基于SSR 分子標記的自然狀態下蕨麻采樣策略研究
2023-09-25 03:42:54田甜李軍喬馬斌王鑫慈曲俊儒
草業學報 2023年9期
田甜,李軍喬,馬斌,王鑫慈,曲俊儒
(1. 青海民族大學生態環境與資源學院,青海 西寧 810007;2. 青海省特色經濟植物高值化利用重點實驗室,青海 西寧 810007;3. 青藏高原蕨麻產業研究院,青海 西寧 810007;4. 青海省湟源縣種子站,青海 西寧 811699)
蕨麻(Potentilla anserina)屬薔薇科(Rosaceae)委陵菜屬,是鵝絨委陵菜的變種。鵝絨委陵菜在世界及中國分布廣泛,只在青藏高原等高寒地區根系才膨大為塊根,稱為“蕨麻”。蕨麻塊根為圓球狀、紡錘狀或棒狀,可食用或藥用,俗稱“人參果”“延壽果”,《晶珠本草》《西寧府新志》《中國藏藥志》《四部醫典》中均有記載[1-5]。據本課題組測定,塊根富含淀粉、蛋白質、氨基酸、礦質元素、多糖、皂苷、總黃酮和鞣質等營養及活性成分,質糯,味甜,口感佳,具有抗缺氧、增強免疫力、保肝護肝、干擾病毒復制、抑制腫瘤細胞生長等功效[6-10]。民間將其作為食品及藏藥已經使用了上千年,市場價值有增無減。農牧民將采挖野生蕨麻作為主要的經濟來源,導致當地生態環境的惡化,野生蕨麻儲藏量和品質逐年下降。為了解決生態、生產和發展的矛盾,本課題組對蕨麻進行了人工馴化、品種審定,通過20 年的研究,蕨麻種植已成為青海省和甘肅省的一個產業。
蕨麻系典型的克隆植物,具有豐富的匍匐莖,人工栽培條件下以無性繁殖為主。無性繁殖時,由塊根長出基株,基株向四周生長出10~20 條一級匍匐莖,紫紅色或綠色,生長迅速,匍匐莖每隔3~5 cm 生長出一個分株,分株處又生長出2、3 級匍匐莖。一株蕨麻一年生長量可覆蓋3~5 m2土地。每個分株下的根系均可膨大,來年每個塊根均可作為基株繼續繁殖,如此周而復始,覆蓋面積逐年增大,野外條件下具體繁殖面積不明。蕨麻還具有完整的花器,花冠5 枚,雄蕊和雌蕊20 枚以上,可產生生物學種子。人工條件下花朵種子結實率不足10%,種子萌發率約為2%,難以成活。課題組前期試驗結果表明,蕨麻有性和無性繁殖并存,人工栽培時,蕨麻繁殖以無性為主,自然條件下,無性和有性繁殖比率相當,有性繁殖占比高于人工栽培,花粉傳播主要依靠昆蟲[11],傳播距離不明。由于野外條件下蕨麻生長了幾千年甚至上萬年,每年蕨麻都以無性繁殖及有性繁殖方式進行繁殖,個體遺傳距離不明確,更無法確定野外狀況下的采樣距離,導致采樣時居群內個體間和居群間的距離無法確定,對采樣及遺傳分析造成了極大的困擾。
目前對于蕨麻有性繁育和遺傳學方面的研究較少,課題組通過對蕨麻簡單重復序列(simple sequence repeat, SSR)研究發現,地理距離較遠的兩個樣本在親緣關系上可以非常近,聚為一支[9,12],這與蕨麻是兼性克隆植物有關[13]。SSR 多用于研究物種遺傳多樣性、指紋圖譜、遺傳結構等,馬斌等[14]利用SSR 分子標記構建了蕨麻3 個品種的指紋圖譜,魏姍姍等[15]用SSR 分子標記技術分析了95 份桃(Prunus persica)品種的遺傳多樣性。課題組使用磁珠富集法開發的SSR 引物,為蕨麻基因組學研究、育種進程提供了新的分子標記技術[12]。
野外采樣時,如何有效避免采集的樣品來自同一親本,主要考慮因素為蕨麻無性繁殖距離為多遠,是否存在有性繁殖,有性繁殖花粉能傳播多遠等。國內外關于蕨麻的研究主要側重于克隆習性、化學成分、藥理活性等方面,蕨麻栽培技術、生物學特性、有性繁殖、分子標記等方面的研究均為本課題組發表的論文,關于蕨麻采樣策略的分子生物學研究未見報道[16-24]。蕨麻無性繁殖的距離在人工種植情況下從形態上難以區分蕨麻的不同親本,而蕨麻種子結實率低,萌發率低,雜交不親和,常規方法無法完成蕨麻有性繁殖的研究。為明確蕨麻個體和居群間的采樣距離,本研究利用SSR 分子標記技術研究野生蕨麻的遺傳相似性,確定自然狀態下野生蕨麻的采樣策略和距離,為野生蕨麻樣品采集、選育和馴化等方面提供理論依據。
1 材料與方法
1.1 研究區域概況與試驗材料
基于蕨麻在西北高原地區的分布面積,試驗分別選取青海省、甘肅省、四川省和西藏自治區為主要地區,分別在不同地區選取不同的居群數量進行試驗。2017 年5-7 月,采樣路線及地點結合蕨麻分布面積和青藏高原山脈分布選定。甘肅省和四川省蕨麻分布面積小,各設置1 個居群,甘肅省為碌曲縣(原點:34°48′54″E,102°43′38″N),四川省為若爾蓋縣(原點:33°59′52″ E,102°46′14″ N);青海省和西藏自治區蕨麻分布面積大,各設置2 個居群,青海省為河南縣(原點:34°47′21″ E,101°33′06″ N)和祁連縣(原點:37°38′35″ E,101°25′31″ N),西藏自治區為八宿縣(原點:31°20′51″ E,97°36′27″ N)和南木林縣(原點:29°33′59″ E,89°04′32″ N)。樣品采集共6 個居群。采樣方法為選取一個原點,沿直線采樣。其中,由于青藏高原海拔高、地形復雜,山脈眾多,蕨麻居群采樣并不完全是直線(采樣分布見圖4,圖6,圖8,圖10,圖12 和圖14)。
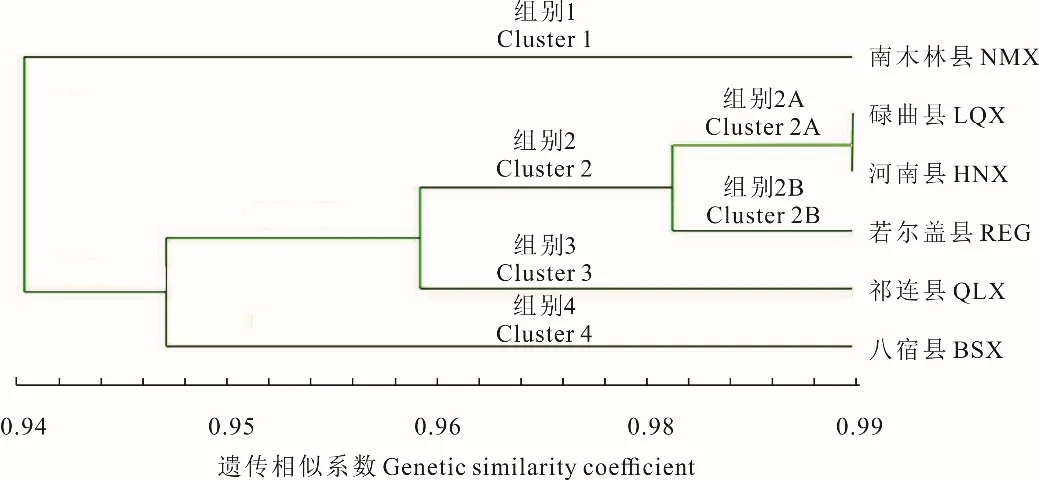
圖1 6 個蕨麻居群的UPGMA 聚類樹Fig.1 UPMGA cluster trees for six P. anserina populations
采樣時,以第一個采樣點為原點,后分別在距離原點1、2、3、4、5、10、15、20、25、30、40、50、60、80、90、100、120、150、160、200、250、300、400、500、1000、1500、2000、2500、5000、10000、15000、20000、30000、40000、50000 m處采樣,1000 m 以內的距離用測輪測量,1000 m 以上的距離用汽車測量。采集新鮮、無病斑、幼嫩的蕨麻葉片,放入硅膠袋中,備用。調查、記錄每一個樣本的采集地海拔和經緯度等信息。采樣點基本信息見表1(伴生植物所列物種在所有采樣點中均有)。
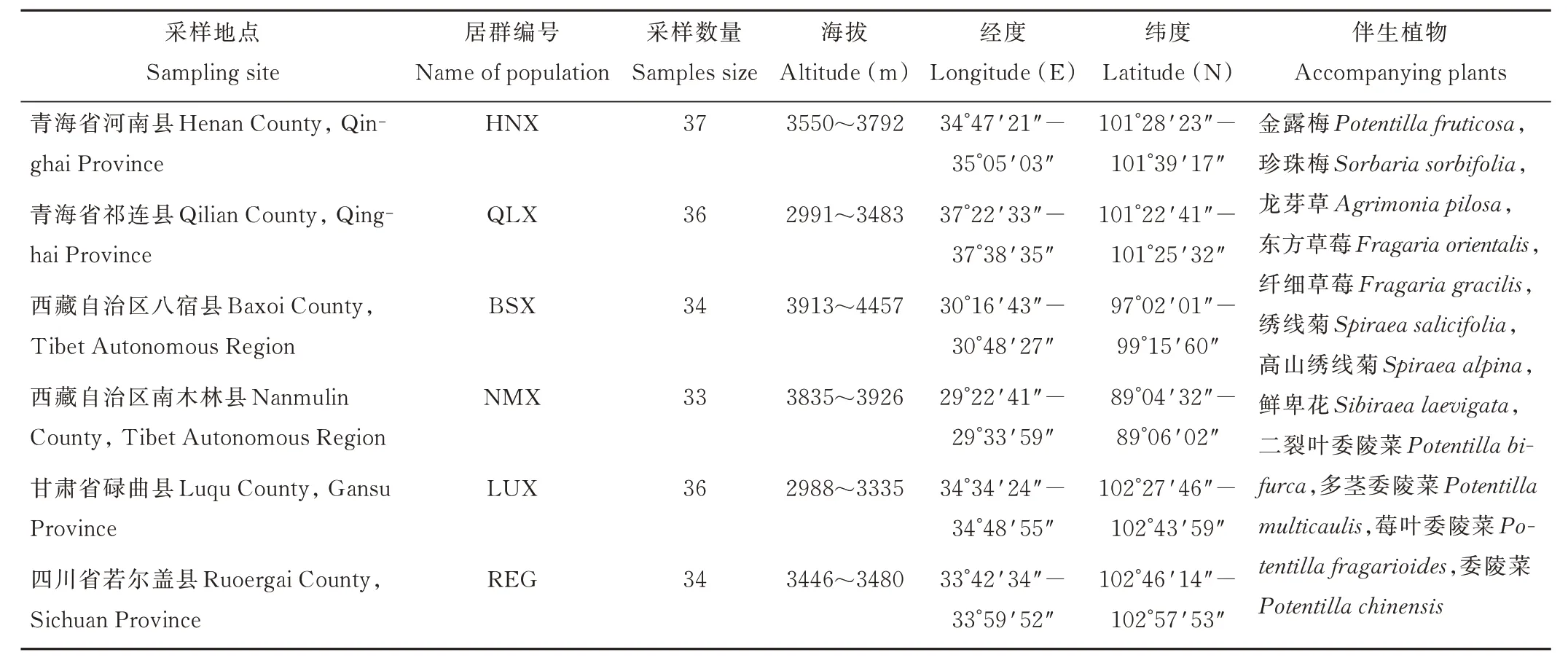
表1 采樣點基本信息Table 1 Basic information of sampling points
1.2 試劑與儀器
1.2.1 DNA 提取試劑 1 mol·L-1Tris-HCL (pH 8.0)、0.5 mol·L-1EDTA(pH 8.0)、5 mol·L-1NaCl、2×CTAB 緩沖溶液、氯仿/異戊醇(24∶1)、異丙醇、75%乙醇、無水乙醇、50×TAE 緩存溶液、4S Red Plus 核酸染色劑(10000×水溶液)、6×Loading buffer 上樣緩沖液,試劑均為分析純。
1.2.2 PCR 反應試劑 Taq 酶、dNTP、10×PCR Buffer,由生工生物工程(上海)股份有限公司提供。
1.2.3 試驗儀器 GR60DP 高壓蒸汽滅菌鍋(ZEAL WAY INSTRUMENT INC)、Neofuge 23R 高速臺式冷凍離心機(香港)、Scientz-192 高通量組織研磨儀(寧波)、MiniBIS pro 凝膠成像系統(以色列)、Mastercycle pro PCR 儀(德國)、3730XL3730XL 測序儀(美國)。
1.3 試驗方法
1.3.1 DNA 提取及檢測 本試驗采用試劑盒改良CTAB 法[25-26]提取6 個居群210 份樣本的DNA。經1%瓊脂糖凝膠檢測DNA 濃度,用滅菌的去離子水稀釋濃度至50 ng·μL-1,保存于4 ℃冰箱中備用。
1.3.2 引物信息與SSR 標記擴增 試驗所用引物為課題組已篩選出的具有高度多態性的SSR 引物[9,12],具體信息見表2。
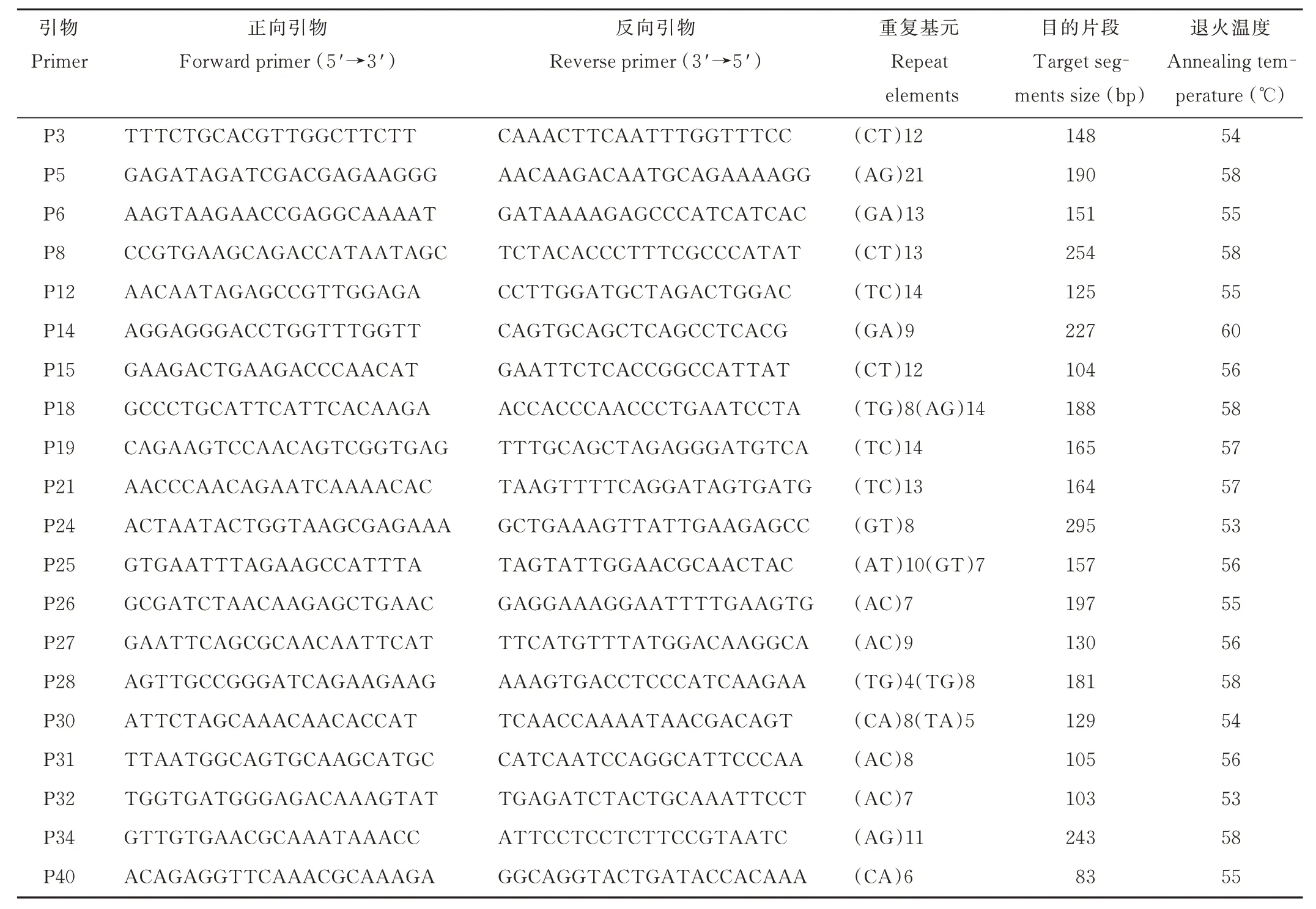
表2 SSR 引物序列信息Table 2 The information of SSR primer sequence
用篩選出的SSR 引物對210 個蕨麻個體材料進行SSR 擴增。PCR 擴增體系總體積為20 μL:Taq(5 U·μL-1)0.14 μL、dNTP(15 mmol·L-1)1.6 μL、Buffer(10×) 2.0 μL、正向引物(20 μmol·L-1)0.4 μL、反向引物(20 μmol·L-1) 0.4 μL、DNA(50 ng·μL-1)2.5 μL、ddH2O 12.96 μL。PCR 反應條件:94 ℃預變性5 min,94 ℃變性30 s,54 ℃(退火溫度以優化的為標準)復性35 s,72 ℃延伸40 s,共35 個循環,最終72 ℃延伸3 min。PCR 產物保存備用。
1.3.3 毛細管電泳 將甲酰胺與分子量內標按體積比100∶1 混合均勻,在測序儀上樣板中加入9 μL,再加入1 μL 稀釋10 倍的PCR 產物[26]。毛細管電泳使用3730XL 測序儀進行,利用GeneMarker 中的Fragment(Plant)片段分析軟件對得到的原始數據進行分析。比較各泳道內分子量內標的位置與各樣品峰值的位置,分析片段大小[27]。
1.4 數據分析
利用GeneMarker 軟件的等位基因峰圖對210 份材料、20 對引物組合擴增得到的4200 個原始數據進行分析,根據擴增有條帶和無條帶統計數據,修正后(有條帶為擴增陽性,記為1;相應位置無條帶為擴增陰性,記為0)轉為0/1 的數列格式,建立原始數列矩陣,利用POPGENE 32[28]、NTSYS-pc 2.10e[29]和GenAIEx 6.5[30]等軟件進行后續分析。
利用POPGENE 32 軟件對6 個居群210 份樣本的原始(0,1)數值矩陣進行遺傳多樣性分析,并計算多態性指數(polymorphic information content, PIC)[31],計算公式為:
式中:Pij表示標記i的第j個等位基因在群體中的分布頻率;標記i的總等位基因為1~n[32],n是等位基因數目。
利用NTSYS-pc 2.10e 軟件對6 個居群210 份樣本的原始(0,1)數值矩陣進行處理,運用clustering 程序中的SHAN 程序進行非加權組平均法(unweighted pair-group method with arithmetic means, UPGMA)聚類分析,繪制親緣關系樹狀圖。
運用GenAlEx 6.5 軟件,依據居群遺傳距離對6 個居群作主成分分析,同時進行空間自相關分析,以揭示空間距離和遺傳距離之間的關系。
2 結果與分析
2.1 SSR 擴增結果
提取的DNA 質量為500 ng·μL-1以上,純度為1.8,達到SSR-PCR 擴增條件。利用20 對核心引物對蕨麻210 份材料進行PCR 擴增,然后通過毛細管電泳檢測分析。20 對引物一共擴增出93 個多態性片段(表3),擴增片段大小為78~306 bp。擴增片段最小引物為P40,最大引物是P24。在20 對SSR 位點共檢測出93 個等位基因,每對引物檢測出2~7 個等位基因,平均為4.65 個。20 個位點的多態性指數(PIC)變化為0.25~0.82,平均為0.61;SSR 引物P3 和P19 獲得的等位基因數最多(7 個),PIC 值分別為0.82 和0.80;引物P34 獲得的等位基因數最少(2個),PIC 值為0.25。PIC 值可作為判斷SSR 引物多態性高低的指標[31],本試驗中90%以上的引物PIC>0.5,表明試驗中所篩選的引物具有高度多態性,擴增結果可反映蕨麻資源的遺傳多樣性。
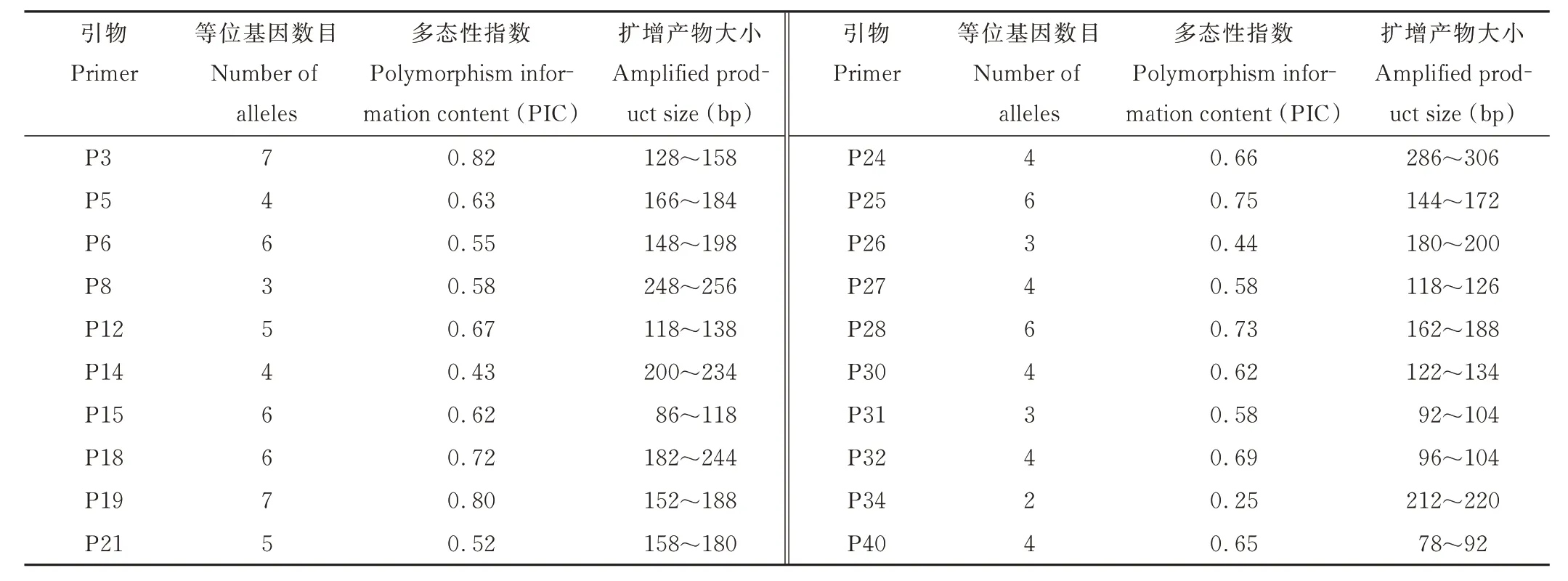
表3 20 對核心引物多態性指數Table 3 The polymorphic information content of 20 SSR primers
2.2 遺傳多樣性分析
利用POPGENE 32 軟件分析得到蕨麻6 個居群的遺傳參數(表4),祁連縣居群(QLX)的遺傳多樣性最高[平均雜合度(average heterozygosity,H)=0.2797,香農指數(Shannon index,I)=0.4287]。河南縣居群(HNX)的遺傳多樣性最低(H=0.2273,I=0.3542)。6 個居群遺傳多樣性由高到低依次為QLX>LQX>BSX>NMX>REG>HNX。
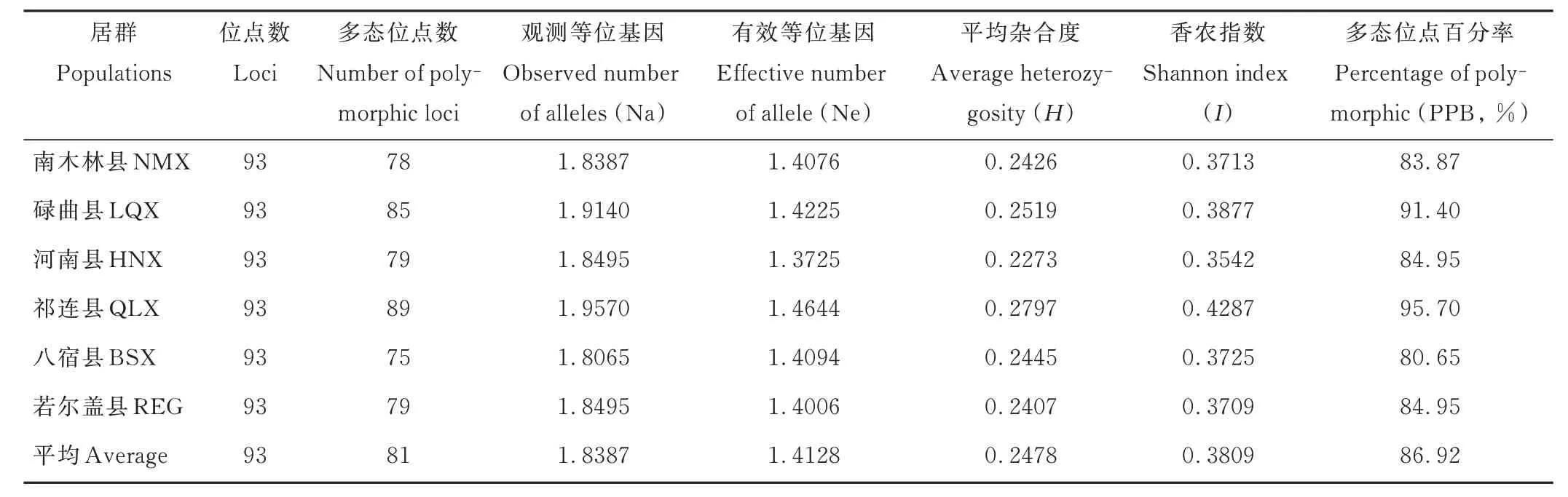
表4 6 個蕨麻居群的遺傳多樣性Table 4 Genetic diversity of the six P. anserina populations
利用GenAIEx 6.5 對蕨麻6 個居群進行AMOVA 分析,結果顯示(表5),主要變異來源于居群內,占比84%,居群間占有16%的遺傳變異。

表5 蕨麻6 個群體分子方差分析Table 5 AMOVA analysis of six P. anserina populations
通過計算群體間的遺傳分化系數及基因流,河南縣居群和碌曲縣居群之間的基因流最大(Nm=0.986),說明兩個居群之間進行基因交流的頻率較多,從而使得遺傳分化指數較低(Fst=0.014),這與遺傳分析結果一致。八宿縣居群和楠木林縣居群之間的遺傳分化指數最大(Fst=0.084),基因流最低(Nm=0.919),說明兩個居群之間遺傳多樣性較低,大型山脈的阻隔可能為主要原因。同時,6 個居群之間的遺傳分化指數(Fst)都小于0.1(<0.15),說明整體的遺傳分化比較小。群體之間的基因流在0.900 以上,表示群體間遺傳差異較小(表6)。
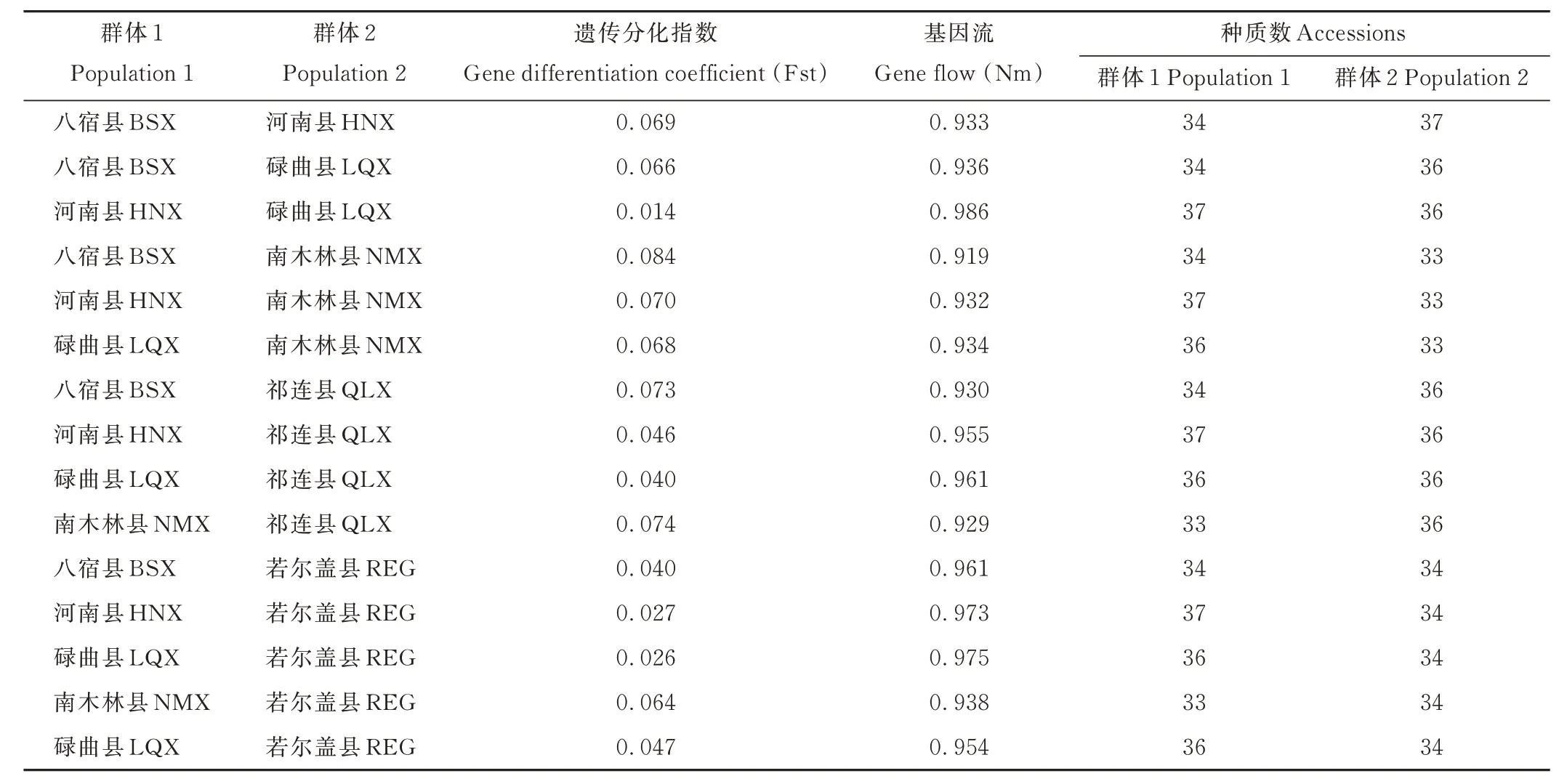
表6 蕨麻群體間的遺傳分化系數和基因流Table 6 The Fst and Nm among P. anserina populations
蕨麻6 個居群的遺傳相似度和遺傳距離顯示(表7),各居群間的遺傳相似度高,為0.9194~0.9860,遺傳距離也較近,為0.0141~0.0840。其中,碌曲縣居群和河南縣居群的遺傳距離最小,遺傳一致度最大;南木林縣居群和八宿縣居群遺傳距離最大,遺傳一致度最小。
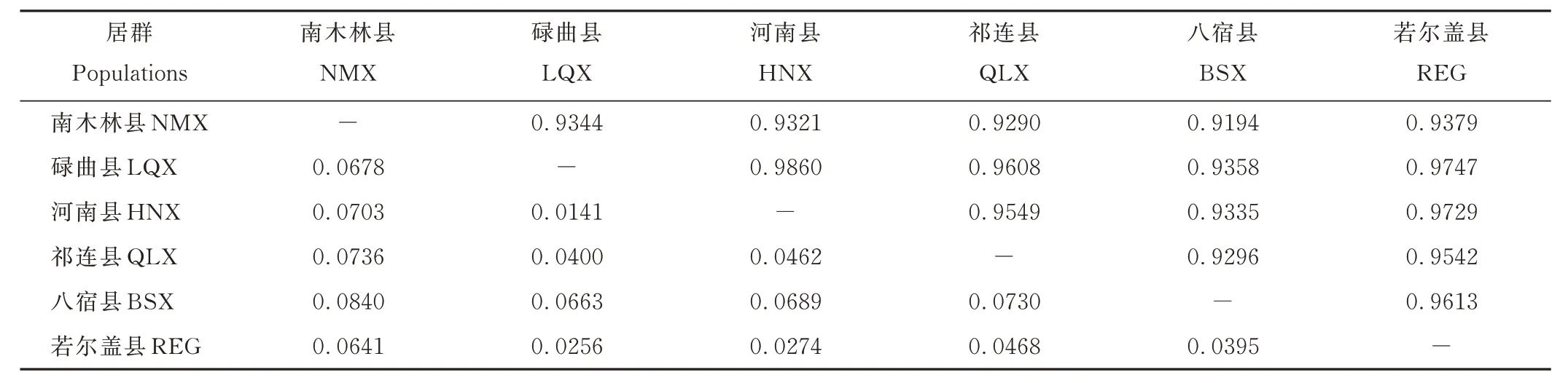
表7 蕨麻各居群之間的遺傳相似度與遺傳距離Table 7 Genetic identity and genetic distance of six P. anserina populations
基于POPGENE 32 分析結果,在歐式聚類平均法(UPGMA)計算中得出,當遺傳相似系數為0.98 時,6 個居群分為4 組(圖1),南木林縣(NMX)、祁連縣(QLX)以及八宿縣(BSX)各自單獨聚類為一組,碌曲縣(LQX)、河南縣(HNX)和若爾蓋縣(REG)聚為一組(Cluster 2),各群體間距離依次增大,親緣關系也逐漸變遠。其中,Cluster 2 包括兩個亞組,Cluster 2A 包括2 個群體,為碌曲縣(LQX)和河南縣(HNX),Cluster 2B 包括1 個群體,為若爾蓋縣(REG)。
利用GenAIEx 6.5 對6 個蕨麻群體的210 個樣本進行主成分分析(圖2),結果與聚類分析結果一致,碌曲縣、河南縣以及若爾蓋縣按照居群聚為一簇,這與其地理位置有關,中間沒有橫亙山脈,為其進行基因交流提供了便利;而遺傳多樣性較高的居群,如祁連縣、南木林縣和八宿縣,由于居群內個體遺傳背景差異較大,聚類較分散,均單獨聚為一類。
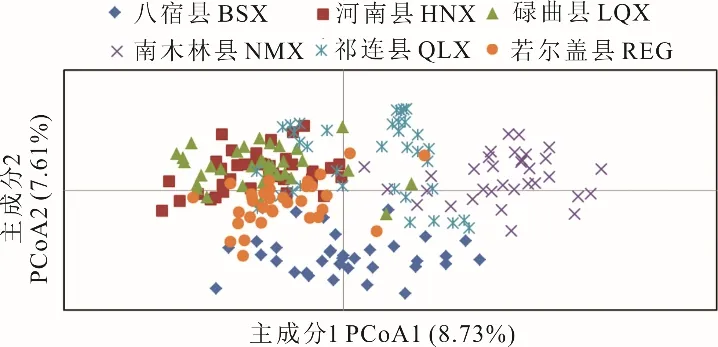
圖2 210 個蕨麻個體的主成分分析Fig. 2 PCoA analysis of 210 individuals in P. anserina populations
南木林縣、祁連縣以及八宿縣分布較為分散,其間的大型山脈阻隔是主要原因。其中,南木林縣和八宿縣的個體最為分散,沒有規律性,這可能與地理距離有關。
2.3 遺傳采樣策略
依據20 對引物對每個地區蕨麻樣品的擴增點位,構建(0,1)的數列矩陣,利用NYSYS 2.1 對蕨麻6 個居群進行分析,對每個居群內的蕨麻樣品進行UPGMA 聚類,并對各居群遺傳距離矩陣和地理距離矩陣進行相關性計算(Mantel 檢驗)。
根據碌曲縣蕨麻樣品UPGMA 聚類(圖3)可知,該地區蕨麻個體間遺傳距離較近,遺傳距離最大為0.58。根據聚類分析及采樣點分布(圖3 和圖4),該地區蕨麻個體間遺傳距離隨地理距離的增大呈逐步增加的態勢,且大概30 號以前的樣品間遺傳距離較近,而30 號以后的樣品與30 號以前的樣品間遺傳距離逐步增大。以30 號樣品為分界線,分別為5 km 以內的個體與5 km 以上的個體,表明在一定范圍內為同一克隆的可能性更大。遺傳距離和地理距離相關性顯示(圖4),R2=0.0086,P=0.13>0.05,兩者之間沒有顯著的聯系。表示聚類并不完全按照地理距離進行,蕨麻個體與地理距離相關性較差,這與采樣個體分布于盤山地區,沒有較強的地理隔離有關。
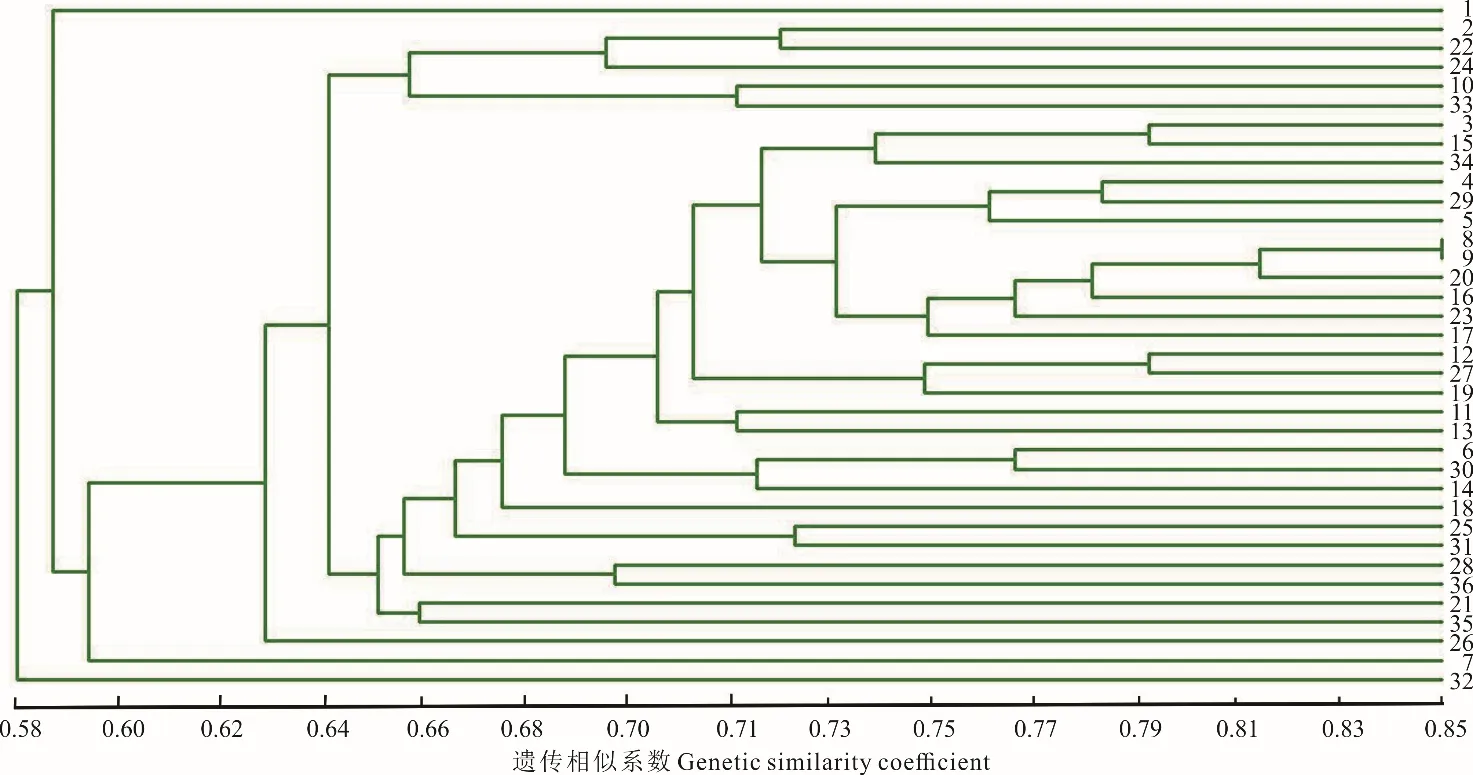
圖3 甘肅省碌曲縣蕨麻樣品UPGMA 聚類分析Fig.3 The UPGMA cluster analysis of P. anserina samples in Luqu County, Gansu Province
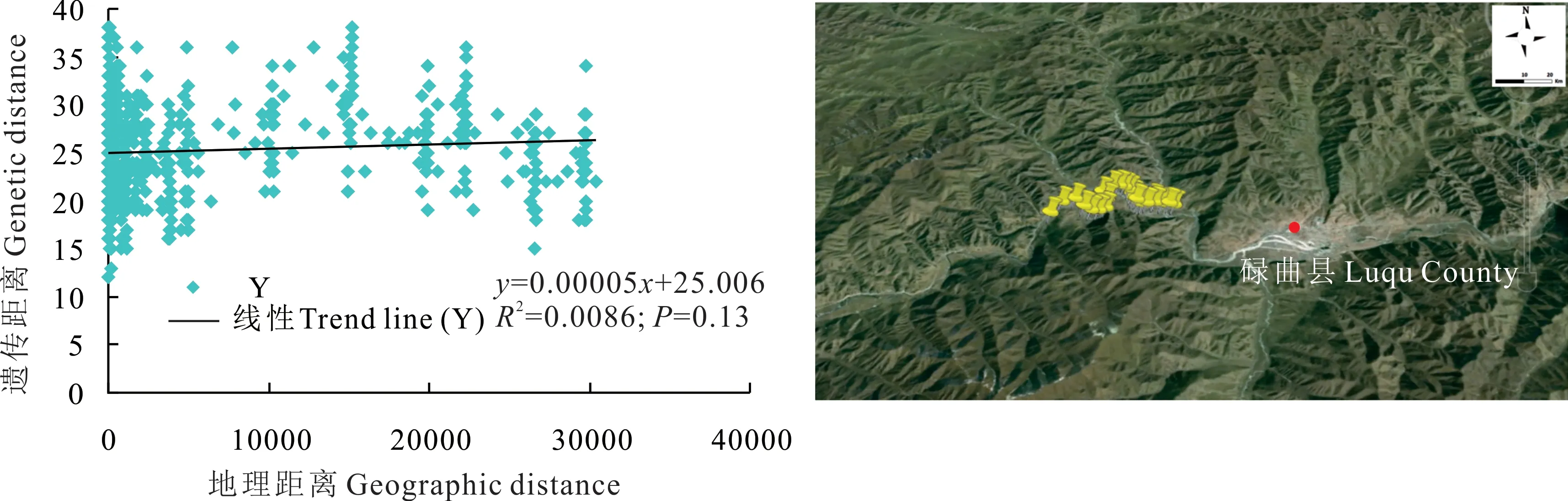
圖4 甘肅省碌曲縣蕨麻遺傳距離與地理距離相關性分析及采樣路線Fig. 4 Sample path and mantle test of relationship between genetic distance and geographic distance of P. anserina in Luqu County, Gansu Province
根據若爾蓋縣(REG)蕨麻樣品UPGMA 聚類(圖5)得出,群體內遺傳距離較近,遺傳距離最大為0.60。根據聚類分析及采樣點分布(圖5 和圖6)得出,該地區蕨麻個體采樣點均分布于較為平坦的山丘地區,遺傳距離和地理距離相關性顯示(圖6),R2=0.0248,P=0.06>0.05,兩者之間沒有顯著的聯系,表明該地區的蕨麻個體間遺傳距離與地理距離相關性較差,且蕨麻個體間遺傳距離較為復雜。該居群的平均雜合度(H)、香農指數(I)高達0.2407 和0.3709(表4),表示其遺傳多樣性較高,這與該地區地勢較為平坦,且多為草原地帶,蕨麻個體間無明顯的地理隔離,導致基因交流較頻繁有關。
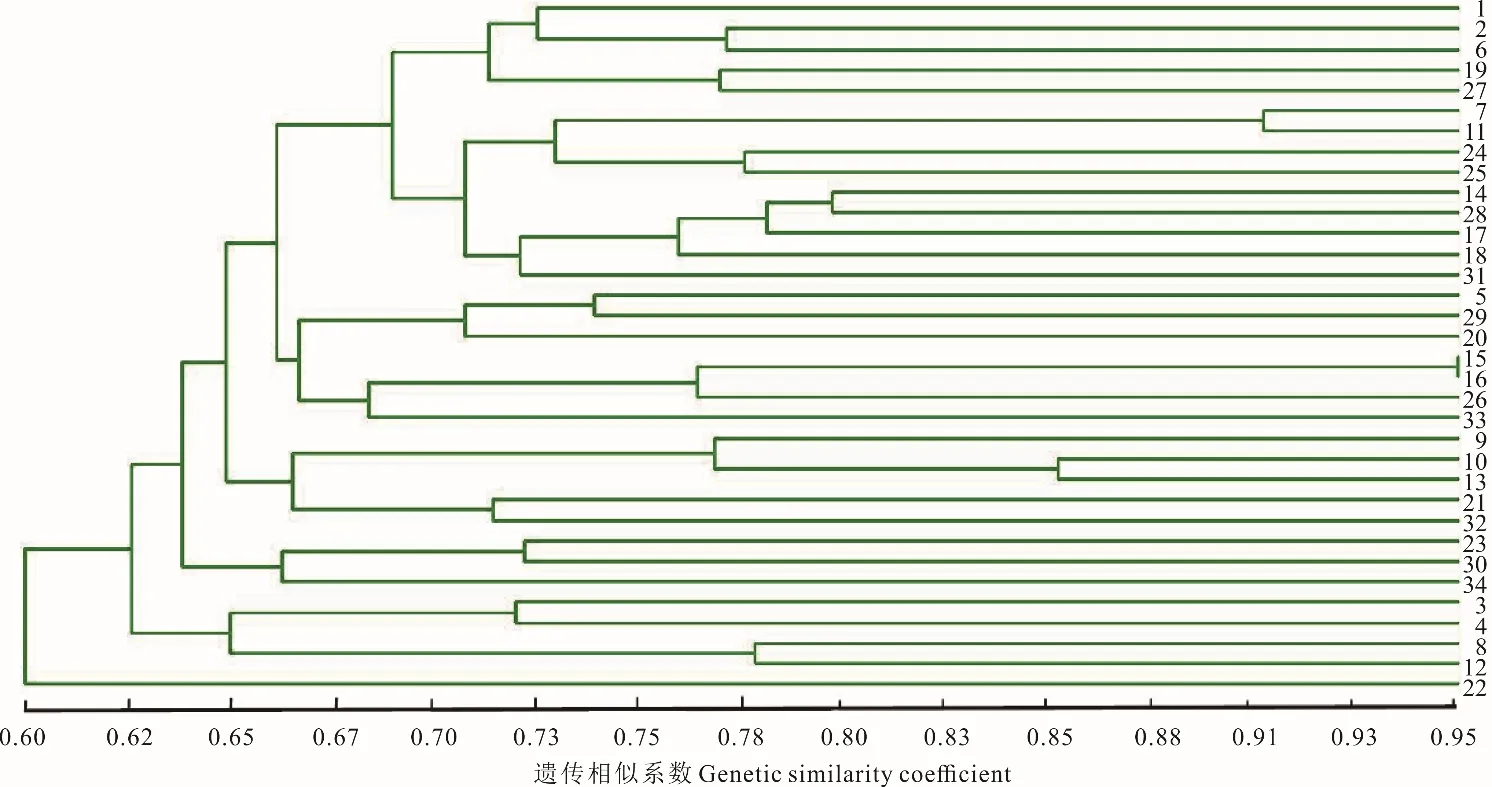
圖5 四川省若爾蓋縣蕨麻樣品UPGMA 聚類分析Fig.5 The UPGMA cluster analysis of P. anserina samples in Ruoergai County, Sichuan Province
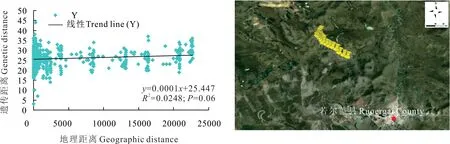
圖6 四川省若爾蓋縣蕨麻遺傳距離與地理距離相關性分析及采樣路線Fig. 6 Sample path and mantle test of relationship between genetic distance and geographic distance of P. anserina in Ruoergai County, Sichuan Province
根據河南縣蕨麻樣品UPGMA 聚類(圖7)可知,該地區蕨麻遺傳距離較近,遺傳距離最大為0.59。根據聚類分析及采樣點分布(圖7 和圖8),在遺傳距離為0.76 處,以30 號蕨麻樣品為分界線,蕨麻樣品被聚為兩類,分別為5 km 以內的蕨麻個體與5 km 以上的蕨麻個體。遺傳距離和地理距離相關性顯示(圖8),R2趨近于0,P=0.45>0.05,兩者之間沒有顯著的聯系,這與該地區蕨麻采樣點分布于較平坦的地區,沿直線排布有關,無明顯地理隔離導致蕨麻個體間基因交流較多。
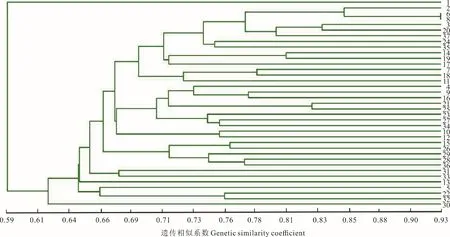
圖7 青海省河南縣蕨麻樣品UPGMA 聚類分析Fig.7 The UPGMA cluster analysis of P. anserina samples in Henan County, Qinghai Province
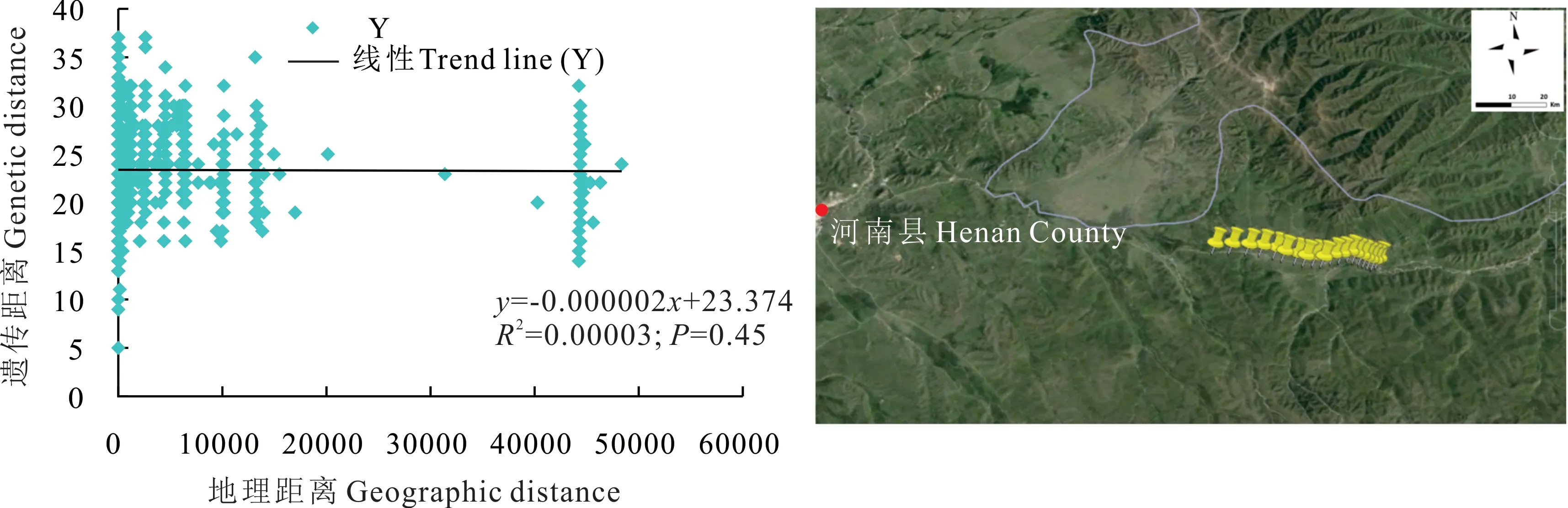
圖8 青海省河南縣蕨麻遺傳距離與地理距離相關性分析及采樣路線Fig. 8 Sample path and mantle test of relationship between genetic distance and geographic distance of P. anserina in Henan County, Qinghai Province
根據祁連縣蕨麻樣品UPGMA 聚類(圖9)可知,該地區蕨麻遺傳距離較近,遺傳距離最大為0.54。根據聚類分析及采樣點分布(圖9 和圖10)可知,1~29 號樣品被明顯聚為一類,這些樣品的采樣點均為5 km 以內,30~36號樣品被聚為一類,這些樣品采樣點距離均為5 km 以上。遺傳距離和地理距離相關性顯示(圖10),R2=0.1982,P=0.01<0.05,兩者之間有顯著的聯系,表明該地區蕨麻個體樣品間遺傳距離也隨著地理距離的增加而增大。該地區蕨麻采樣點分布較均勻,并按照一定的直線排布,蕨麻個體間遺傳距離與地理距離相關性較好。
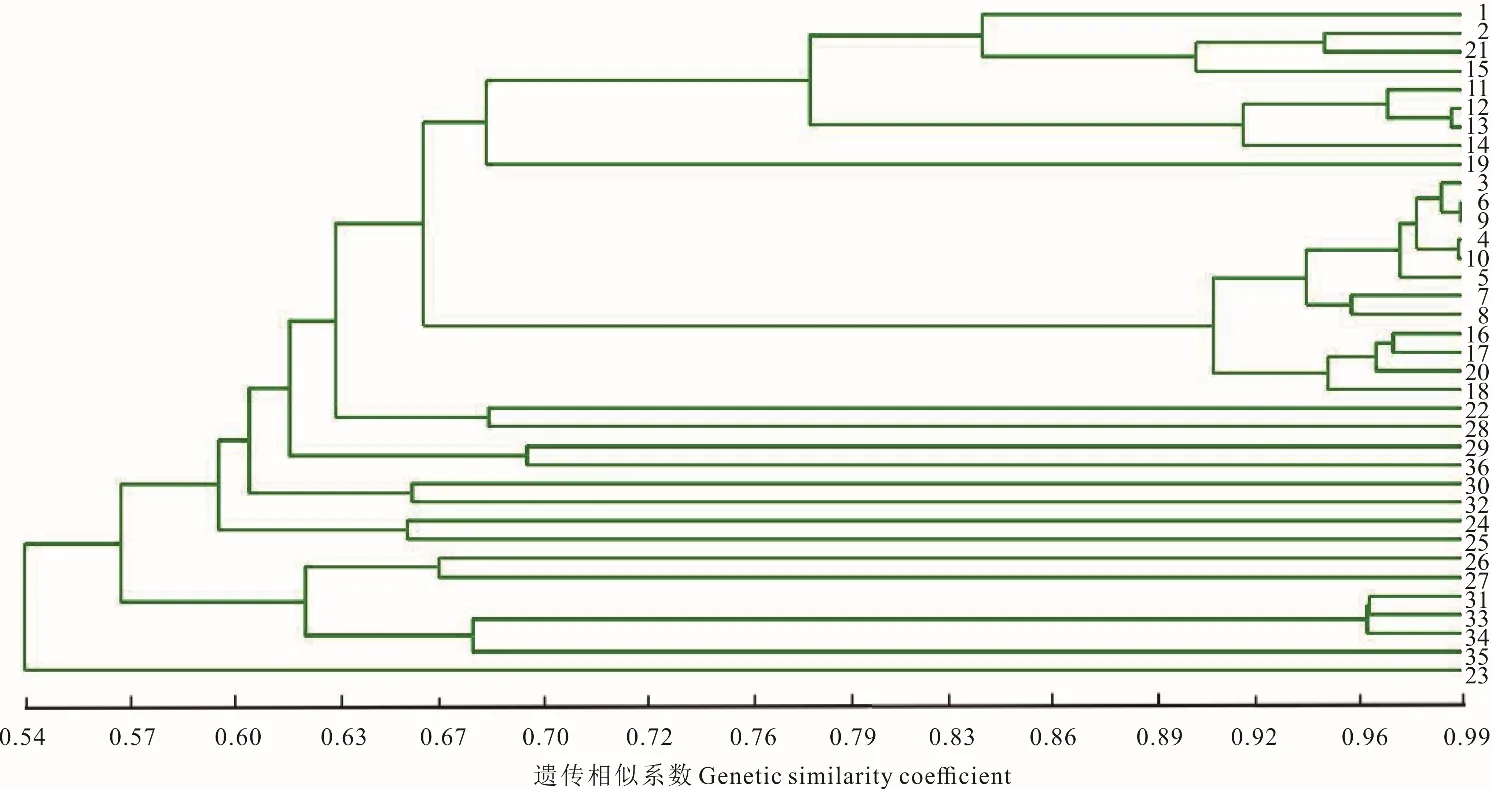
圖9 青海省祁連縣蕨麻樣品UPGMA 聚類分析Fig.9 The UPGMA cluster analysis of P. anserina samples in Qilian County, Qinghai Province
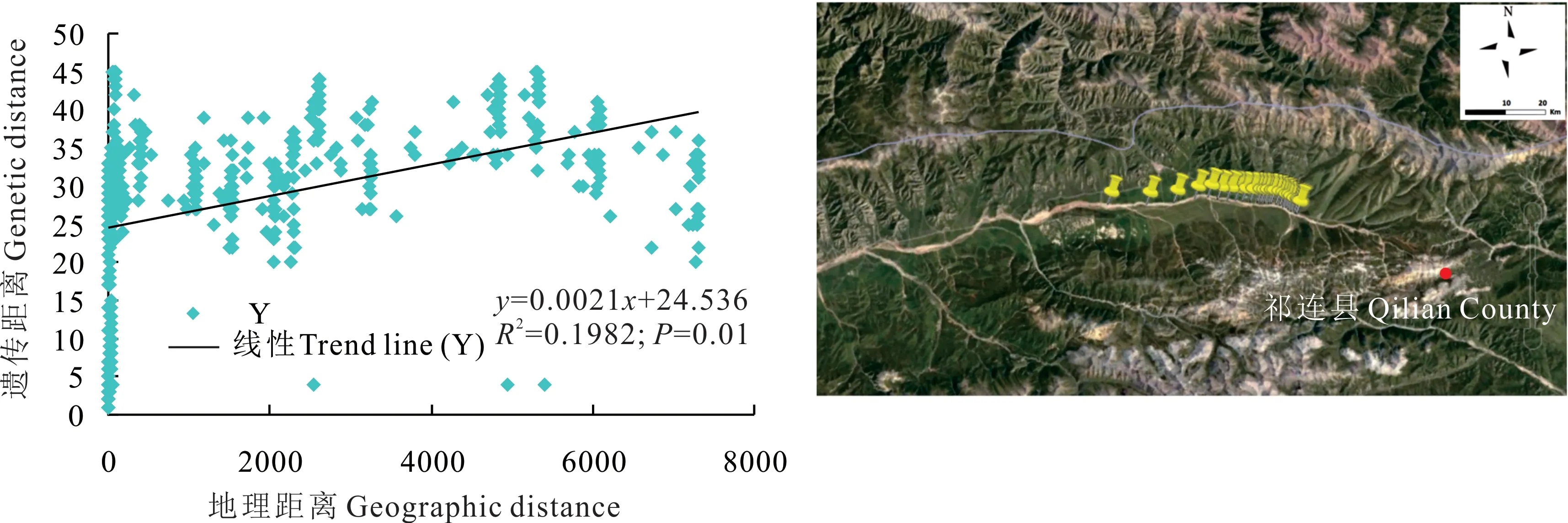
圖10 青海省祁連縣蕨麻遺傳距離與地理距離相關性分析及采樣路線Fig. 10 Sample path and mantle test of relationship between genetic distance and geographic distance of P. anserina in Qilian County, Qinghai Province
根據八宿縣蕨麻樣品UPGMA 聚類(圖11),蕨麻遺傳距離較近,遺傳距離最大為0.63。由聚類分析及采樣點分布(圖11 和圖12)可知,該地區蕨麻個體采樣點均分布于山谷中間,且地勢較復雜。遺傳距離和地理距離相關性顯示(圖12),R2=0.0118,P=0.16>0.05,兩者之間沒有顯著聯系。蕨麻個體間遺傳距離與地理距離相關性較差。由于西藏八宿縣位于橫斷山脈,地理情況較為復雜,且該地區為冰川期野生植物的避難所,遺傳多樣性較豐富,基因交流較為頻繁,但該地區部分蕨麻個體采樣點分布不是按照直線排布,所以出現蕨麻個體間遺傳距離與地理距離無明顯相關的現象。
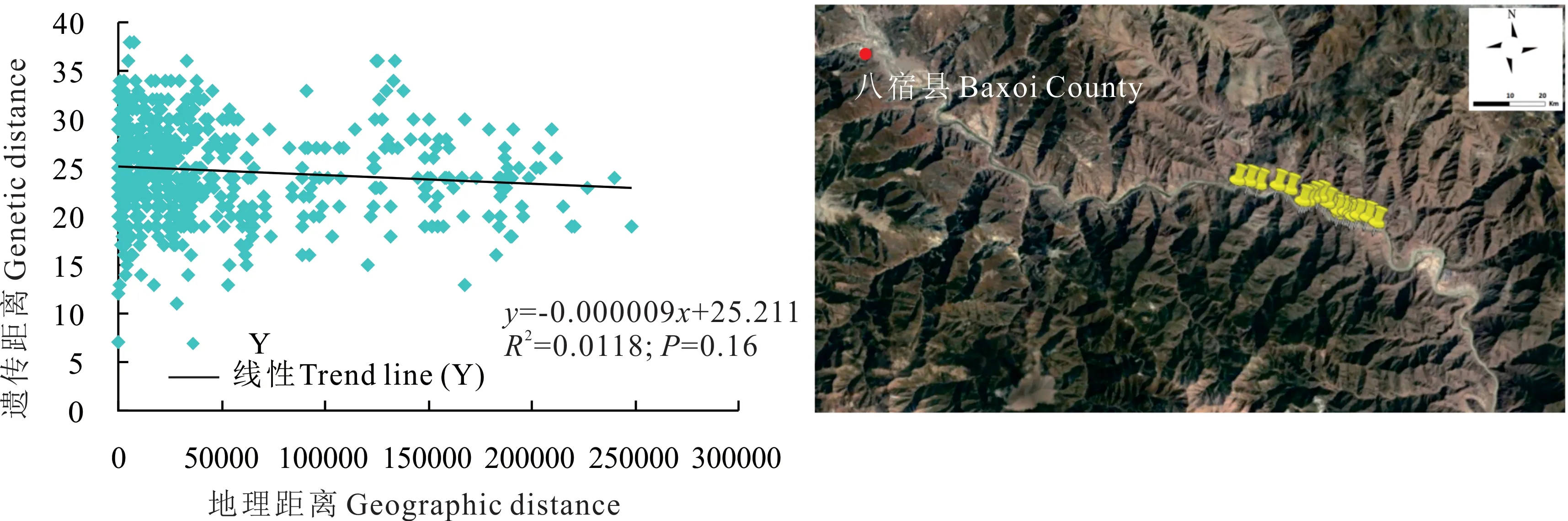
圖12 西藏自治區八宿縣蕨麻遺傳距離與地理距離相關性分析及采樣路線Fig. 12 Sample path and mantle test of relationship between genetic distance and geographic distance of P. anserina in Baxoi County, Tibet Autonomous Region
根據南木林縣蕨麻樣品UPGMA 聚類(圖13)可以得出,該地區蕨麻遺傳距離較近,遺傳距離最大為0.54。根據聚類分析及采樣點分布(圖13 和圖14)可知,該地區蕨麻個體間遺傳距離隨地理距離的增加而增大,具有一定的相關性。蕨麻個體間遺傳距離均較近,5 km 以上的蕨麻個體與5 km 以內的蕨麻個體遺傳距離較近,并沒有明顯的距離劃分。遺傳距離和地理距離相關性(圖14)顯示,R2=0.0204,P=0.13>0.05,兩者之間沒有顯著聯系。蕨麻個體間遺傳距離與地理距離相關性較差。這與該地區呈兩邊為高山,中間為狹長的河谷地區,蕨麻個體間基因交流并無明顯的阻隔有關。
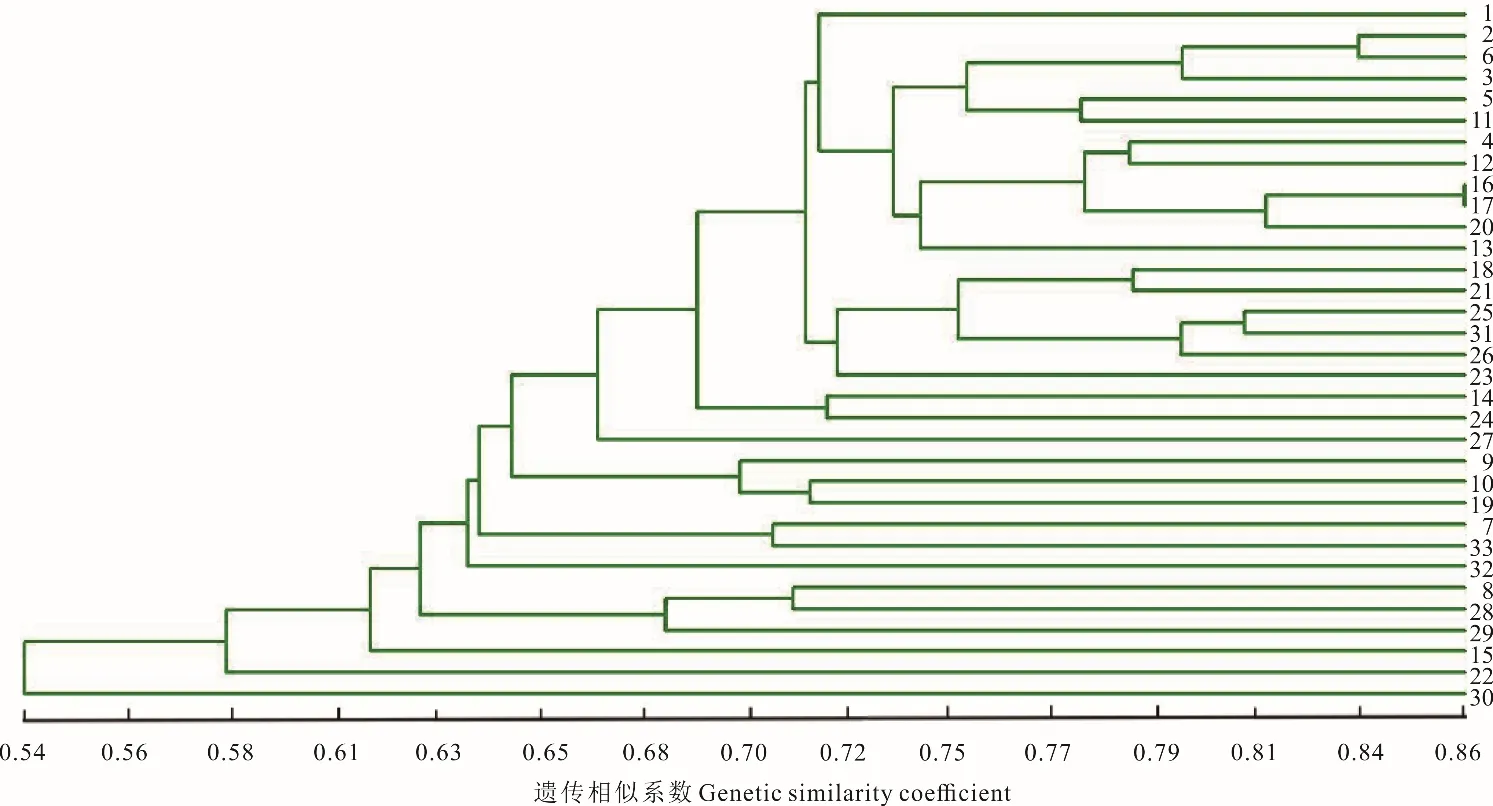
圖13 西藏自治區南木林縣蕨麻樣品UPGMA 聚類分析Fig.13 The UPGMA cluster analysis of P. anserina samples in Nanmulin County, Tibet Autonomous Region
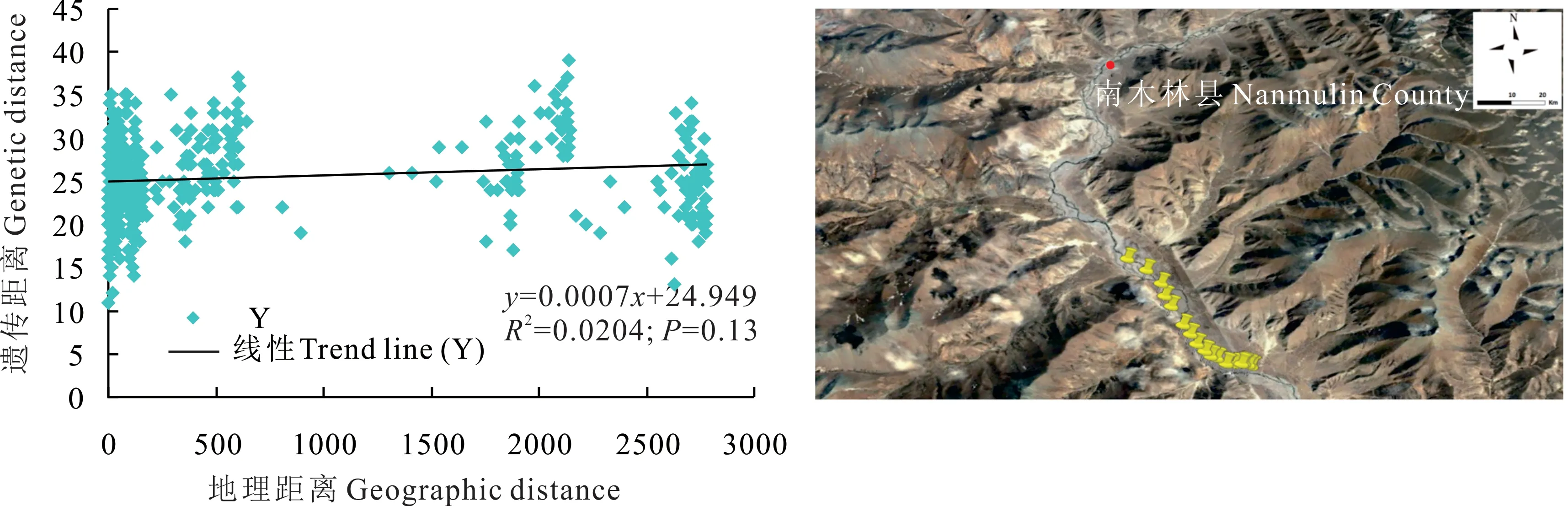
圖14 西藏自治區南木林縣蕨麻遺傳距離與地理距離相關性分析及采樣路線Fig. 14 Sample path and mantle test of relationship between genetic distance and geographic distance of P. anserina in Nanmulin County, Tibet Autonomous Region
3 討論
3.1 遺傳多樣性分析
本試驗使用20 對SSR 引物對蕨麻6 個居群210 個樣本進行PCR 擴增,共擴增出93 個多態性片段,平均PIC值為0.6145,根據Botstein 等[31]的劃分標準,20 對引物平均表現出高多態性,表明這些位點可用于蕨麻種質的遺傳多樣性分析。
多態位點百分率(percentage of polymorphic bands,PPB)在一定程度上可以反映物種的遺傳多樣性[33],兩者一般呈正相關,PPB 越大說明物種對環境適應得越好,容易擴展其分布范圍和開拓新的環境,遺傳多樣性也越豐富。本研究中各居群的PPB 均在80%以上,最高達91.40%(碌曲縣),說明蕨麻的遺傳多樣性豐富。蕨麻具有較高的遺傳多樣性,同時也具有中等水平的遺傳變異。一般認為,克隆植物遺傳多樣性都比較低[34],但在蕨麻中顯示出了不同的結果。越來越多的研究表明克隆植物的種群也可以保持很高的遺傳多樣性,兼性有性生殖可以有效補償遺傳變異的損失,在種群內提高遺傳變異,以此來消減種群間的遺傳變異;或者通過基因突變來提高遺傳變異,如三葉 草(Trifolium)、蒲公英(Taraxacum mongolicum)等[35]。劉春 香等[36]對 匍匐莖克 隆植物蛇 莓(Duchesnea indica)的研究發現,33 個野生蛇莓種群也具有很高的遺傳多樣性,遺傳距離與地理距離不存在顯著相關關系(r=0.052,P=0.150),這與蛇莓的無性繁殖以及對環境壓力的選擇有關。蕨麻遺傳多樣性高的原因可能為蕨麻是兼性克隆植物,種群內可以通過風媒、蟲媒在長距離范圍內進行花粉傳播,從而增加居群內的基因交流;蕨麻在高寒地區生長,周邊環境一般為山脈,阻礙了花粉和種子的傳播,且克隆生長受到限制,從而將居群之間分隔開來,形成生境隔離,各居群為適應不同環境而形成不同的進化;同時,采樣時各居群的地理環境不同,由于山脈、河流隔斷,大多數樣本不是直線距離采集,最終形成蕨麻的高遺傳多樣性。
蕨麻遺傳變異主要來源于居群內(居群變異方差和占比分別為12.745 和84%),居群間變異較低(居群變異方差和占比分別為2.477 和16%),這與杜夏瑾等[37]的研究結論一致。金燕等[38]認為,一個物種的分布區域大小可以直接影響其遺傳多樣性的高低,兩者成正比,分布區域廣的物種會由于寬闊的生境之間的差異而進化,從而提高遺傳變異。蕨麻本身為克隆植物,單個居群占地面積也較廣,在一個區域內,覆蓋率可達100%,在自然生長的居群中,充分發揮繁殖優勢,分布范圍廣,但由于是在高寒地區,生境差異大,為適應環境而變異,這也是自然選擇的結果。青藏高原山脈平均海拔4000~6000 m,距離地面高度平均為1000~1500 m(唐古拉山脈高6800 m,距離地面高度1800 m,昆侖山脈高5500~6000 m,距離地面高度1200~1700 m,祁連山脈高5800 m,距離地面1000~2000 m),各地區環境、氣候差異大。本研究發現,各居群間、居群內均有大小不同的山脈作為阻斷,且采樣時由于地勢的不一致,所采集的樣品并不能按照直線進行采集,從而使得群體的遺傳多樣性較高,居群內遺傳背景變大。
采樣區域內從南到北分布的大型山脈有喜馬拉雅山脈、橫斷山脈、昆侖山脈和祁連山脈,唐古拉山脈和巴顏喀拉山脈為小型山脈。南木林縣居群與其他5 個居群遺傳距離最遠,該居群位于拉薩以西,周邊環繞著橫斷山脈、唐古拉山脈、巴顏喀拉山脈等數條大型山脈,山脈阻隔最顯著,地理距離最遠,遺傳距離也最遠;八宿縣位于昌都以南,周邊環繞橫斷山脈,使得該區蕨麻與其他地區具有天然的生殖隔離屏障,地形最為復雜,遺傳多樣性豐富,該區和其他地區地理隔離也很明顯;祁連縣與上述兩個地區地理距離較遠,且中間橫亙昆侖山脈和祁連山脈,遺傳距離較遠,但由于其采樣時分布較均勻,按照直線進行采樣,其遺傳多樣性較為豐富,遺傳距離和地理距離呈顯著相關;若爾蓋縣位于昆侖山脈和橫斷山脈之間,山脈阻隔,遺傳距離和地理距離具有一定相關性;碌曲縣居群和河南縣居群位于昆侖山脈和祁連山脈中間,沒有山脈阻隔,遺傳距離最近,遺傳一致度最高(遺傳相似系數=0.986,遺傳距離=0.0141),且聚類為一簇。
3.2 遺傳采樣策略
克隆植物的遺傳距離對植物種質資源收集具有很重要的作用,沈曉婷[33]對克隆植物毛竹(Phyllostachys edulis)的研究發現,毛竹的單個克隆最大延伸距離可達1448 m,這為毛竹的資源整合提供了一定的理論基礎。蕨麻是典型的克隆植物,在野外長年的生長增大了繁殖面積,一定程度上也影響著遺傳距離的大小。6 個蕨麻居群不同距離分布的個體聚類結果表明,每個地區1~30 號樣品間地理距離較短,均在5 km 以內,31~36 號樣品間,地理距離較遠,均在5 km 以上。其中,祁連縣居群蕨麻個體間遺傳距離與地理距離相關性最好,可以明顯看出,同一地區蕨麻居群內,蕨麻個體間遺傳距離隨地理距離的增加,呈增大的態勢,其中1~30 號樣品的遺傳距離較近,31~35 號樣品間遺傳距離較遠,根據地理距離分布,可劃分為5 km 以內的樣品個體和5 km 以上的樣品個體。其他地區遺傳距離與地理距離的相關性雖然不是很大,但根據居群聚類分析可以得出,5 km 是一個分界點,即5 km 可以作為一個克隆的延長距離。
由于在實際采樣過程中,地理情況和采樣點的分布情況均較為復雜。祁連縣的蕨麻個體間遺傳距離與地理距離表現出較好的線性關系(R2=0.1983,P=0.01<0.05),其遺傳距離和地理距離呈正相關,且遺傳多樣性最高(H=0.2797,I=0.4287)。而其他居群蕨麻個體間遺傳距離并不是按照地理距離線性排布,造成該情況的發生可能有以下幾點原因:首先,不能排除蕨麻生長過程中出現種子傳播的可能性;其次,蕨麻是無性繁殖為主,有性繁殖為輔,這會導致野生蕨麻具有基因交流的過程;最后,蕨麻采樣過程中,同一居群內雖然有地理海拔差距,但并沒有大型的山脈等明顯的地理隔離,使居群內部沒有產生生殖隔離,所以同一地區的蕨麻遺傳距離較近,從而降低了與地理距離的相關性。
根據居群聚類結果可知,蕨麻聚類一般以地理距離和地理阻隔結合進行聚類,碌曲縣和河南縣兩個居群地理位置近,聚為一支(遺傳距離=0.0141,遺傳相似系數=0.986),可能是因為該地勢較為平坦,利于蟲媒、風媒的傳播,增加兩個居群之間的基因流[39],從而使得兩個居群之間變異少,聚為一支。碌曲縣與若爾蓋縣地理位置也近,但由于有大巴山脈(海拔為2000~2500 m)作為阻隔,阻礙了有性繁育花粉的傳播,減少了基因流,使得其沒有聚在一起(遺傳距離=0.0256,遺傳相似系數=0.9747)。南木林縣、祁連縣、八宿縣3 個居群之間的聚類也較遠,其間不僅地理距離遠,更有大型山脈作為阻隔,阻斷了居群之間的交流。由此可看出,居群可以以大型山脈進行區分。
4 結論
結合采樣點地理分布、遺傳多樣性和聚類分析可知:1)當采樣地勢較為平坦時,可以有效增加居群間的基因交流,在采樣時,需要增大居群之間的距離以避免采集為同一親本。碌曲縣和河南縣兩個地區的直線距離為100 km,兩個居群遺傳距離相似度最高,因此在沒有大型山脈隔離的情況下,蕨麻有性繁殖即花粉傳播距離可能達到100 km,花粉在兩地之間進行長距離傳播,促使居群之間基因交流,從而使得兩地的遺傳關系近,該距離可以作為在沒有大型山脈隔離條件下,蕨麻居群間采樣的最小距離;2)當有山脈作為采樣隔斷時,居群之間的地理距離即使比較近,但由于山脈的阻斷,居群之間的基因流降低,生境的隔離也成為居群之間變異較大的重要原因,因此,山脈隔斷可以有效避免采集樣本來自同一居群,居群間采樣應該以翻越青藏高原大型山脈后采集樣品為宜;3)根據居群內聚類分析可以得出,多個居群中均顯示,30 號樣品之前大多聚在一起,地理距離在5 km 之內,而在5 km之外的樣品則單獨聚在一起,說明在一定范圍內(5 km 左右)的分株都來源于同一親本。因此采樣時,一個居群內個體間的距離至少達到5 km 。
根據研究結果,對6 個野生蕨麻居群,提出針對性保護建議:除了主要采取就地保護策略保障其原始居群的遺傳多樣性外,還可以適當對不同地區蕨麻種質資源進行遷地保護,維持其基因流的穩定性。蕨麻在西部地區生長好,且在海拔1700 m 以上地區塊根才膨大,西部地區大部分位于我國第一階梯,海拔較高,最合適蕨麻生長,且蕨麻繁殖擴增的模式也能為西部地區貧瘠土地的修復帶來良好的生態效益。同時,蕨麻在農牧民日常生活中既可以作為食品食用,也可以作為商品進行售賣以增加收入,因此在西部地區,蕨麻在秋季時會受到大面積的采挖,采挖過后的蕨麻來年生長勢急劇下降,從而影響其生態修復能力。因此,提出以下建議,比如青海省內蕨麻居群遺傳多樣性高的祁連縣居群、甘肅省的碌曲縣居群以及西藏自治區的八宿縣居群,可以將蕨麻現有居群建立自然保護區,適當開展遷地保護,規劃采挖區域,以減少過度采挖對蕨麻多樣性和生態的破壞。遺傳多樣性較低的居群,比如河南縣居群、若爾蓋居群,可以對其進行種質篩選,培育出純品種,將其作為一個品種來進行研究,發掘其藥用、食用以及生態價值。同時,研究中也發現同個省份內不同居群之間遺傳多樣性有差異,本試驗在同一省份內采樣居群數量較少,只能部分概括同一區域內的遺傳多樣性結果,在后續研究中,可以對某一區域通過不同種群野生資源進行遺傳資源研究,通過分子生物學方法探究其遺傳多樣性,探討其不同海拔下不同自然居群的遺傳多樣性,從不同生境下居群的遺傳結構進行分子變異分析,再確定其保護策略。
