沉默NCAPH基因對子宮內膜癌細胞惡性生物學表型影響的研究
2023-09-25 08:45:28楊夏楠趙丹丹楊平娟樊冬梅
黑龍江醫藥 2023年17期
關鍵詞:水平
楊夏楠,趙丹丹,司 晗,楊平娟,樊冬梅
河南科技大學第一附屬醫院婦三科,河南 洛陽 471003
子宮內膜癌(Endometrial Cancer,EC)是女性中第四大常見癌癥,也是最常見的婦科癌癥。子宮內膜癌是一種具有侵襲或轉移能力的惡性腫瘤,主要發生在更年期和絕經后婦女中[1]。子宮內膜癌根據病因和臨床行為可以分為I 型和II 型。I 型腫瘤是雌激素依賴性高分化腺癌,起源于子宮內膜增生,占子宮內膜癌病例的80%。與I 型腫瘤相反,II 型腫瘤是不依賴雌激素的、分化差的腺癌,其起源于萎縮性子宮內膜[2]。在我國,隨著社會的發展和經濟條件的改善,子宮內膜癌的發病率亦逐年升高,因此,闡明子宮內膜癌的發病機制對提高子宮內膜癌患者的生存時間至關重要。
非SMC 縮合蛋白I 復合物亞單位H(non-SMS condensin Ⅰcomplex subunit H,NCAPH),稱為凝聚素,在有絲分裂和減數分裂過程中參與染色體的組裝和分離,對維持染色體的結構和分離至關重要[3]。近年來,研究發現,NCAPH 在多種腫瘤的發生和發展過程中起重要作用[4]。Kim 等[5]發現,NCAPH 在胰癌的臨床組織和細胞系中表達上調,且沉默NCAPH 可以通過激活DNA 損傷反應(Chk1/Chk2)信號通路誘導染色體的畸變和DNA損傷,進而抑制胰癌細胞的增殖和誘導胰癌細胞的凋亡[5]。Sun等[6]發現,超表達NCAPH 可以促進肝細胞癌細胞的增殖、遷移和侵襲,表明NCAPH 可以作為一種新的預后生物標志物和肝細胞癌患者的潛在治療靶點。Cui 等[7]發現,NCAPH 的表達水平在前列腺癌中顯著上調,且NCAPH 的過表達與前列腺癌患者的不良生存率有關。Yin等[8]發現,NCAPH 在結腸癌組織中的表達水平明顯高于其在相鄰非癌組織中的表達水平,且沉默NCAPH 可以抑制結腸癌細胞的增殖、遷移和異種移植腫瘤的形成。然而,NCAPH在子宮內膜癌中的具體作用至今尚不清楚。
本研究通過構建NCAPH 沉默的細胞模型,探究NCAPH在子宮內膜癌細胞的增殖、遷移、侵襲和EMT過程中的作用,為今后子宮內膜癌的診斷和治療提供理論依據。
1 材料與方法
1.1 試驗材料
正常人子宮內膜細胞(hEEC),子宮內膜癌細胞(Ishikawa,RL95-2 和KLE)均購自武漢普諾賽生物科技有限公司;si-NCAPH 和其陰性對照si-NC 購自蘇州貝信生物技術有限公司;CCK-8試劑盒購買于碧云天生物生物技術;DMEM 培養基、Trizol 試劑盒、反轉錄試劑盒、熒光定量PCR 試劑盒均購自上海賽默飛世爾科技有限公司;Lipofectamine 2000購自Invitrogene公司。
1.2 細胞培養
將子宮內膜癌Ishikawa 和KLE 細胞接種在含有10%胎牛血清和1%雙抗的DMEM 培養基內,在恒溫37 ℃含有5%CO2的培養箱中培養,待Ishikawa 和KLE 細胞生長至培養瓶底部鋪滿細胞進行細胞傳代,收集生長狀態良好的細胞用于后續實驗。
1.3 細胞轉染
細胞轉染前一天將細胞接種到6 孔板中,待細胞轉染時長至70%的融合度,使用Lipofectamine 2000將si-NCAPH和si-NC 轉染到Ishikawa 和KLE 細胞中。轉染后繼續培養細胞48 h,然后收取細胞,通過qRT-PCR 和western blot檢測NCAPH的表達水平。
1.4 CCK-8
將轉染后的Ishikawa 和KLE 細胞重懸,然后接種到96孔板中,培養24 h、48 h或者72 h后,按照試劑盒操作步驟,每孔加入10 μL 的CCK-8 試劑,在95%空氣、5%CO2的37 ℃條件下孵育3 h,隨后使用酶標儀在450 nm 處檢測吸光度OD值。
1.5 劃痕愈合實驗
將轉染后的Ishikawa 和KLE 細胞接種到6 孔板中,待細胞長至基本融合時用10 μL 無菌槍頭垂直6 孔板劃痕,用PBS 清洗Ishikawa 和KLE 細胞3 次,然后放入37 ℃、5% CO2培養箱中培養24 h,然后在顯微鏡下觀察并隨機選取5 張視野拍照,通過Image J 圖像處理軟件量化遷移面積,計算其遷移率。
1.6 Transwell實驗
收集轉染后的Ishikawa 和KLE 細胞,用無血清的培養基將其重懸,然后將細胞懸液接種到預先涂有基質膠的Transwell 上室,下室加入含有血清的完全培養基。在37 ℃、5% CO2的條件下培養24 h后,取出Transwell小室,棄去孔內培養基,擦去上層小室內的細胞。將細胞用甲醇固定30 min,并用0.1%結晶紫染色10 min,顯微鏡下觀察并隨機選取5張視野拍照,對侵襲細胞進行計數。
1.7 RT-qPCR
通過Trizol 試劑(Takara,Tokyo,Japan)從Ishikawa和KLE細胞中提取的總RNA,使用逆轉錄試劑盒(Takara)合成cDNA,然后使用SYBR Prellix Ex Taq RT-qPCR 試劑盒(Takara)和NCAPH 特異性引物進行RT-qPCR 反應。以β-actin 作為內參基因,使用2-△△Ct方法計算基因mRNA的相對表達水平。
1.8 Western Blot
用RIPA 蛋白裂解液從細胞中提取總蛋白,使用BCA蛋白質測定試劑盒測定總蛋白質濃度,使用SDS-聚丙烯酰胺凝膠對蛋白質進行分離,并轉移到PVDF膜上,PVDF膜用5%脫脂牛奶封閉,后加入NCAPH 和β-action 抗體(稀釋比例為1∶1 000),4℃孵育過夜,再加入二抗,37 ℃孵育1 h,通過ECL 化學發光液顯影后在化學發光成像系統拍照,應用Image J圖像處理軟件進行定量分析。
1.9 統計學方法
采用Graphpad Prism 軟件進行數據統計學分析,組間數據比較采用t檢驗或者單因素方差分析(One-way ANOVA),以P<0.05為差異有統計學意義。
2 結果
2.1 NCAPH在子宮內膜癌中的表達水平
基于TCGA數據庫分析NCAPH在正常子宮內膜和子宮內膜癌中的表達水平。如圖1A和1B所示,NCAPH 在癌旁組織的表達水平明顯低于其在子宮內膜癌的表達水平,差異有統計學意義(P<0.001)。
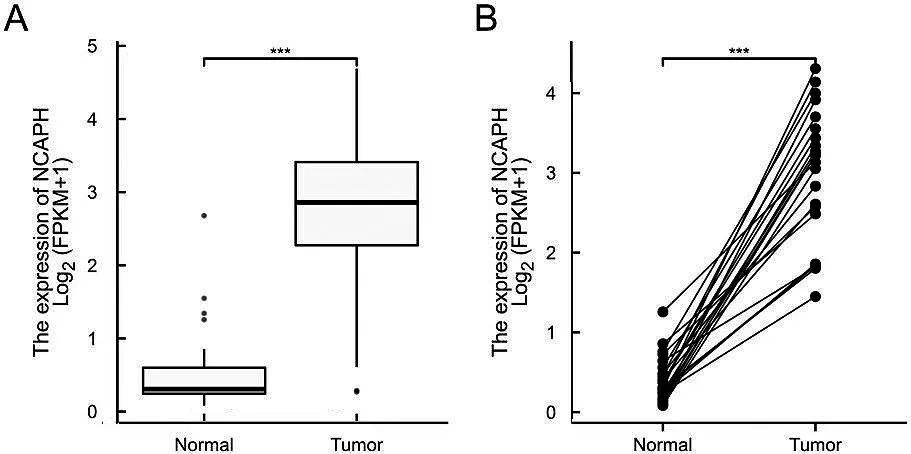
圖1 NCAPH在子宮內膜癌中的表達水平
2.2 NCAPH 在子宮內膜癌中的診斷價值及其與預后的關系
ROC 曲線分析結果顯示,NCAPH 在子宮內膜癌具有臨床診斷價值(圖2A)。此外,使用KM-plotter 在線數據庫進行生存分析,如圖1B 所示,NCAPH 高表達子宮內膜癌患者的總生存期(圖2B)和無進展生存期(圖2C)比NCAPH 低表達的子宮內膜癌患者的總生存期較短,差異有統計學意義(P<0.05)。
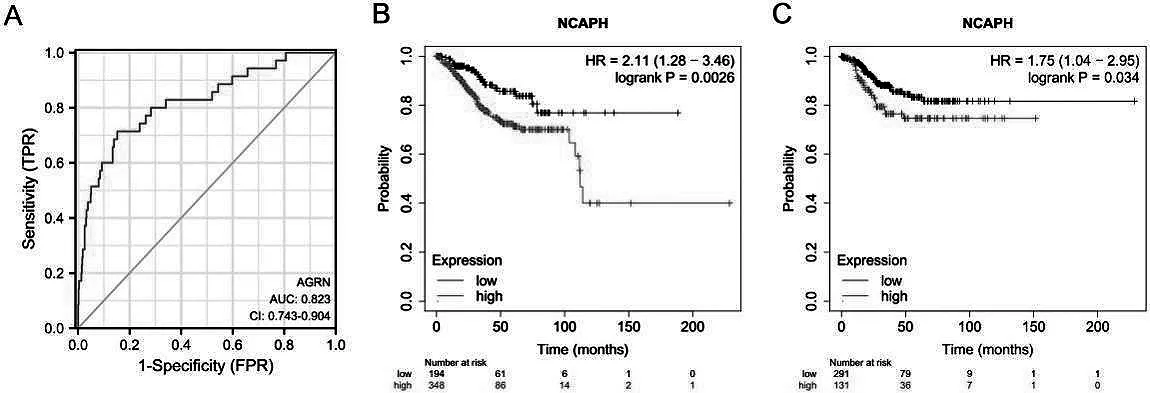
圖2 NCAPH在子宮內膜癌中的診斷價值及其和預后的關系
2.3 NCAPH 在正常子宮內膜細胞及子宮內膜癌細胞中的表達水平
使用qRT-PCR 和Western blot 實驗分析正常子宮內膜細胞hEEC 和子宮內膜癌細胞Ishikawa、RL95-2 和KLE 中NCAPH 的表達情況。由圖3 可知,與正常子宮內膜細胞hEEC 細胞相比,NCAPH 在子宮內膜癌Ishikawa、RL95-2和KLE 細胞的表達明顯升高,差異有統計學意義(P<0.001)。
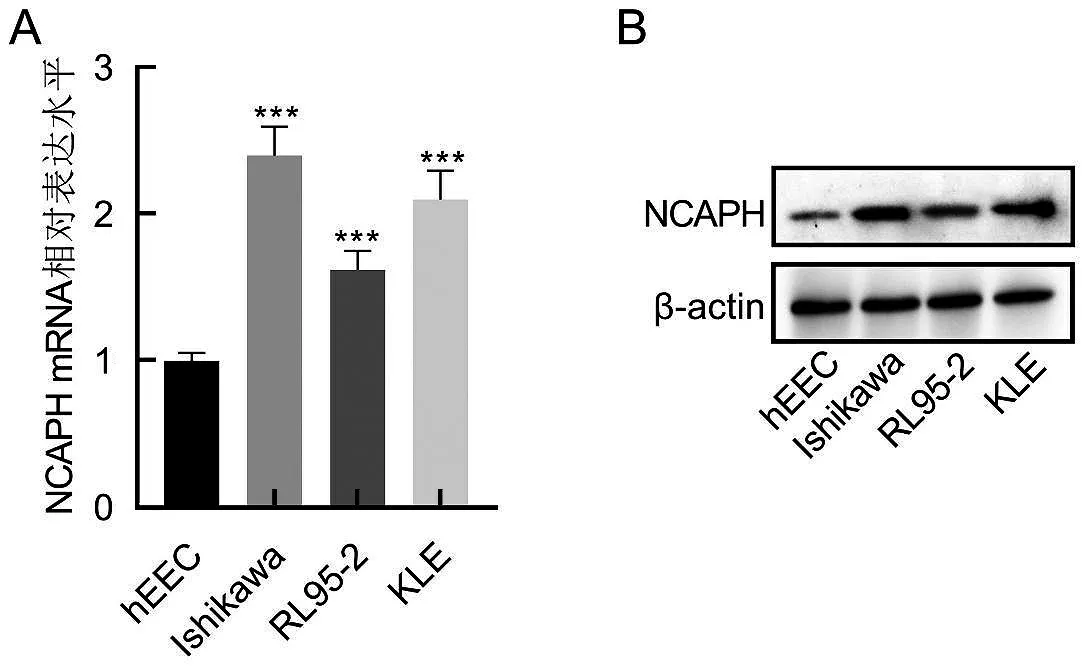
圖3 正常子宮內膜細胞及子宮內膜癌細胞中NCAPH的表達
2.4 沉默NCAPH對子宮內膜癌細胞增殖的影響
用si-NC和si-NCAPH轉染Ishikawa和KLE細胞,轉染48 h后,研究用qRT-PCR和Western Blot檢測NCAPH的表達。圖4A 和4B 結果顯示,在Ishikawa 和KLE 細胞中,與si-NC 相比,si-NCAPH 組中NCAPH 的表達明顯降低,差異有統計學意義(P<0.001)。
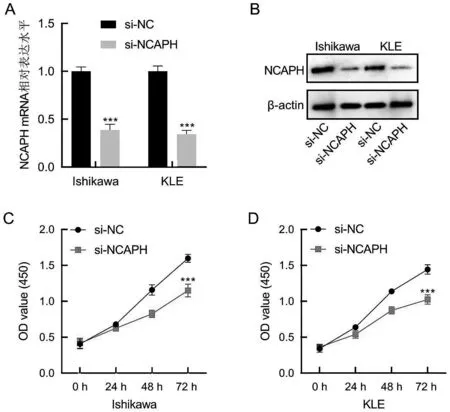
圖4 沉默NCAPH對Ishikawa和KLE細胞增殖能力的影響
使用CCK-8 方法檢測沉默NCAPH 對子宮內膜癌Ishikawa 和KLE 細胞增殖活性的影響,如圖4C 和4D 所示,與si-NC 組相比,si-NCAPH 組Ishikawa 和KLE 細胞的增殖活性明顯下降,且差異有統計學意義(P<0.001),表明沉默NCAPH可以降低子宮內膜癌細胞的增殖能力。
2.5 沉默NCAPH 對子宮內膜癌細胞遷移和侵襲能力的影響
Ishikawa 和KLE 細胞轉染si-NC 或si-NCAPH 24 h 后,研究者進行劃痕愈合實驗,評估Ishikawa 和KLE 細胞的遷移能力。如圖5A 所示,與si-NC 組Ishikawa 和KLE 細胞的遷移率相比,si-NCAPH 組Ishikawa 和KLE 細胞的遷移率顯著下降,差異有統計學意義(P<0.001)。結果表明沉默NCAPH 抑制子宮內膜癌細胞的遷移能力。與此同時,Transwell 小室實驗的結果表明,與si-NC 組相比,si-NCAPH 組Ishikawa和KLE細胞的侵襲能力明顯下降,見圖5B。由此可見,沉默NCAPH 可以抑制子宮內膜癌細胞侵襲和遷移能力。
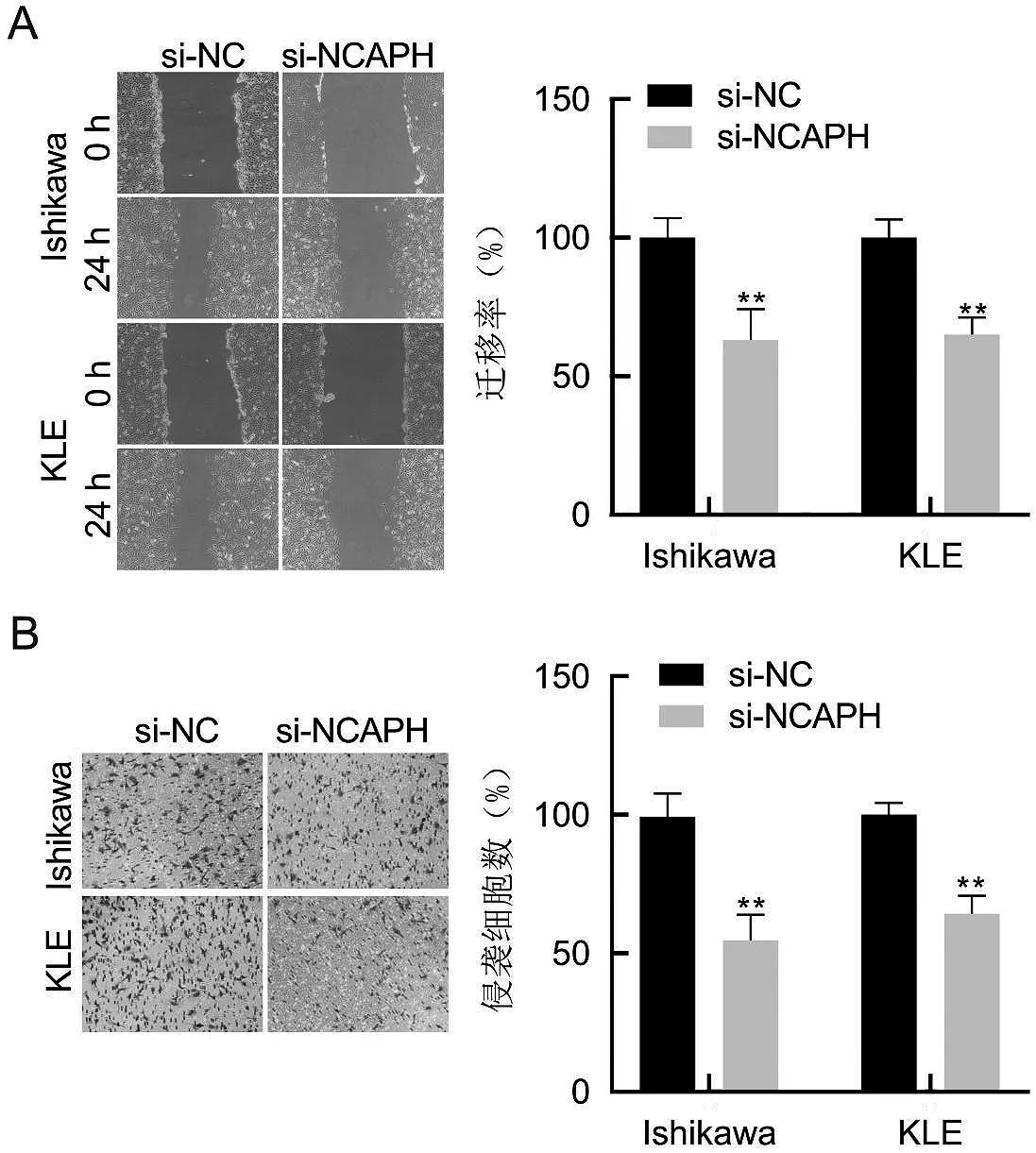
圖5 沉默NCAPH對Ishikawa和KLE細胞遷移能力的影響
2.6 沉默NCAPH對子宮內膜癌細胞中EMT的影響
為了探究沉默NCAPH 對子宮內膜癌細胞中EMT 的影響,研究使用Western Blot方法檢測了轉染48 h后Ishikawa和KLE細胞中EMT的相關基因E-鈣黏蛋白(E-cadherin)、波形蛋白(Vimentin)和N-cadherin 的蛋白表達水平。如圖6A 和6B 所示,在Ishikawa 和KLE 細胞中,與si-NC 組相比,敲減NCAPH 可以引起Ishikawa 和KLE 細胞中Ecadherin 的水平顯著升高,而引起Ishikawa 和KLE 細胞中Vimentin 和N-cadherin 的蛋白表達水平明顯下降,差異有統計學意義(P<0.001)。
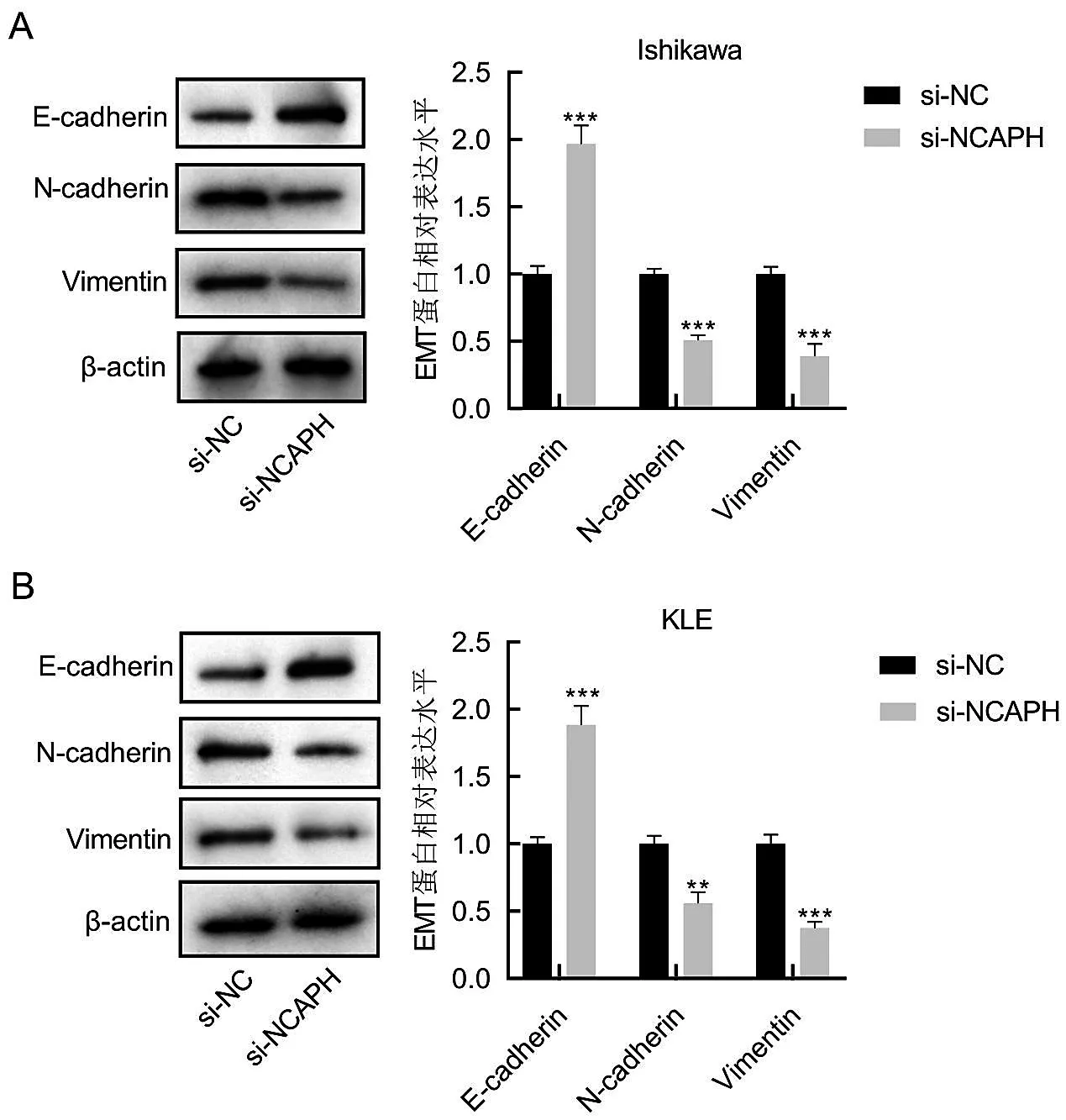
圖6 沉默NCAPH抑制Ishikawa和KLE細胞的EMT
3 討論
子宮內膜癌是最常見的女性生殖系統腫瘤之一,并是導致死亡的第三位常見婦科惡性腫瘤(僅次于卵巢癌和宮頸癌)。近年來子宮內膜癌的發病率仍在不斷升高,但是由于女性內分泌系統的復雜性,子宮內膜癌的發病機制至今尚未完全闡明。研究表明子宮內膜癌的發病機制可能與基因的異常表達密切相關[9]。因此,闡明子宮內膜癌的發病機制對提高患者的生存時間尤為重要。
已有研究發現NCAPH 在多種腫瘤的發生和發展過程中起重要作用[10]。沉默NCAPH 可以誘導胃癌細胞G1細胞周期停滯,進而抑制胃癌細胞的增殖、遷移和侵襲[11]。在漿液性卵巢癌中,NCAPH 的表達水平明顯升高,且NCAPH 的表達水平與漿液性卵巢癌的組織學腫瘤分級和淋巴轉移呈正相關,而與漿液性卵巢癌患者的總生存時間呈負相關。由此可見,NCAPH 有望成為漿液性卵巢癌的預后標志物和治療靶點。同樣地,Qi等[12]發現NCAPH 可以通過上調PI3K/PDK1/AKT通路促進卵巢癌的發展。在宮頸癌中,NCAPH 的表達水平明顯高于其在正常宮頸和高級別鱗狀上皮內病變組織中的表達水平,且其表達水平與腫瘤大小、浸潤深度和淋巴結轉移明顯相關。此外,高表達NCAPH 的宮頸癌患者的生存結果明顯比低表達NCAPH的宮頸癌患者的生存結果更好,且沉默NCAPH 可以通過PI3K/Akt/SGK 信號通路抑制宮頸癌細胞的增殖、遷移、侵襲和上皮-間充質轉化[13]。在膀胱癌中,NCAPH可以通過MEK/ERK 信號通路促進UMUC3細胞的增殖,抑制UMUC3細胞的凋亡[14]。在乳腺癌中,NCAPH 在MCF-7 和MCF-10A 細胞中的表達水平明顯升高,且其上調表達與乳腺癌患者的預后不良明顯相關,表明NCAPH 有可能作為乳腺癌的預后標志物[15]。在非小細胞癌中,NCAPH 沉默可以抑制A549 和H1299 細胞的增殖,誘導細胞周期在G/M 期停滯,并阻止A549 和H1299 細胞的集落形成、遷移和侵襲[16]。Shimomura 等[17]研究發現,NCAPH 在口腔鱗狀細胞癌中的表達高于其在正常口腔黏膜中的表達,且NCAPH 的免疫染色與淋巴結轉移和淋巴浸潤密切相關。此外,機制研究發現,NCAPH 可以促進內皮細胞向口腔鱗狀細胞癌細胞的遷移和黏附,降低口腔鱗狀細胞癌細胞對鉑類抗癌藥物的敏感性。然而,NCAPH 在子宮內膜癌中的功能作用至今尚未見報道。在此,研究通過分析TCGA和KM plotter 數據庫發現,NCAPH 在子宮內膜癌組織的表達水平明顯高于其在子宮內膜癌旁組織的表達水平,且其表達水平與患者的預后生存時間呈負相關。由此可見,NCAPH 有可能作為促癌基因,在子宮內膜癌的發生發展過程中起重要作用。這與Qiu 等[18]的研究結論一致,NCAPH 在子宮內膜癌中上調并且與較差的臨床病理學特征相關。本研究通過在子宮內膜癌細胞中沉默NCAPH 發現,沉默NCAPH 能夠導致子宮內膜癌細胞的增殖活性降低。此外,劃痕愈合實驗和Transwell實驗結果顯示,沉默NCAPH能夠顯著抑制子宮內膜癌細胞的遷移和侵襲能力。
EMT 是指細胞失去上皮特性而獲得間質特性的動態可逆過程,在胚胎發育、傷口愈合、癌癥轉移和纖維化等過程中發揮了重要作用。EMT 的主要特征有細胞黏附分子(如E-cadherin)表達的減少、細胞角蛋白細胞骨架轉化為Vimentin 為主的細胞骨架及形態上具有間充質細胞的特征。在EMT過程中,細胞將失去其黏附和極性特征,上皮細胞的標志物E-cadherin 的表達水平逐漸降低,而間充質細胞的標志物Vimentin 和N-cadherin 的表達水平會逐漸升高,而且N-cadherin 可以通過其鋅指結構與E-cadherin 啟動子區域的E-box元件結合而直接抑制E-cadherin的表達,從而啟動EMT 過程。然而,NCAPH 是否參與子宮內膜癌細胞的EMT 過程尚未見報道。本研究通過Western blot 檢測了EMT 相關基因的表達水平,結果發現,沉默NCAPH上調了子宮內膜癌細胞中E-cadherin 的表達水平,同時下調了Vimentin 和N-cadherin 的表達水平。這些結果表明,沉默NCAPH抑制了子宮內膜癌的EMT過程。
綜上所述,NCAPH 在子宮內膜癌組織和細胞中異常過表達,沉默NCAPH 能夠抑制子宮內膜癌細胞的增殖、遷移、侵襲和EMT。因此,NCAPH 有希望成為診斷子宮內膜癌患者的有效生物標志,并可能作為治療子宮內膜癌的潛在基因靶點,但是其中的分子機制還需要進一步的研究和證明。
猜你喜歡
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
火花(2019年12期)2019-12-26 01:00:28
人大建設(2019年6期)2019-10-08 08:55:48
人大建設(2019年12期)2019-05-21 02:55:32
雜文月刊(2018年21期)2019-01-05 05:55:28
人大建設(2017年6期)2017-09-26 11:50:44
學苑創造·A版(2015年11期)2016-01-14 09:03:27
俄羅斯問題研究(2012年1期)2012-03-25 09:54:45
中國火炬(2010年12期)2010-07-25 13:26:22
中國火炬(2010年8期)2010-07-25 11:34:30
