基于SSR 標(biāo)記的河南省假儉草群體遺傳多樣性研究
2023-09-25 03:42:56吉軼楠任雪鋒茍?zhí)鹛?/span>臧國(guó)長(zhǎng)鄭軼琦
草業(yè)學(xué)報(bào) 2023年9期
關(guān)鍵詞:研究
吉軼楠,任雪鋒,茍?zhí)鹛穑皣?guó)長(zhǎng),鄭軼琦
(河南科技大學(xué)園藝與植物保護(hù)學(xué)院,河南 洛陽(yáng) 471000)
假儉草(Eremochloa ophiuroides)為禾本科(Gramineae)蜈蚣草屬(Eremochloa)的一種多年生草本植物,是世界三大暖季型草坪草之一[1]。假儉草因植株低矮、青綠期長(zhǎng)、適應(yīng)性強(qiáng)而被廣泛應(yīng)用于草坪綠化、建植運(yùn)動(dòng)場(chǎng)草坪、邊坡防護(hù)和庫(kù)區(qū)生態(tài)改良等[2]。前人利用DNA 擴(kuò)增指紋(DNA amplified fingerprints, DAF)、擴(kuò)增片段長(zhǎng)度多態(tài)性(amplified fragment length polymorphism, AFLP)、隨機(jī)擴(kuò)增多態(tài)性DNA(random amplified polymorphic DNA, RAPD)、簡(jiǎn)單序列間重復(fù)序列(inter-simple sequence repeat, ISSR)、相關(guān)序列擴(kuò)增多態(tài)性(sequencerelated amplified polymorphism, SRAP)、簡(jiǎn)單重復(fù)序列(simple sequence repeat, SSR)分子標(biāo)記技術(shù)已開(kāi)展了大量的假儉草遺傳多樣性研究[3-14],Weaver 等[3]利用DAF 研究美國(guó)5 個(gè)假儉草種質(zhì)之間的遺傳關(guān)系,結(jié)果表明多態(tài)性水平相對(duì)較低,供試種質(zhì)可能有共同的來(lái)源。我國(guó)學(xué)者利用AFLP 標(biāo)記[4]、RAPD 標(biāo)記[5-6]、ISSR 標(biāo)記[7-8]、SRAP 標(biāo)記[11]研究我國(guó)的假儉草種質(zhì)資源的遺傳多樣性,結(jié)果均表明假儉草具有較豐富的種質(zhì)資源遺傳多樣性。Li 等[14]基于下一代測(cè)序技術(shù)開(kāi)發(fā)了大量假儉草SSR 引物序列,并研究我國(guó)43 個(gè)假儉草種質(zhì)資源遺傳多樣性,結(jié)果顯示供試假儉草具有較豐富的遺傳多樣性。
在全球氣候變化的背景下,研究人員越來(lái)越關(guān)注溫度、降水和日照等環(huán)境因素對(duì)植物種群分布格局的影響。由于氣候和環(huán)境異質(zhì)性,種群面臨不同程度的選擇壓力[15],植物種群的位置固定且擴(kuò)散能力較低,因此在環(huán)境異質(zhì)性中面臨的環(huán)境選擇壓力較大,可能導(dǎo)致植物基因型頻率的差異[16],產(chǎn)生適應(yīng)性進(jìn)化,以不斷響應(yīng)生境的改變[17-18]。Hamasha 等[19]利用AFLP 分子標(biāo)記研究了來(lái)自約旦不同生物氣候區(qū)4 種針茅(Stipa capillata)種群,結(jié)果表明其遺傳多樣性與溫度和降水相關(guān),環(huán)境梯度顯著影響針茅種群的遺傳結(jié)構(gòu)。Wu 等[20]利用SSR 標(biāo)記研究中國(guó)58 個(gè)穗狀狐尾藻(Myriophyllum spicatum)種群,結(jié)果表明環(huán)境和地理隔離共同影響狐尾藻種群遺傳結(jié)構(gòu),其中環(huán)境的影響較大。Gao 等[21]利用SSR 標(biāo)記研究中國(guó)黃河三角洲的30 個(gè)蘆葦(Phragmites australis)自然斑塊,結(jié)果表明環(huán)境異質(zhì)性與蘆葦種群遺傳結(jié)構(gòu)變異密切相關(guān)。在種群歷史動(dòng)態(tài)、環(huán)境異質(zhì)性和人類的干擾下,物種生境的片段化和隔離已成為野生種質(zhì)資源保護(hù)中的一個(gè)嚴(yán)重問(wèn)題。這種隔離對(duì)種群遺傳多樣性的影響由于受多種因素干擾一直是研究的熱點(diǎn)[22-23]。生境的片段化和隔離通常導(dǎo)致形成分布區(qū)核心和邊緣居群。邊緣居群是指位于物種分布區(qū)的邊緣區(qū)域的居群[24],由于邊緣居群生境較核心居群惡劣,對(duì)環(huán)境變化的響應(yīng)可能更明顯,物種對(duì)于環(huán)境變化的響應(yīng)主要取決于邊緣居群的響應(yīng)[25],因此,研究邊緣居群的遺傳多樣性與群體遺傳結(jié)構(gòu)對(duì)種質(zhì)資源保護(hù)十分重要。關(guān)于核心和邊緣居群的遺傳多樣性,有兩種不同的觀點(diǎn)。第一種觀點(diǎn)認(rèn)為,由于邊緣居群數(shù)量少、孤立,處于適宜生境的邊緣,因此可能會(huì)遭受近親繁殖和遺傳漂變,從而導(dǎo)致邊緣居群的遺傳多樣性較低。許多先前的研究表明,核心居群的遺傳多樣性高于邊緣居群[25-26],另一種觀點(diǎn)認(rèn)為,邊緣居群經(jīng)歷高度自然選擇,強(qiáng)烈的選擇壓力將篩選出具有優(yōu)質(zhì)基因的種質(zhì)資源并產(chǎn)生新物種的來(lái)源,造成邊緣居群的遺傳多樣性較高[27-29]。李霓等[27]利用SSR 分子標(biāo)記研究肋果沙棘(Hippophae neurocarpa)北緣5 個(gè)居群,結(jié)果表明肋果沙棘北緣居群遺傳多樣性豐富,居群內(nèi)變異較居群間大。Van 等[28]研究來(lái)自法國(guó)和盧森堡21 個(gè)蠅子草(Silene nutans)邊緣和核心居群等位酶的變異,結(jié)果顯示其邊緣種群的遺傳多樣性較高。Liu 等[29]利用SSR 分子標(biāo)記研究了我國(guó)4 個(gè)省毛紅椿(Toona ciliatavar.pubescens)核心和邊緣居群的遺傳多樣性,結(jié)果表明位于云貴高原的邊緣種群,在適應(yīng)高原生境的過(guò)程中分化出不同生態(tài)型或基因型從而導(dǎo)致其遺傳多樣性水平高于核心種群。Shi 等[30]利用SSR 分子標(biāo)記研究我國(guó)6 個(gè)省12 個(gè)核心苦櫧(Castanopsis sclerophylla)種群和5 個(gè)邊緣種群,結(jié)果表明邊緣苦櫧種群遺傳多樣性更高。假儉草主要分布在長(zhǎng)江流域和我國(guó)南部地區(qū),包括河南、四川、湖北、湖南、廣西和廣東等17 個(gè)省市[31],根據(jù)假儉草的分布及前人對(duì)假儉草種質(zhì)的調(diào)查收集研究[10]可知,河南省是假儉草在華中地區(qū)分布的北緣,河南省的假儉草種質(zhì)資源主要分布于大別山和桐柏山[32]。因此探究河南省假儉草種質(zhì)資源的遺傳多樣性與影響其分布的因素,對(duì)假儉草的開(kāi)發(fā)保護(hù)及利用具有重要意義。綜上所述,本研究利用SSR 技術(shù)研究河南省假儉草種質(zhì)資源的遺傳多樣性與群體遺傳結(jié)構(gòu),探究地理距離、環(huán)境因子對(duì)群體結(jié)構(gòu)的影響,旨在揭示河南省假儉草種質(zhì)資源的遺傳多樣性及種群遺傳分布格局形成的原因,為假儉草種質(zhì)資源的收集、開(kāi)發(fā)及保護(hù)奠定基礎(chǔ),為今后進(jìn)一步開(kāi)展轉(zhuǎn)錄組學(xué)研究,進(jìn)行基因功能注釋,探討河南省假儉草適應(yīng)性進(jìn)化機(jī)制提供理論依據(jù)。
1 材料與方法
1.1 試驗(yàn)材料
根據(jù)中國(guó)數(shù)字植物標(biāo)本館(https://www.cvh.ac.cn)、《河南植物志》[32]以及相關(guān)文獻(xiàn)記載[10],結(jié)合本課題組前期的實(shí)地踏查發(fā)現(xiàn)河南省野生假儉草主要分布于河流沿岸,本研究于2021 年4-10 月根據(jù)其分布現(xiàn)狀選取8個(gè)假儉草野生居群(C1~C8),居群內(nèi)取樣間隔大于10 m,采集無(wú)病蟲(chóng)害的健康葉片,共采集141 份材料,儲(chǔ)存在裝有硅膠的密封袋中,以備后期試驗(yàn)。8 個(gè)居群分布情況見(jiàn)圖1,采樣地及樣本數(shù)量詳見(jiàn)表1。部分居群受人為因素破壞,樣本取樣數(shù)量不足20 份,其中C1居群位于水稻(Oryza sativa)田的田埂上,大面積水稻種植使假儉草生境破壞嚴(yán)重,該居群只采集到5 份材料;C6居群位于正在施工的橋梁下,只采集到16 份材料。
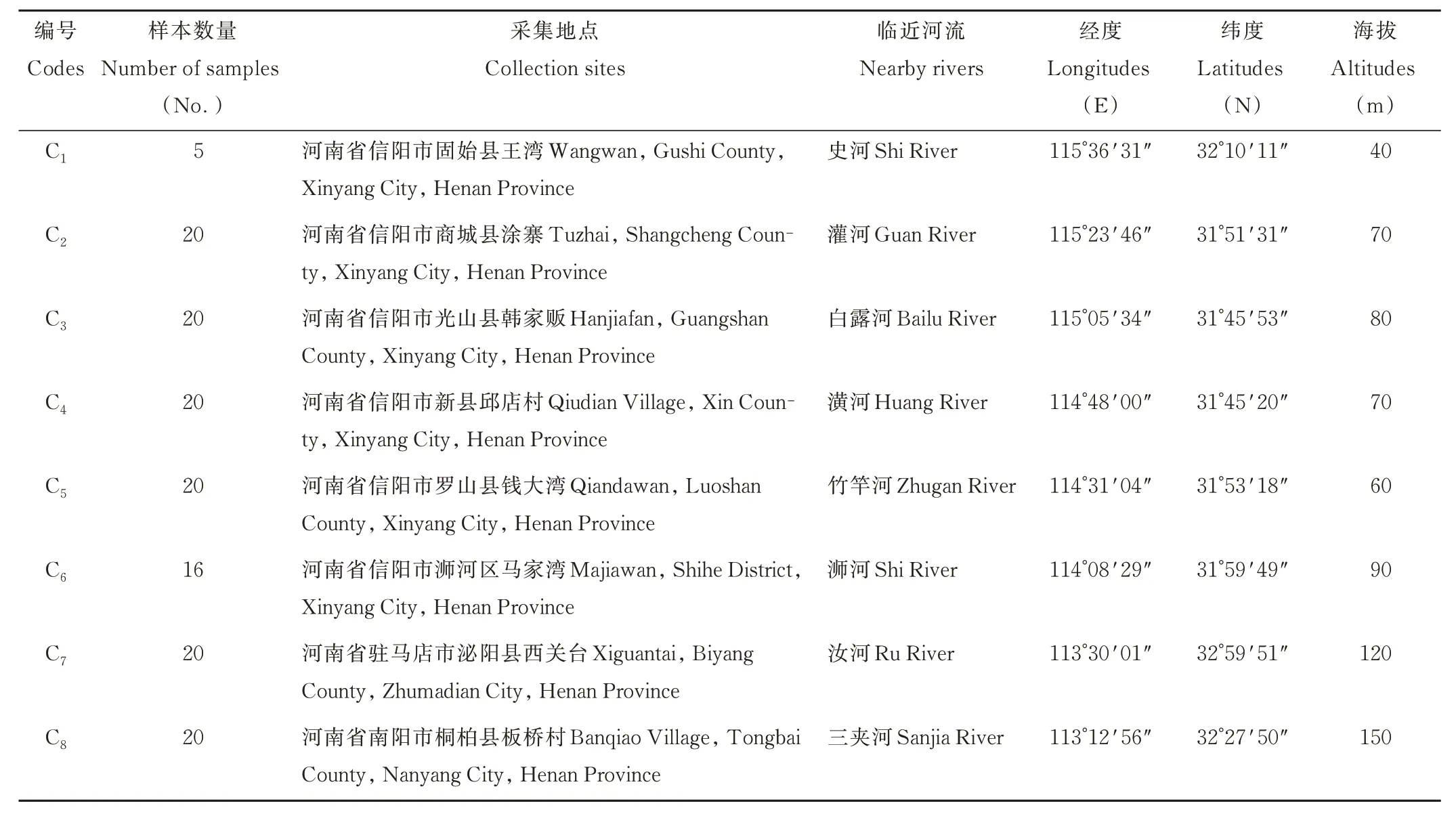
表1 假儉草采集地概況Table 1 General situation of collecting areas of centipedegrass
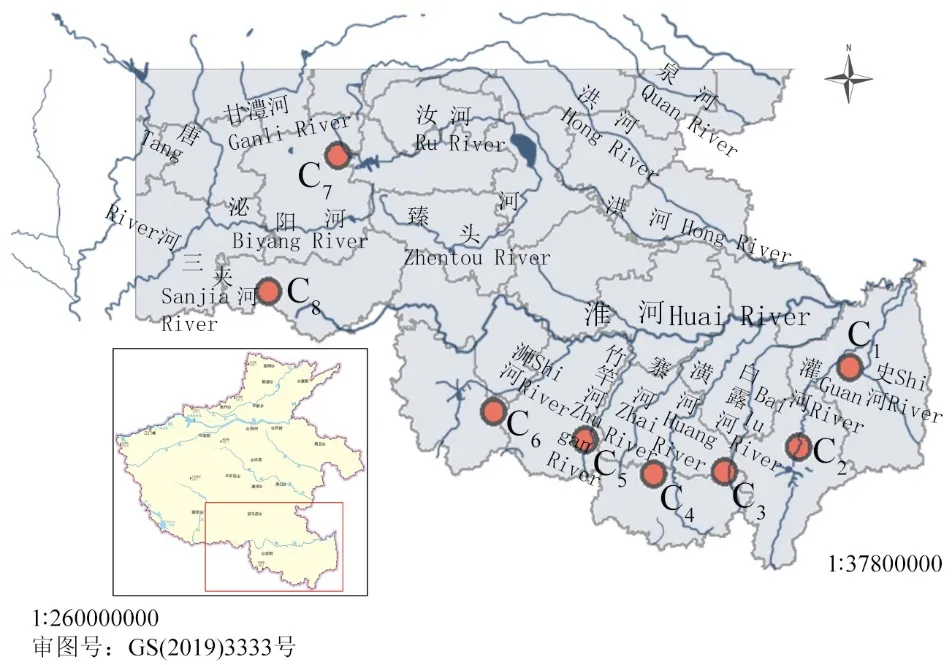
圖1 8 個(gè)假儉草居群的地理分布Fig.1 Geographic distribution of 8 centipedegrass populations
1.2 氣候數(shù)據(jù)
假儉草采集地氣候數(shù)據(jù)獲取自WorldClim 網(wǎng)站(www.worldclim.org),利用Arcgis 10.2 軟件根據(jù)采集地經(jīng)緯度提取。從WordClim 網(wǎng)站獲取假儉草居群的19 個(gè)環(huán)境變量,分別是年平均溫、年降水量、溫度年變化范圍、溫度季節(jié)性、降水季節(jié)、等溫性、平均日較差、最熱月最高溫度、最暖季平均氣溫、最暖季降水、最干旱月份的降水量、最冷季平均氣溫、最冷月份最低溫度、最冷季降水、最干燥季平均氣溫、最干燥季降水、最濕季平均溫度、最濕季月份降水量、最濕季降水。為了避免過(guò)高估計(jì)環(huán)境變量的貢獻(xiàn),剔除12 個(gè)Pearson 強(qiáng)相關(guān)的環(huán)境變量,保留7 個(gè)環(huán)境變量(表2)。
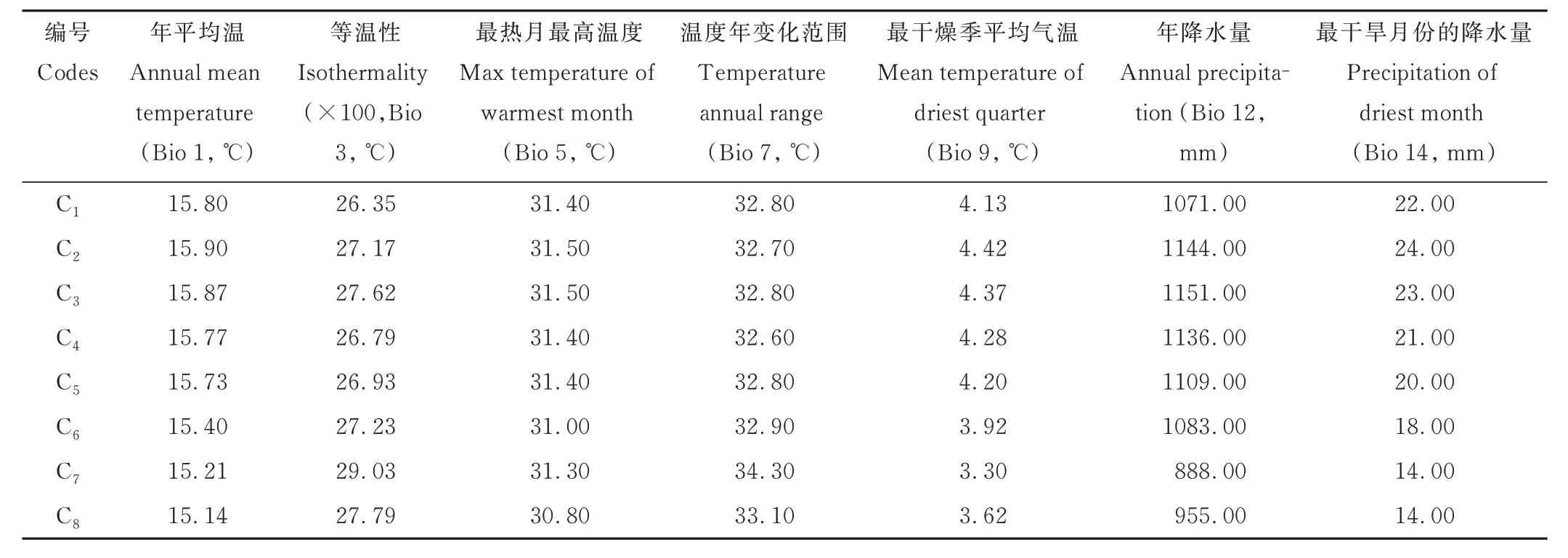
表2 提取的假儉草居群的環(huán)境變量Table 2 Environmental variables extracted from the populations of centipedegrass
1.3 試驗(yàn)方法
利用北京天根生化科技有限公司的DP350-02 植物DNA 提取試劑盒提取供試材料的DNA。分別用1%的瓊脂糖凝膠電泳、紫外分光光度計(jì)(T6 新世紀(jì),北京普析通用儀器有限責(zé)任公司)檢測(cè)所提取DNA 的質(zhì)量、濃度及純度,并將DNA 溶液稀釋至30 ng·μL-1,保存在-20 ℃環(huán)境。從Li 等[33]的報(bào)道中選用30 對(duì)多態(tài)性信息含量(polymorphism information content, PIC)高的假儉草SSR 引物進(jìn)行預(yù)試驗(yàn),最終篩選出10 對(duì)多態(tài)性高且重復(fù)性穩(wěn)定的引物(表3),擴(kuò)增反應(yīng)體系和程序參照Li 等[33]的方法。

表3 本研究所采用的 SSR 引物序列Table 3 Sequences of SSR primers used in the study
1.4 數(shù)據(jù)分析
對(duì)擴(kuò)增得到的帶型用GeneMarker 進(jìn)行判定。用GenAlEx 6.5[34]轉(zhuǎn)換數(shù)據(jù)格式,進(jìn)行分子方差分析(analysis of molecular variance, AMOVA)。用POPGENE 1.32[35]分析遺傳多樣性的相關(guān)參數(shù):等位基因數(shù)(observed number of alleles, Na)、有 效 等 位 基 因 數(shù)(effective number of alleles, Ne)、Shannon’s 遺 傳 多 樣 性 信 息 指 數(shù)(Shannon’s information index,I)、Nei’s 基 因 多 樣 性 指 數(shù)(Nei’s gene diversity,H)、觀 測(cè) 雜 合 度(observed heterozygosity, Ho)及期望雜合度(expected heterozygosity, He)。并用Cervus 3.07[36]計(jì)算多態(tài)性信息含量、等位基因頻率。
多態(tài)性信息含量(polymorphic information content, PIC)計(jì)算公式為:
式中:Pi為第i個(gè)等位基因頻率。
用NTSYS-pc 2.1 軟件[37]的非加權(quán)組平均法(unweighted pair-group method with arithmetic means, UPGMA)進(jìn)行聚類,并對(duì)地理距離與遺傳距離進(jìn)行Mantel 檢驗(yàn)。根據(jù)供試材料的遺傳距離用MEGA 6.0[38]得到141 份假儉草的樹(shù)狀聚類圖,Bootstrap 的初始值設(shè)為1000 來(lái)檢驗(yàn)發(fā)育樹(shù)的可信度。用Structure 2.3.4[39]分析假儉草的群體遺傳結(jié)構(gòu),為了計(jì)算K 值,本研究從1~10 依次測(cè)試,對(duì)每個(gè)K 值獨(dú)立運(yùn)行10 次。并將結(jié)果導(dǎo)入在線程序Structure harvester(http://taylor0.biology.ucla.edu/structureHarvester/)預(yù)測(cè)最佳群體數(shù),依據(jù)Q 值>0.7[40]來(lái)劃分不同亞群。用R 語(yǔ)言ade4 工具包[41]進(jìn)行主坐標(biāo)分析(principal coordinate analysis,PCoA)。用CANOCO 5.0 進(jìn)行冗余分析(redundancy analysis, RDA)[42]來(lái)檢測(cè)環(huán)境變量對(duì)遺傳結(jié)構(gòu)的相對(duì)貢獻(xiàn)。利用軟件BayeScan 2.1[43]計(jì)算離群位點(diǎn),將后驗(yàn)概率>0.76,PO>0.5 的log10 值視為異常位點(diǎn),用ARLEQUIN 3.5 軟件[44]中的分層孤島模型計(jì)算異常位點(diǎn),將等位基因頻率>95%的位點(diǎn)判斷為異常位點(diǎn)。將兩種方法共同檢測(cè)到的視為最終確定的異常位點(diǎn)。用R 語(yǔ)言包LEA 中的潛在因子混合模型(latent factor mixed model,LFMM)[45]進(jìn)行環(huán)境位點(diǎn)關(guān)聯(lián)分析,將檢測(cè)結(jié)果中|Z|>3,P<0.001 的視為環(huán)境關(guān)聯(lián)位點(diǎn)。
2 結(jié)果與分析
2.1 SSR 擴(kuò)增結(jié)果
10 對(duì)引物共檢測(cè)到99 個(gè)等位基因(表4)。單個(gè)位點(diǎn)觀測(cè)等位基因數(shù)為6~12 個(gè),平均為9.90 個(gè)。引物多態(tài)性信息含量(PIC)為0.53~0.88,引物TJIB.EO-92 和TJIB.EO-51 的PIC 值最高,引物TJIB.EO-40 的PIC 值最低,10 對(duì)引物的平均值為0.78。引物TJIB. EO-51 和TJIB. EO-90 檢測(cè)到的等位基因數(shù)量最多(12.00),引物TJIB.EO-40 檢測(cè)到的等位基因最少(6.00)。有效等位基因數(shù)為2.54~8.96,最大的為引物TJIB.EO-92,最小的為引物TJIB.EO-40,平均為6.12。Shannon’s 遺傳多樣性信息指數(shù)為1.11~2.31,最高的是引物TJIB.EO-51,最低的是引物TJIB.EO-40,平均為1.91。Nei’s 基因多樣性指數(shù)為0.61~0.89,最高的是引物TJIB.EO-92和TJIB.EO-51,最低的是引物TJIB.EO-40,平均為0.81。觀測(cè)雜合度為0.26~0.99,平均為0.71,期望雜合度為0.61~0.89,平均為0.81。大多數(shù)(60.00%)位點(diǎn)的期望雜合度大于觀測(cè)雜合度。結(jié)果表明,本研究所用的引物均具有較高的多態(tài)性。
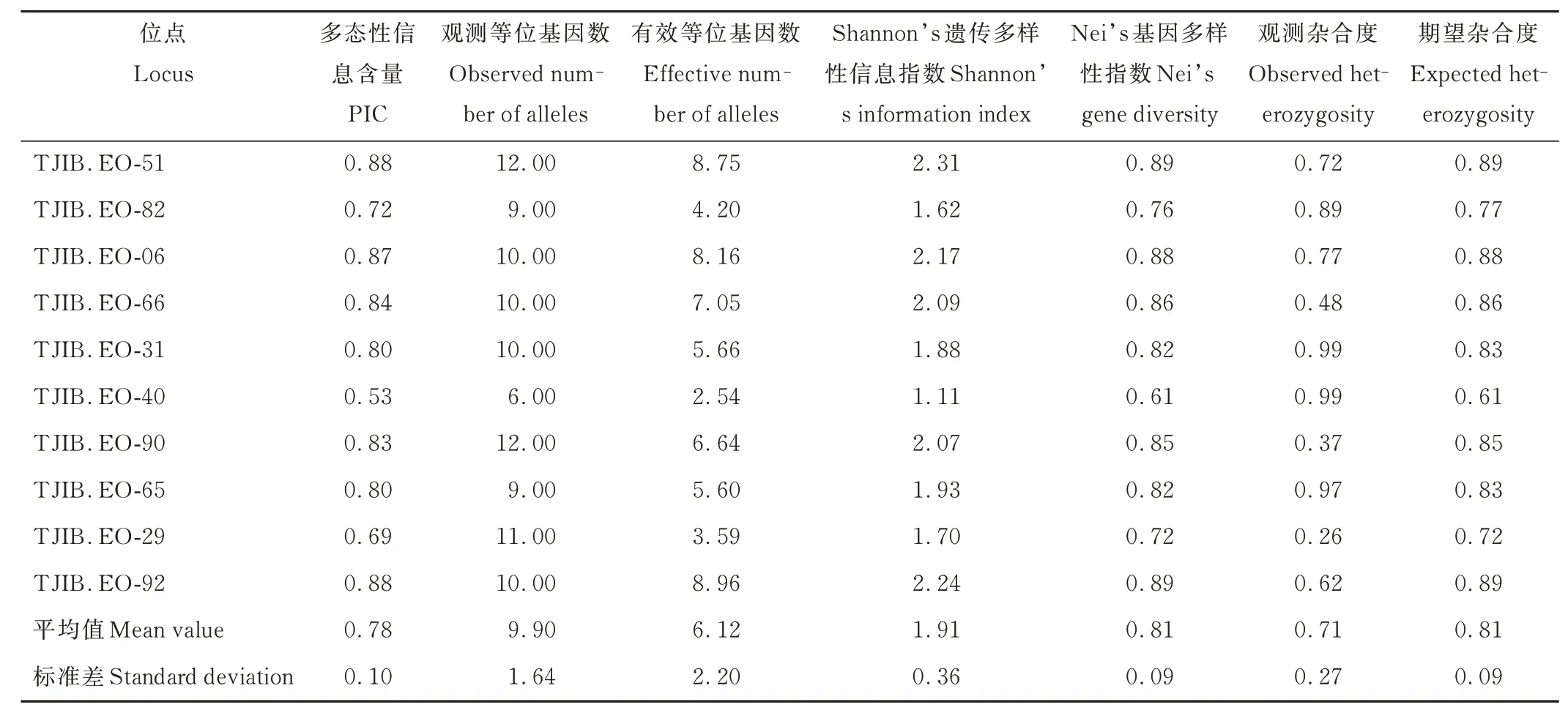
表4 引物擴(kuò)增結(jié)果Table 4 Primers amplification results
2.2 遺傳多樣性及方差分析
供試假儉草的平均Shannon’s 遺傳多樣性信息指數(shù)和Nei’s 基因多樣性指數(shù)分別為1.44 和0.69,多態(tài)位點(diǎn)百分率為98.75%(表5),表明供試材料的遺傳多樣性水平較高。各居群觀測(cè)等位基因數(shù)為4.30~7.90,平均值為6.14,其中C4居群最大,C1居群最小。有效等位基因數(shù)為3.11~4.97,平均值為3.94,其中C4居群最大,C8居群最小。Shannon’s 遺傳多樣性信息指數(shù)為1.15~1.73,平均值為1.44,其中C4居群最大,C8居群最小。Nei’s 基因多樣性指數(shù)為0.61~0.77,平均值為0.69,其中C4居群最大,C8居群最小。觀測(cè)雜合度為0.58~0.79,平均值為0.69,其中C7居群最大,C1居群最小。期望雜合度為0.63~0.79,平均值為0.72,其中C4居群最大,C8居群最小。C4居群的觀測(cè)等位基因數(shù)、Shannon’s 遺傳多樣性信息指數(shù)、期望雜合度、有效等位基因數(shù)及Nei’s 基因多樣性指數(shù)均最大。綜上所述,各居群的不同遺傳多樣性指標(biāo)存在一定差異,基于Shannon’s 遺傳多樣性信息指數(shù),居群遺傳多樣性從大到小依次為C4>C3>C6>C2>C5>C7>C1>C8。
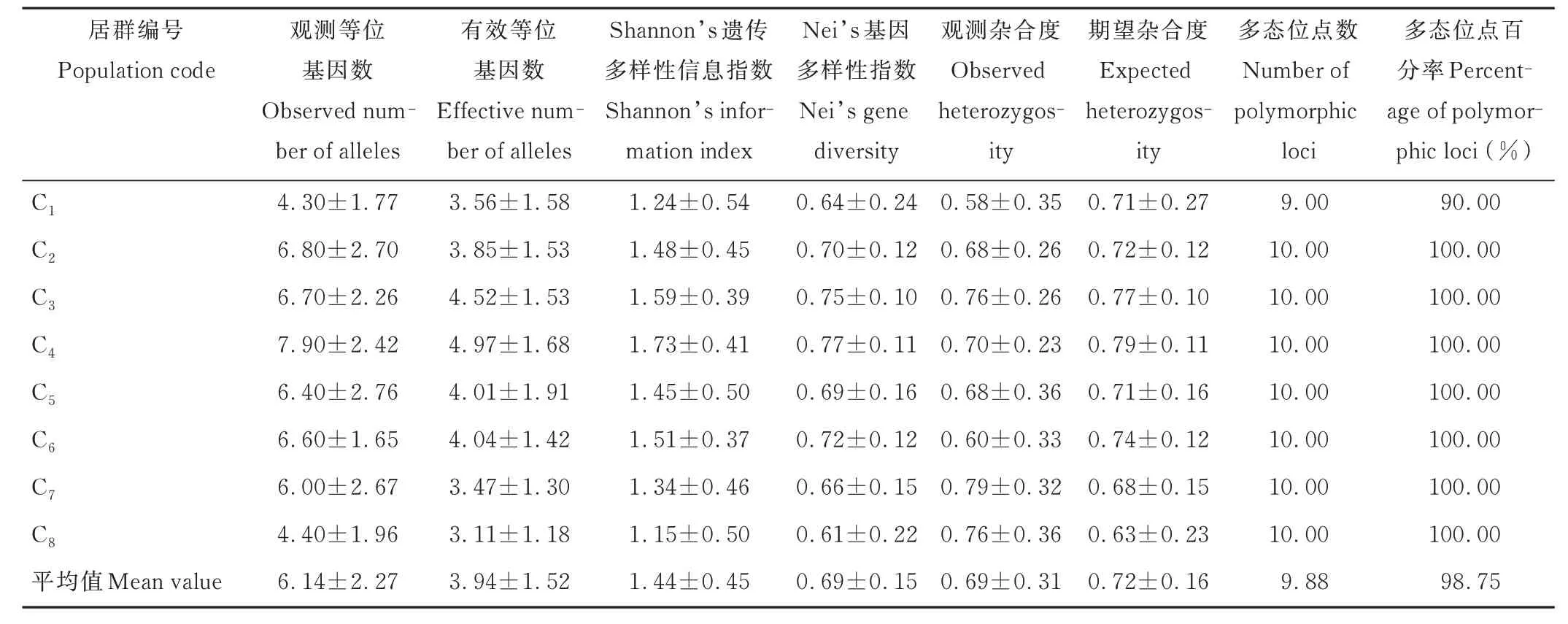
表5 假儉草居群的遺傳多樣性Table 5 Genetic diversity of centipedegrass populations (mean±SD)
假儉草居群SSR 位點(diǎn)F統(tǒng)計(jì)結(jié)果(表6)表明,居群間的遺傳分化系數(shù)(fixation index, Fst)為0.06~0.35,平均值為0.14,說(shuō)明居群存在中度遺傳分化。采用分子方差分析居群間和居群內(nèi)的遺傳分化程度(表7),居群間的遺傳分化貢獻(xiàn)為13.04%,而居群內(nèi)的遺傳分化貢獻(xiàn)為86.96%,表明河南假儉草居群的遺傳分化主要發(fā)生于居群內(nèi)。基因流(number of effective migrants, Nm)均值為1.49,表明各居群間存在較頻繁的基因交流(表6)。
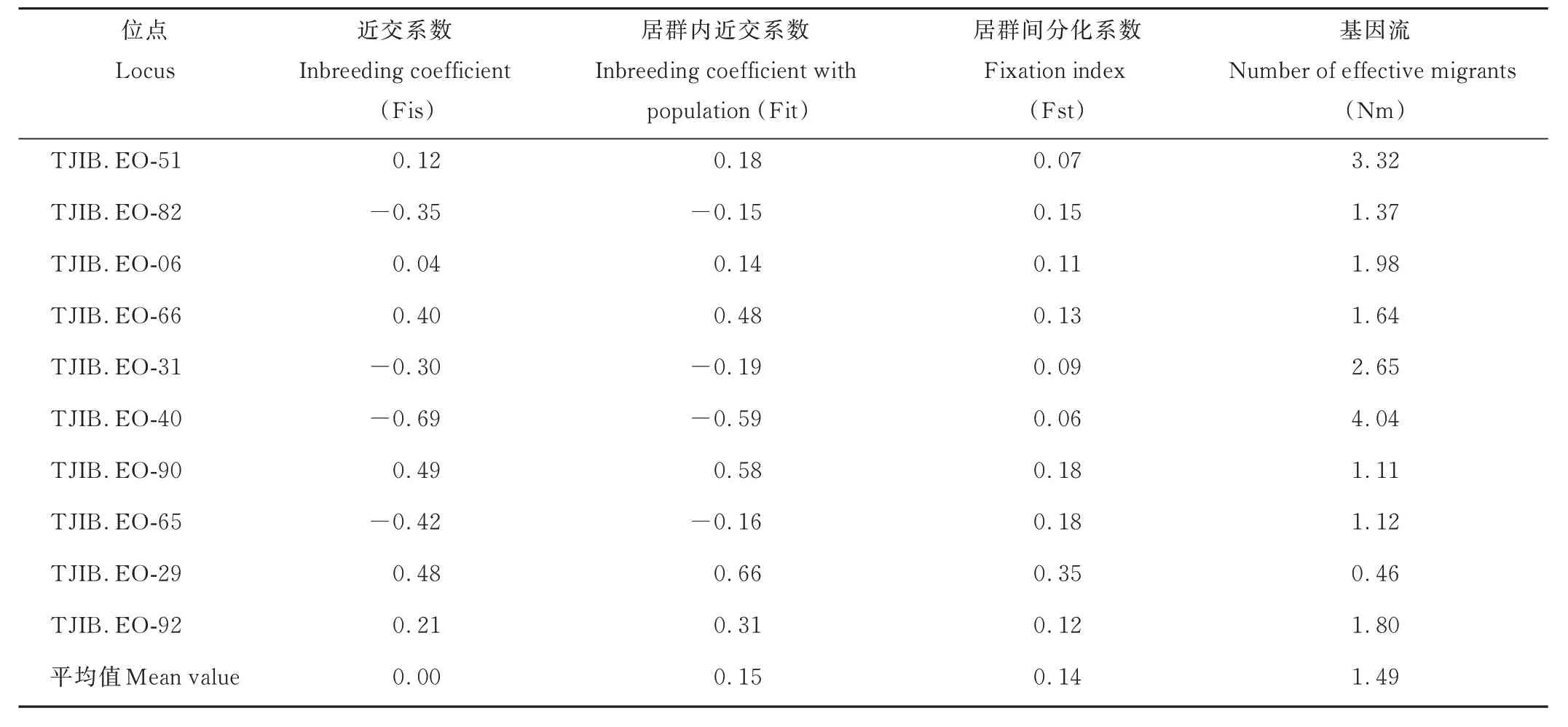
表6 假儉草居群SSR 位點(diǎn)F 統(tǒng)計(jì)Table 6 F-statistics of SSR in populations of centipedegrass

表7 分子方差分析Table 7 Molecular analysis of variance
2.3 聚類分析及Mantel 檢驗(yàn)
8 個(gè)居群UPGMA 聚類分析結(jié)果(圖2)表明,在遺傳距離為0.15 處,供試材料共分為3 組,第Ⅰ組包含C1和C6居群,第Ⅱ組包含C7和C8居群,第Ⅲ組包含C2、C3、C4和C5居群。結(jié)果顯示8 個(gè)居群沒(méi)有完全按照地理來(lái)源分組,如地理來(lái)源較遠(yuǎn)的C1和C6居群聚為一組。Mantel 檢驗(yàn)結(jié)果表明遺傳距離與地理距離間無(wú)相關(guān)性(r=0.30,P=0.95>0.05)。
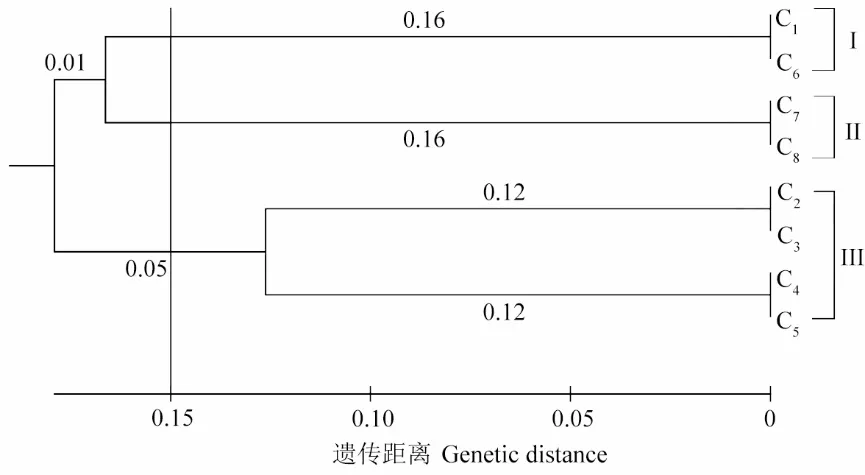
圖2 8 個(gè)居群間UPGMA 聚類分析Fig.2 UPGMA cluster analysis among 8 populations
聚類分析結(jié)果(圖3)表明,141 份假儉草材料分為3 組,第Ⅰ組包含C1、C2、C7、C8居群的全部材料,C3居群的6份材料及C6居群的4 份材料;第Ⅱ組包含C5居群的全部材料,C4居群的8 份材料及C6居群的12 份材料;第Ⅲ組包含C3居群的14 份材料和C4居群的12 份材料。結(jié)果表明,部分來(lái)自同一居群的材料被分到不同組,如來(lái)自C3居群的26~30 號(hào)材料被聚在第Ⅰ組,而31~45 號(hào)材料被聚在第Ⅲ組。
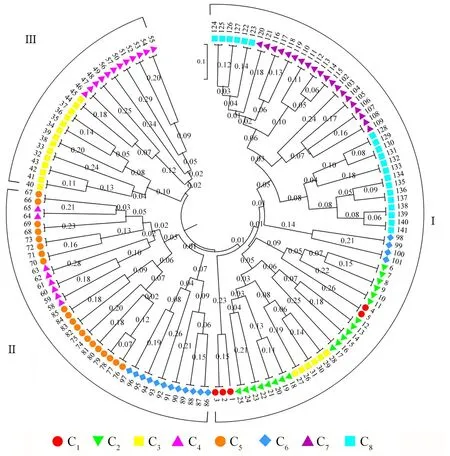
圖3 141 份假儉草材料的UPGMA 聚類分析Fig.3 UPGMA cluster analysis of 141 centipedegrass materials
2.4 群體遺傳結(jié)構(gòu)分析和主坐標(biāo)分析
群體遺傳結(jié)構(gòu)分析結(jié)果表明,當(dāng)K=3 時(shí),ΔK 達(dá)到峰值(圖4),表明供試的141 份假儉草材料被分為3 組(圖5),第1 個(gè)亞群(紅色)主要包括C1、C2、C3居群共47 份材料(33.33%),第2 個(gè)亞群(藍(lán)色)主要包括C4、C5、C6居群共39 份材料(27.66%),第3 個(gè)亞群(綠色)主要包括C7、C8居群共34 份材料(24.11%),其他21 份材料(14.93%)的Q 值<0.7,屬于遺傳背景復(fù)雜的混合型群體。
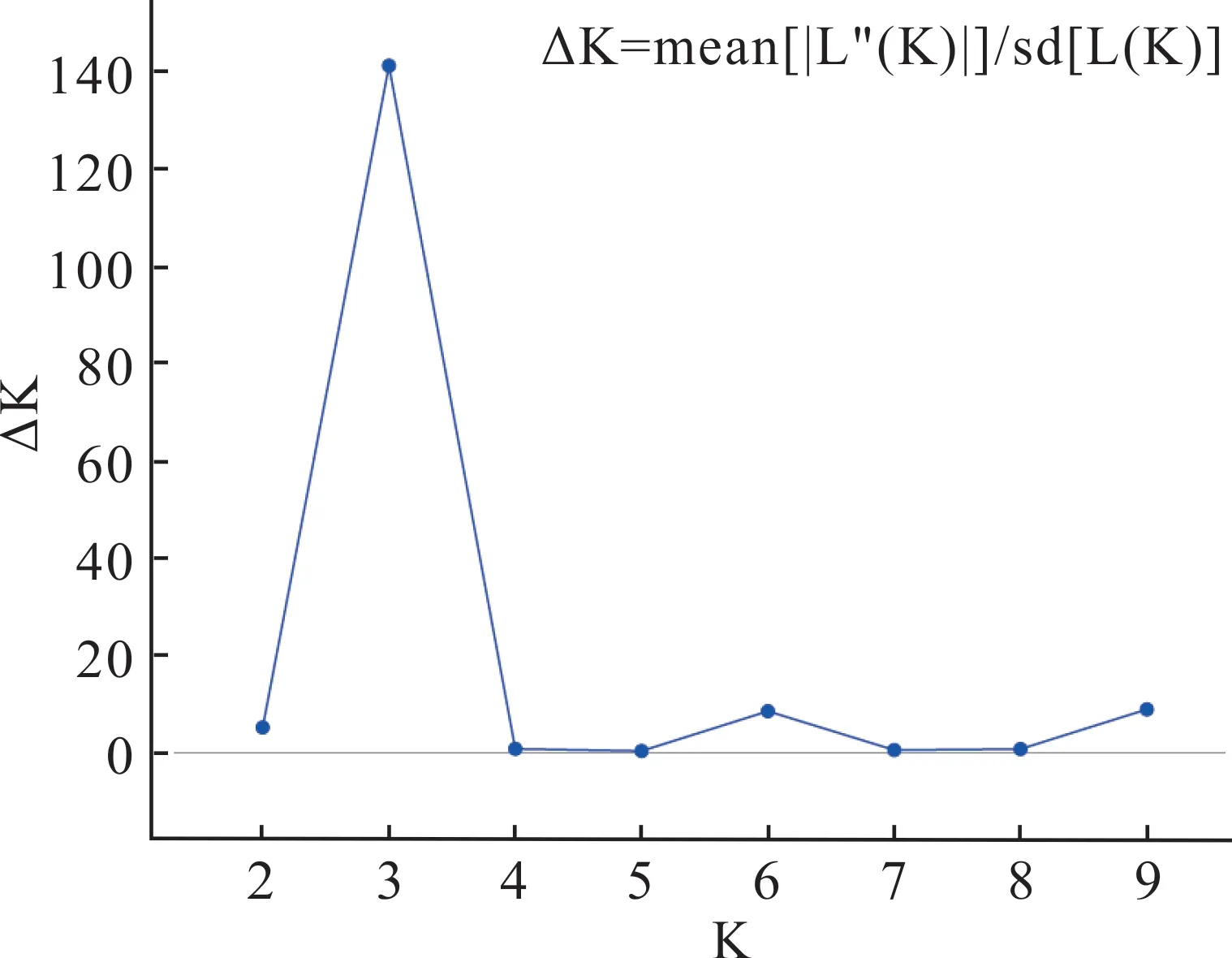
圖4 K 與?K 的變化關(guān)系Fig.4 Variation of K and ?K
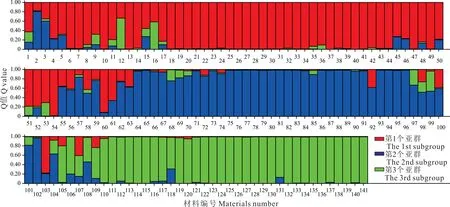
圖5 基于SSR 標(biāo)記的K=3 時(shí)141 份假儉草群體遺傳結(jié)構(gòu)Fig.5 Genetic structure of 141 centipedegrass populations at K=3 based on SSR markers
主坐標(biāo)分析結(jié)果表明,第1 主成分對(duì)群體遺傳變異的解釋為12.04%,第2 主成分為9.25%(圖6)。圖中材料位置越靠近表明兩者親緣關(guān)系越近,反之表明兩者親緣關(guān)系較遠(yuǎn)。PCoA 分析結(jié)果顯示將材料明顯分為3 個(gè)亞群,分別是S1(紅色)、S2(藍(lán)色)和S3(綠色),混合群(黃色)介于3 個(gè)亞群之間,與群體遺傳結(jié)構(gòu)分析結(jié)果一致。
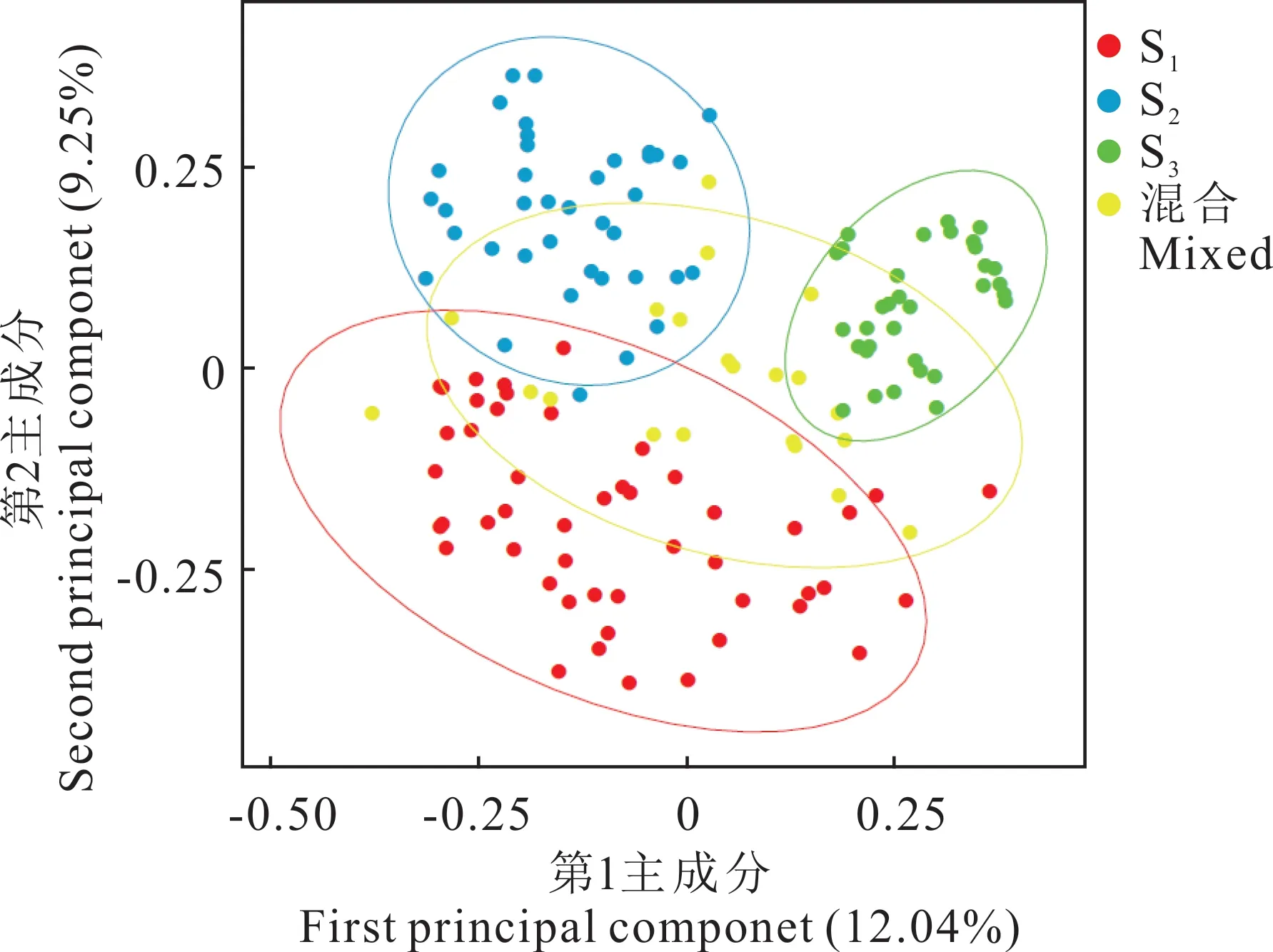
圖6 假儉草種質(zhì)資源的主坐標(biāo)分析Fig. 6 Principal coordinate analysis of the centipedegrass germplasm resources
2.5 冗余分析
冗余分析(RDA)以遺傳變量和經(jīng)過(guò)篩選的7 個(gè)環(huán)境變量(表2)分別作為響應(yīng)變量和解釋變量,結(jié)果表明,軸1 和軸2 的方差百分比分別為14.88%和4.26%(圖7)。RDA 結(jié)果解釋了遺傳變量與環(huán)境變量之間的關(guān)系(表8),各環(huán)境因子的貢獻(xiàn)度及相關(guān)性由大到小依次為最干旱月份的降水量、年平均溫、最熱月最高溫度、最干燥季平均氣溫、年降水量、溫度年變化范圍、等溫性,其中年平均溫、最熱月最高溫度、最干燥季平均氣溫、年降水量和最干旱月份的降水量與軸1 呈極顯著負(fù)相關(guān),等溫性和溫度年變化范圍與軸2 呈顯著負(fù)相關(guān),最干旱月份的降水量的貢獻(xiàn)率及相關(guān)性最高,表明最干旱月份的降水量對(duì)河南省假儉草的種群遺傳結(jié)構(gòu)影響最顯著。
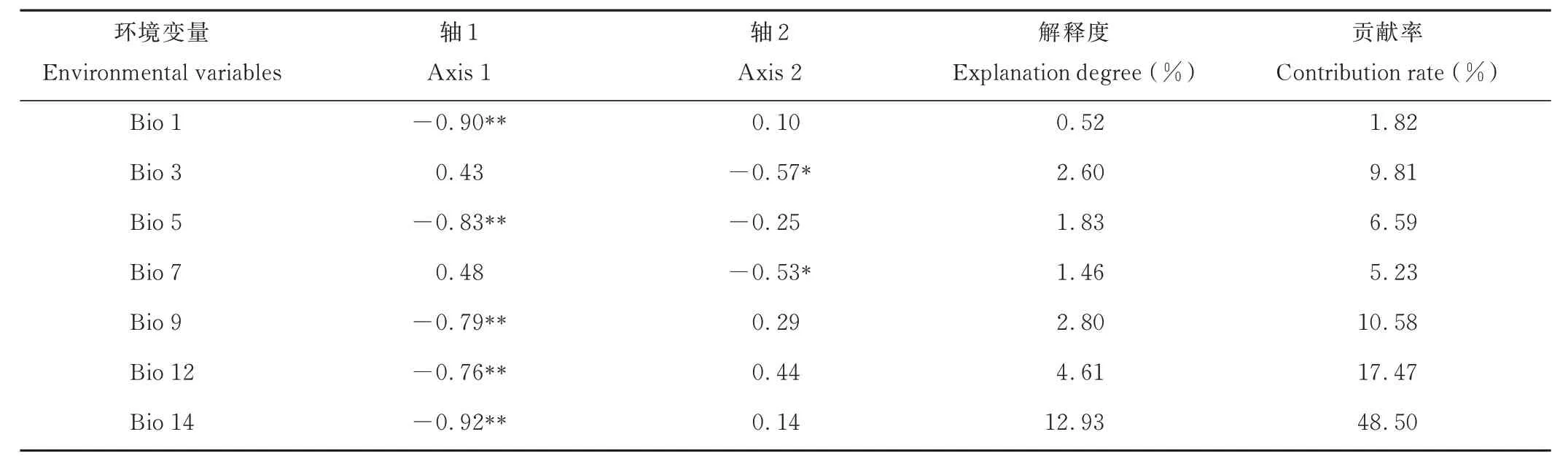
表8 各環(huán)境變量與排序軸的相關(guān)性、解釋度及貢獻(xiàn)率Table 8 Correlation, degree of explanation and contribution rate of each environmental variables to the ranking axis
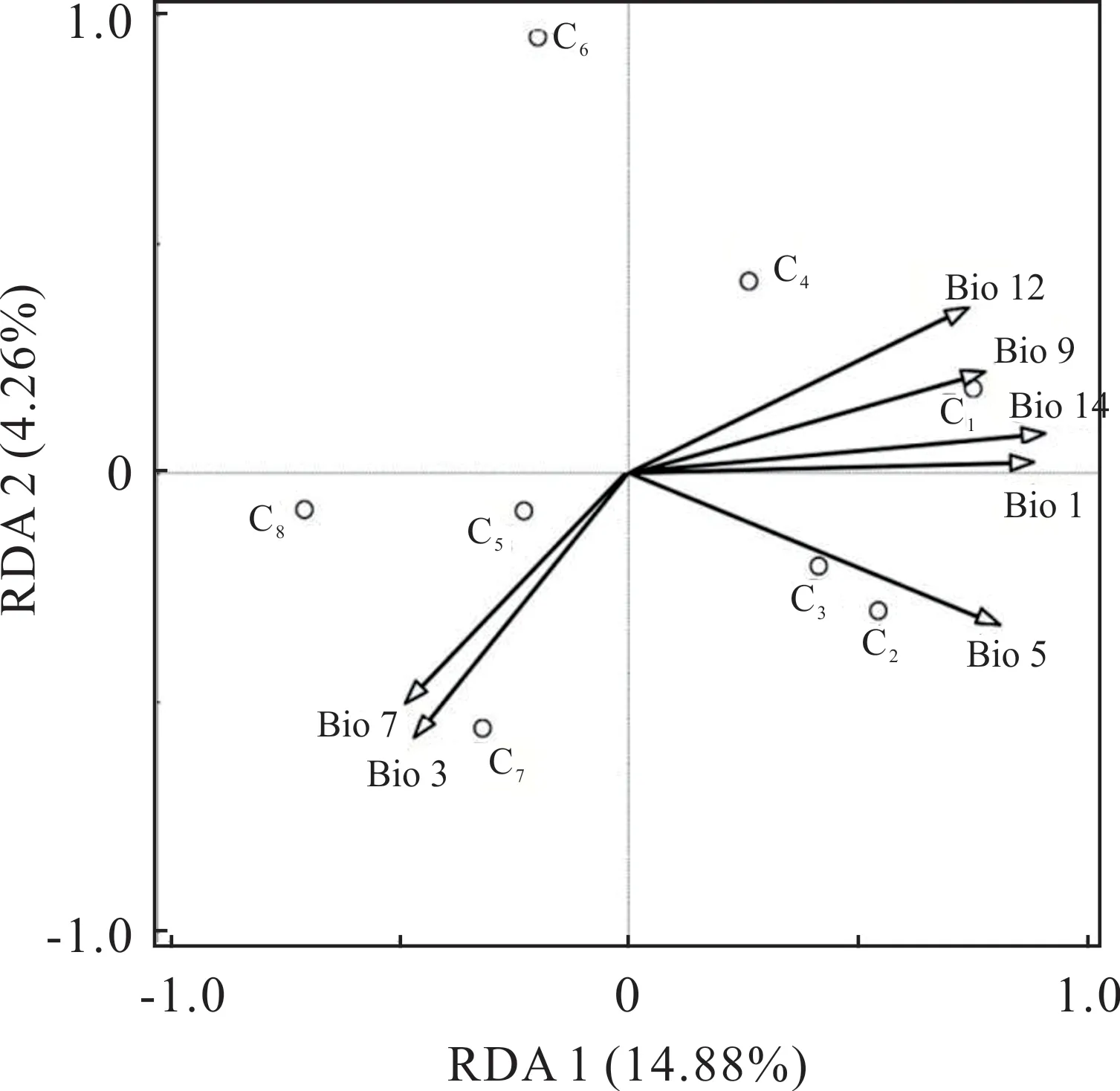
圖7 RDA 分析環(huán)境變量對(duì)遺傳結(jié)構(gòu)的相對(duì)貢獻(xiàn)Fig. 7 The relative contribution of environmental variables to genetic structure was analyzed by RDA
2.6 異常位點(diǎn)檢測(cè)及環(huán)境關(guān)聯(lián)分析
用BayeScan 方法共檢測(cè)到6 個(gè)異常位點(diǎn),占總位點(diǎn)的6.06%,引物TJIB.EO-82 檢測(cè)到3 個(gè)異常位點(diǎn),引物TJIB. EO-65 檢測(cè)到2 個(gè)異常位點(diǎn),引物TJIB.EO-29 檢測(cè)到1 個(gè)異常位點(diǎn)。利用Arlequin 的分層島模型,F(xiàn)DIST2 方法檢測(cè)到22 個(gè)異常位點(diǎn),占總位點(diǎn)的22.22%,引物TJIB. EO-51 檢測(cè)到7 個(gè)異常位點(diǎn),引物TJIB. EO-66 檢測(cè)到4 個(gè)異常位點(diǎn),引物TJIB.EO-90 和TJIB.EO-29 分別檢測(cè)到3 個(gè)異常位點(diǎn),引物TJIB. EO-31 和TJIB. EO-65 分別檢測(cè)到2 個(gè)異常位點(diǎn),引物TJIB. EO-82 檢測(cè)到1 個(gè)異常位點(diǎn)。為使結(jié)果更真實(shí)地代表離群位點(diǎn),將FDIST2 和BayeScan 兩種方法共同檢測(cè)到的異常位點(diǎn)作為最終確定的異常位點(diǎn),共有4 個(gè)異常位點(diǎn)(圖8),占總位點(diǎn)的4.04%。進(jìn)一步用R 包LEA 中的LFMM 做環(huán)境關(guān)聯(lián)分析,4 個(gè)共同檢測(cè)到的異常位點(diǎn)都至少與一個(gè)環(huán)境變量相關(guān),|Z|得分見(jiàn)表9。位點(diǎn)TJIB.EO-82-3 和TJIB.EO-65-6與1 個(gè)環(huán)境變量相關(guān),位點(diǎn)TJIB. EO-65-2 和TJIB.EO-29-6 與7 個(gè)環(huán)境變量顯著相關(guān)。等溫性(Bio 3)和溫度年變化范圍(Bio 7)相關(guān)的環(huán)境關(guān)聯(lián)位點(diǎn)數(shù)量較多。

表9 |Z|得分確定的環(huán)境關(guān)聯(lián)位點(diǎn)Table 9 Environment association site determined by the score |Z|
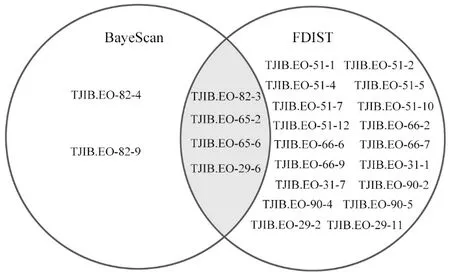
圖8 異常位點(diǎn)的檢測(cè)結(jié)果Fig.8 Detection results of outlier loci
3 討論
3.1 假儉草的遺傳多樣性
SSR 標(biāo)記具有高重復(fù)性、高穩(wěn)定性、操作簡(jiǎn)單、所需DNA 樣品量較少且質(zhì)量要求不高等優(yōu)點(diǎn),被廣泛用于分析植物的遺傳多樣性。本研究選用10 對(duì)SSR 引物對(duì)采自河南省8 個(gè)野生假儉草居群共141 份材料進(jìn)行了分析,結(jié)果表明10 對(duì)SSR 引物的平均PIC 值為0.78,Shannon’s 遺傳多樣性信息指數(shù)平均值為1.91。Li等[14]選用50 個(gè)SSR 標(biāo)記評(píng)估43 份假儉草種質(zhì)的遺傳多樣性,引物PIC 平均值為0.58,Shannon’s 遺傳多樣性信息指數(shù)平均值為1.17。與前人的研究結(jié)果相比,本研究所選的SSR 引物PIC 和Shannon’s 遺傳多樣性信息指數(shù)均較高。Susana 等[10]利用SRAP 標(biāo)記對(duì)采自我國(guó)7 個(gè)省的假儉草居群遺傳多樣性進(jìn)行了分析,結(jié)果表明供試假儉草的7 個(gè)居群中,河南省野生假儉草具有較高的遺傳多樣性,本研究結(jié)果與其一致。河南省是假儉草在華中地區(qū)分布的北緣,Thomas 等[17]認(rèn)為邊緣居群處于相對(duì)惡劣的條件下,可能面臨更大的生存壓力,使得居群中營(yíng)養(yǎng)繁殖機(jī)會(huì)減少,有性繁殖增加,從而導(dǎo)致遺傳多樣性較高。不同假儉草居群的遺傳多樣性存在差異,基于Shannon’s 遺傳多樣性信息指數(shù)居群遺傳多樣性從大到小依次為C4>C3>C6>C2>C5>C7>C1>C8。穆瑩等[46]研究表明居群遺傳多樣性大小與居群個(gè)體數(shù)量及人為干擾有關(guān)。本研究在前期踏查中發(fā)現(xiàn),C4居群位于人跡罕至的叢林中,居群內(nèi)假儉草個(gè)體數(shù)量最多,且該地受人為干擾較少,生境完整。據(jù)此推測(cè)居群個(gè)體數(shù)量多、人為干擾較少、生境完整是C4居群遺傳多樣性高的重要原因。C1居群位于水稻田的田埂上,受人為干擾影響較大,大面積水稻種植使假儉草生境遭到嚴(yán)重破壞,居群個(gè)體數(shù)量最少,居群內(nèi)的假儉草個(gè)體呈片段化分布狀態(tài),造成C1居群的遺傳多樣性較低。C8居群靠近村莊道路,受人為及動(dòng)物的干擾,無(wú)性繁殖在居群內(nèi)更占優(yōu)勢(shì),居群內(nèi)多數(shù)假儉草可能來(lái)自同一母株,因而C8居群遺傳多樣性最低。綜上所述,C4居群的假儉草種質(zhì)具有豐富的遺傳變異,適應(yīng)環(huán)境變化的能力較強(qiáng),具有較高的進(jìn)化潛力,可作為優(yōu)良種質(zhì)進(jìn)行進(jìn)一步研究。此外受城鎮(zhèn)化建設(shè)及人類活動(dòng)的影響,野生假儉草居群的生境遭到不同程度的破壞,今后應(yīng)重視假儉草種質(zhì)資源的收集、評(píng)價(jià)及保護(hù)。
3.2 假儉草居群的遺傳分化
供試假儉草居群間的遺傳分化指數(shù)(Fst)為0.14,AMOVA 分析表明河南省假儉草居群的遺傳分化主要發(fā)生于居群內(nèi)(86.96%),各居群間存在較頻繁的基因交流(Nm=1.49)。宣繼萍等[6]利用RAPD研究我國(guó)6 個(gè)省份50 份假儉草材料,結(jié)果表明假儉草居群間遺傳分化為30.39%,居群內(nèi)的遺傳分化貢獻(xiàn)率為69.61%,基因流(Nm)為1.15。Susana 等[10]利用SRAP 研究我國(guó)7 個(gè)省49 份假儉草材料,結(jié)果表明,居群間遺傳分化為6.40%,居群內(nèi)的遺傳分化為93.60%。本研究結(jié)果與上述結(jié)果基本一致,均表明假儉草居群內(nèi)遺傳分化較高,居群間基因交流較頻繁。假儉草為常異交植物,自交不親和[47]。異交種的遺傳變異多發(fā)生在居群內(nèi)導(dǎo)致其遺傳分化較高[6]。由于假儉草在河南的分布較局限,主要集中在河南東南部,人、動(dòng)物的活動(dòng),風(fēng)、水流等自然媒介的傳播導(dǎo)致不同居群間花粉、種子及匍匐莖等的交流,造成基因流水平較高。
3.3 地理隔離及基因流對(duì)假儉草群體遺傳結(jié)構(gòu)的影響
不同居群和141 份材料的聚類分析結(jié)果均表明,假儉草沒(méi)有完全按照地理來(lái)源分組,與趙瓊玲等[7]、鄭軼琦[9]、王婷[12]的研究結(jié)果一致,均表明供試材料的地理距離與遺傳距離之間無(wú)顯著相關(guān)性。種群遺傳結(jié)構(gòu)受自然選擇、地理隔離和環(huán)境隔離等多種因素的影響[48-51]。Mantel 檢驗(yàn)結(jié)果表明地理隔離不是影響假儉草種群結(jié)構(gòu)的主要因素,假儉草種群結(jié)構(gòu)的形成可能受環(huán)境的影響,因此本研究進(jìn)一步分析環(huán)境因子對(duì)假儉草遺傳變異的影響,從而揭示假儉草種群遺傳分布格局形成的原因。
通過(guò)分析種群的遺傳結(jié)構(gòu),可以直觀地表明材料之間的遺傳關(guān)系和基因交流[52]。本研究應(yīng)用STRUCTURE 軟件將141 份假儉草材料分為3 個(gè)亞群,分群結(jié)果與聚類分析及主坐標(biāo)分析結(jié)果基本一致。大部分材料(85.11%)群體遺傳背景單一,基因交流較少,21 份材料屬于遺傳背景較復(fù)雜的混合型群體,基因交流較頻繁。由于人為干擾或動(dòng)物的活動(dòng),以及風(fēng)力或水流的作用,導(dǎo)致不同居群間存在花粉、種子及匍匐莖等的交流,從而導(dǎo)致不同居群間基因交流較頻繁,最終形成混合型群體。基因流結(jié)果也表明各居群間存在較頻繁的基因交流。
3.4 環(huán)境因子對(duì)假儉草群體遺傳結(jié)構(gòu)的影響
為探究環(huán)境因子對(duì)河南省假儉草群體遺傳結(jié)構(gòu)的影響,本研究對(duì)假儉草居群的遺傳變量與環(huán)境變量進(jìn)行冗余分析,結(jié)果表明,5 個(gè)環(huán)境變量(年平均溫、最熱月最高溫度、最干燥季平均氣溫、年降水量、最干旱月份的降水量)對(duì)供試假儉草群體遺傳結(jié)構(gòu)有顯著影響,其中3 個(gè)環(huán)境變量與溫度有關(guān),2 個(gè)與降水有關(guān),最干旱月份的降水量對(duì)河南省假儉草群體遺傳結(jié)構(gòu)的影響最大。近年來(lái),國(guó)內(nèi)外學(xué)者就環(huán)境因素與植物群體遺傳結(jié)構(gòu)間的關(guān)系進(jìn)行了諸多研究,部分學(xué)者認(rèn)為溫度是影響群體遺傳結(jié)構(gòu)的主要因素,如Jiang 等[53]的研究結(jié)果表明等溫性是影響嶺南青岡(Quercus championii)群體遺傳結(jié)構(gòu)的最主要環(huán)境因素,Shryock 等[54]的研究結(jié)果表明年平均氣溫是影響麻黃(Ephedra nevadensis)和秋葵(Sphaeralcea ambigua)群體遺傳結(jié)構(gòu)的主要環(huán)境因素,Jia 等[55]認(rèn)為溫度因子是影響側(cè)柏(Platycladus orientalis)群體遺傳結(jié)構(gòu)的關(guān)鍵因子。部分研究發(fā)現(xiàn)降水是導(dǎo)致群體遺傳分化的主要因素,如Miao 等[56]認(rèn)為最暖季降水是影響黃櫨(Cotinus coggygria)群體遺傳結(jié)構(gòu)的主要因素,Zhang 等[57]認(rèn)為年降水量和最濕季降水量是影響白皮松(Pinus bungeana)群體遺傳結(jié)構(gòu)的主要因素,Yang 等[58]認(rèn)為最干旱季降水是影響連翹(Forsythia suspensa)群體遺傳結(jié)構(gòu)的主要因素。本研究中由于假儉草在河南省的分布地區(qū)較狹窄,供試的8 個(gè)采樣點(diǎn)氣候差異較小,提取的19 個(gè)氣候因子中僅有7 個(gè)不存在極顯著相關(guān)。8 個(gè)居群中C1(22 mm)、C2(24 mm)和C3居群(23 mm)的最干旱月份的降水量較多,C7(14 mm)和C8(14 mm)居群最少,而C4(21 mm)、C5(20 mm)和C6居群(18 mm)居中,群體遺傳結(jié)構(gòu)的分群結(jié)果將供試材料分為3 組,與最干旱月的降水量的分布情況一致,所以制約假儉草在華中地區(qū)北緣分布的最主要因素可能不是溫度,而是降水。在前期踏查中也發(fā)現(xiàn),河南省假儉草主要沿河流分布,生境較單一,可能是在長(zhǎng)期的自然壓力下為了彌補(bǔ)降水不足帶來(lái)的水分虧缺而選擇臨河而居。
隨著第二代測(cè)序技術(shù)的廣泛應(yīng)用,更易獲得物種基因組信息,可為研究物種適應(yīng)性基因奠定良好基礎(chǔ)[59]。目前,通過(guò)篩選環(huán)境異常位點(diǎn)進(jìn)而探究異常位點(diǎn)注釋的相應(yīng)基因功能的方法,被廣泛應(yīng)用到研究物種的局地適應(yīng)性。Li 等[16]利用簡(jiǎn)化基因組測(cè)序技術(shù)研究我國(guó)22 個(gè)楓楊(Pterocarya stenoptera)居群適應(yīng)性分化,異常位點(diǎn)檢出比例為8.10%,將LFMM 和Samβada 兩種方法共同篩選出的異常位點(diǎn)進(jìn)行功能注釋,定位到了與溫度、水分和光適應(yīng)相關(guān)的候選基因,闡釋了自然選擇對(duì)楓楊種群局地適應(yīng)的影響。Yan 等[60]利用RAD-seq 技術(shù)分析我國(guó)27 個(gè)短花針茅(Stipa breviflora)種群遺傳分化分子機(jī)制,利用Fst 異常值檢測(cè)的方法共篩選到494 個(gè)異常位點(diǎn),異常位點(diǎn)檢出比例為1.92%,28 個(gè)異常位點(diǎn)成功比對(duì)到轉(zhuǎn)錄組序列,反映出高溫、干旱、紫外線等環(huán)境因素是影響短花針茅種群遺傳分化的重要因素。本研究結(jié)合FDIST2 和BayeScan 兩種方法共同檢測(cè)到4 個(gè)異常位點(diǎn),異常位點(diǎn)檢出比例為4.04%,由于標(biāo)記技術(shù)及位點(diǎn)數(shù)量有限,本研究結(jié)果雖然檢測(cè)到部分與環(huán)境因子關(guān)聯(lián)的位點(diǎn),但需要進(jìn)一步結(jié)合第二代測(cè)序技術(shù)開(kāi)展與轉(zhuǎn)錄組學(xué)相結(jié)合的研究,進(jìn)行基因功能注釋,系統(tǒng)地揭示假儉草的適應(yīng)性進(jìn)化機(jī)制。
4 結(jié)論
本研究利用SSR 技術(shù)對(duì)采自河南省8 個(gè)野生假儉草居群共141 份材料進(jìn)行群體遺傳結(jié)構(gòu)分析。結(jié)果表明河南省假儉草資源遺傳多樣性較高,居群內(nèi)的遺傳分化較高,居群間基因交流較頻繁,揭示了河南省假儉草種群遺傳分布格局的形成主要是由最干旱月的降水量決定的。未來(lái)可結(jié)合第二代測(cè)序技術(shù)開(kāi)展與轉(zhuǎn)錄組學(xué)相結(jié)合的研究,進(jìn)行基因功能注釋,系統(tǒng)地揭示假儉草的適應(yīng)性進(jìn)化機(jī)制。
猜你喜歡
體育科技文獻(xiàn)通報(bào)(2022年3期)2022-05-23 13:46:54
天津外國(guó)語(yǔ)大學(xué)學(xué)報(bào)(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機(jī)設(shè)計(jì)與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學(xué)報(bào)(2017年2期)2017-07-05 08:13:02
國(guó)際商務(wù)財(cái)會(huì)(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
