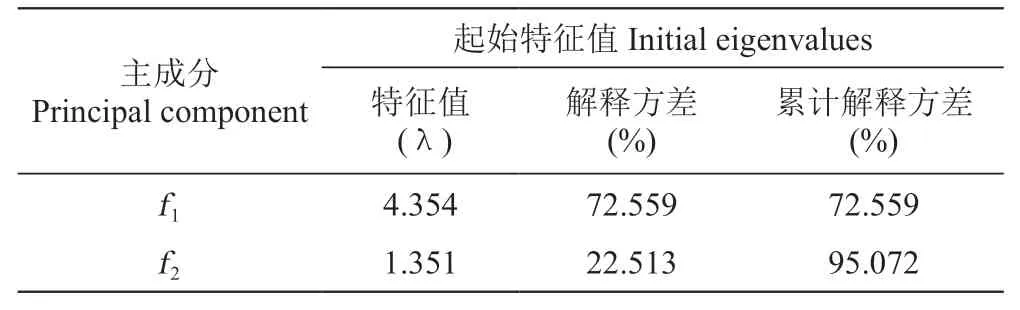
表3 主成分特征值Table 3 Component eigenvalue
為突出主要因子對評價結果的影響,對公因子進行最大正交旋轉,得到的旋轉載荷矩陣見表4。主成分f1載荷值較高的抗寒指標包括萌芽率、半致死溫度、脯氨酸和SOD,載荷權數依次為:0.940、0.892、0.868和0.951,這4項指標對第1主成分產生正向影響。
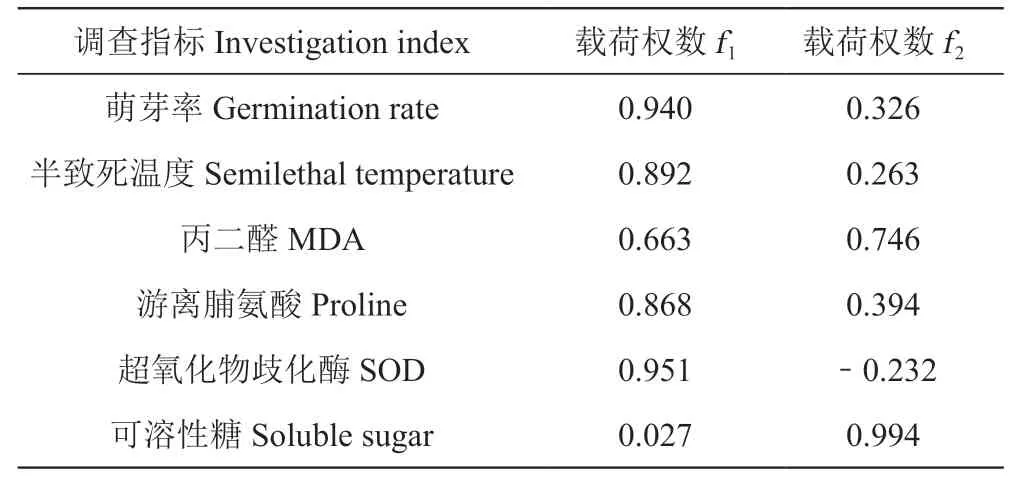
表4 旋轉后的主成分載荷矩陣Table 4 Rotated principal component loading matrix
第2主成分載荷值較高且符號為正的指標包括:丙二醛和可溶性糖,載荷權數依次是0.746和0.994,這2項指標對第2主成分產生正向影響。
用各指標的主成分載荷除以與主成分對應的特征根,開算數平方根,得出2個主成分每項指標對應的系數(即為特征向量),將特征向量與測得的數據結果相乘,即可得出2個主成分的函數表達式(ZX為標準化后的結果):
上述表達式中,X1為萌芽率、X2為半致死溫度、X3為丙二醛、X4為脯氨酸、X5為SOD、X6為可溶性糖。
以各主成分的特征值占特征值總和的比例作為權重,得出綜合評價函數fz=A1f1+A2f2,其中A1=λ1/(λ1+λ2),A2=λ2/(λ1+λ2)。λ1和λ2分別為2個主成分的特征值。表5所示,綜合得分由高到低排序依次為:雙紅>北冰紅>雪蘭紅>左優紅。
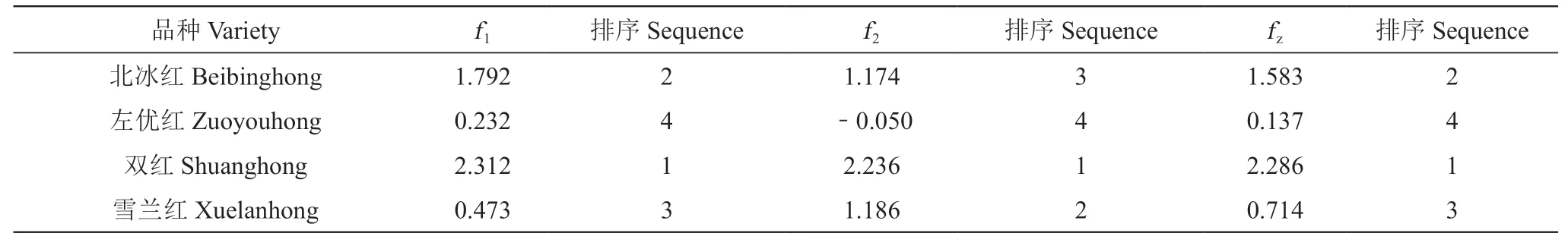
表5 不同山葡萄品種主成分得分、綜合得分及排序Table 5 Scores of each principal component, composite scores and sequence of different Vitis amurensis Rupr. varieties
3 討論與結論
植被抗寒機理較為復雜,評價一個品種的抗寒能力無法單獨通過某一個指標來確定,而應通過多項指標進行全面綜合評判[17]。Lyons等[18]發現,植被遭受凍害時,受害程度較低或抗寒性較強的品種不但電導率增加幅度較小,且細胞滲透性變化可以逆轉;受害程度較重或抗寒性較差的品種則反之。本研究發現,伴隨處理溫度的降低,不同山葡萄品種的電導率逐漸升高,通過Logistic曲線方程模擬了4個山葡萄品種的LT50范圍在-29.193~-20.236 ℃,天山北麓生態區域冬季平均氣溫-10.6 ℃,平均最低氣溫-24.6 ℃,按照本試驗判斷‘北冰紅’和‘雙紅’均可自然越冬。然而生產上天山北麓生態區域山葡萄自然越冬下,仍表現出干枯、抽梢凍害等現象,推測引起釀酒葡萄產生凍害的原因不僅與當年低溫有關,還與當年降雪覆蓋量、控水控肥和開春時干熱風發生頻率有關。因此為防止凍害發生,生產上宜通過鋪設地膜提高地溫等抗寒輔助措施進行預防。
MDA是植物在受到低溫凍害時產生的過氧化產物,會對細胞產生一定的毒害作用[19-20]。游離Pro可以調節細胞的滲透壓,防止細胞過度脫水進而增強植被抗寒性[6,21]。本研究發現,‘雙紅’的MDA含量較小,抗寒性較強,‘北冰紅’和‘雙紅’游離Pro含量較高,適應低溫環境的能力優于其他兩個品種。可溶性糖作為蛋白質和細胞膜的重要保護劑,其積累量越高越能幫助植被免于凍害[22-23]。本研究中的4個山葡萄品種隨著溫度降低,對外界環境做出應激反應,從而使可溶性糖含量持續增加,出現這種情況主要是由于低溫脅迫之初,山葡萄對外界不良環境做出的應激反應,使可溶性糖含量增加;當溫度過低造成細胞死亡,可溶性糖含量下降。
植被活性氧清除系統包括酶促及非酶促抗氧化系統,酶促系統包含過氧化物酶、超氧化物歧化酶、過氧化氫酶、脫氫抗壞血酸還原酶和谷胱甘肽還原酶等[5,24],非酶促抗氧化系統主要由還原性谷胱甘肽和抗壞血酸構成。本研究發現,‘北冰紅’和‘雙紅’的SOD表現優于‘雪蘭紅’和‘左優紅’。楊盛[25]等研究發現,低溫脅迫下,植物體酶活高于正常水平。本研究供試的4個山葡萄品種酶活性變化幅度明顯不同,抗寒性強的品種在受低溫脅迫后酶活性增加幅度大,抗寒性弱的品種反之,且隨著低溫加劇,在到達-40 ℃后酶活性迅速降低,由此證明在輕度低溫脅迫環境下,山葡萄可通過增加酶活性減輕脅迫傷害,在脅迫程度超過保護酶系統清除能力時,氧化酶活性開始下降。
為了更全面地對植被的抗寒性進行評價,避免單一指標鑒定的不足,科研工作者先后提出了隸屬函數法[26]、直接比較法[27]和分級比較法[5]等。劉鈺璽等[5]對不同葡萄砧木抗寒性進行了主成分分析和聚類分析,所得結果與田間觀察基本一致。本研究基于主成分分析法得出, 4個山葡萄品種的抗寒性由高到低依次為:雙紅>北冰紅>雪蘭紅>左優紅,與Logistic曲線方程模擬半致死溫度結果大致吻合。
