低溫脅迫對2 個石榴品種幼苗光合參數和生理特性的影響
2023-10-04 21:23:59關思慧柴亞倩崔洪鑫宋晉泰朱光裕劉慧英郝慶刁明
果樹學報 2023年5期
關思慧 柴亞倩 崔洪鑫 宋晉泰 朱光裕 劉慧英 郝慶 刁明
摘要:【目的】研究不同低溫脅迫下2 個石榴品種光合、葉綠素熒光及生理特性的響應差異。【方法】采用盆栽方式,以1 年生喀什酸石榴和突尼斯軟籽石榴2 個品種幼苗為試驗材料,在人工氣候室中模擬低溫,設置5 個溫度水平的處理,探究低溫處理期間2 個石榴品種幼苗光合特性、葉綠素熒光參數、光合色素和滲透調節物質含量的變化,分析石榴的低溫生理響應特性。【結果】隨著脅迫溫度的下降,葉綠素a 含量、類胡蘿卜素含量、總葉綠素含量、凈光合速率(Pn)、蒸騰速率(Tr)、氣孔導度(Gs)、水分利用效率(water use efficiency,WUE)和最大光化學效率(Fv/Fm)、光化學猝滅系數(qP)、實際光化學效率[Y(Ⅱ)]均呈現下降趨勢,脯氨酸含量、可溶性糖含量、可溶性蛋白含量、相對電導率、丙二醛(malondialdehyde,MDA)含量、非光化學淬滅系數(non-photochemical quenching,NPQ)呈上升趨勢,胞間CO2濃度(Ci)呈先升后降趨勢;隨脅迫時間的延長,2 個石榴品種凈光合速率、氣孔導度、水分利用效率、Fv/Fm和[Y(Ⅱ)]呈下降趨勢,其余指標的變化趨勢均有所不同。【結論】喀什酸石榴植株抗寒性顯著強于突尼斯軟籽石榴。喀什酸石榴葉片較厚重,在低溫脅迫下抗脫水能力強,光合機構受損傷程度低,有較強維持葉綠素含量的能力,光合效率較高,同時維持細胞滲透調節能力和膜脂過氧化能力強。突尼斯軟籽石榴在重度低溫脅迫持續6 d 或極重度低溫脅迫持續4 d 后其光合系統被嚴重破壞,葉片生長被抑制,出現低溫凍害。該研究結果為寒冷地區引進軟籽石榴的適栽區劃分和防寒栽培管理技術的選擇等提供理論依據。
關鍵詞:石榴幼苗;低溫脅迫;光合;滲透調節
中圖分類號:S665.4 文獻標志碼:A 文章編號:1009-9980(2023)05-0946-13
石榴(Punica granatum L.)屬石榴科(Punicaceae)石榴屬(Punica L.)植物[1],是集生態作用與經濟價值為一體的特色果樹[2]。石榴產業是新疆特色林果業之一,目前已成為南疆鄉村振興、農民致富的支柱產業。在生產中,新疆當地種植的硬籽石榴品種盡管有很強的適應性,但存在品種較為單一、口味和宜食性等品質指標無法滿足消費者需求等問題[3]。
近年來,突尼斯軟籽石榴因籽粒柔軟、粒大多汁、品質好和可食率較高等特性具有巨大的市場潛力[4-5],已成為新疆石榴主產區更新換代的主流品種。軟籽石榴喜溫畏寒,低溫是限制其引種和分布的主要因素。在新疆南疆的石榴主產區,軟籽石榴可露地栽培,但冬季需采取防凍保護措施。早春的低溫和倒春寒往往會對軟籽石榴造成不同程度的冷害和凍害,而其中幼樹的抗寒性更弱,容易遭受低溫危害,進而抑制石榴的生長和發育,影響后期的產量和品質[6-7]。因此,在新疆寒冷地區引進軟籽石榴新品種時,研究軟籽石榴對低溫脅迫的生理響應是評價其在該地區生長適應性的主要依據之一。
光合作用是植物最基本的生理現象,是干物質積累的唯一途徑[8]。光合能力強弱與植物產量和品質密切相關[9]。低溫脅迫破壞葉綠體結構使葉綠素合成受阻,導致葉綠素含量顯著下降[10],植株黃化,葉片凈光合速率(Pn)下降[11]。強耐寒型品種植株的Pn相對較高[12];低溫脅迫會影響光能的吸收、轉換與光電子傳遞,致使過剩激發能大量積累于光合系統Ⅱ(PSⅡ)反應中心[13],進而損傷光合機構,降低最大光化學效率(Fv/Fm)、實際光化學效率(ΦPSⅡ)、電子傳遞速率(electron transport rate,ETR)和光化學淬滅系數(qP),導致葉片光合能力減弱[14]。低溫脅迫還會改變細胞膜透性,丙二醛(malondialdehyde,MDA)及滲透調節物質如脯氨酸、可溶性糖和可溶性蛋白含量增加[15],以維持細胞滲透平衡,減緩低溫帶來的損傷[16]。
目前,已有低溫脅迫對秋菊[17]、砂糖橘[18]、臍橙[19]、小麥[20]、棉花[21]等影響的相關生理研究。石榴生理方面的相關研究主要集中在枝條忍耐低溫的極限溫度上,而對于突尼斯軟籽石榴幼苗能忍受的低溫脅迫時間卻鮮見報道。在早春低溫的生產中,短暫低溫下植物可以恢復生長能力,但持續低溫會對植物造成不可逆的影響。在新疆眾多引種的軟籽石榴品種中,突尼斯軟籽石榴栽培最廣泛。為解決生產上遇到的實際問題,本研究以新疆本地的喀什酸石榴和引進的突尼斯軟籽石榴幼苗作為研究對象,采用人工模擬早春低溫方法,探究不同低溫脅迫下2個石榴品種的色素含量、光合氣體交換和葉綠素熒光參數、葉片細胞相對電導率、MDA含量及滲透調節物質含量等指標的變化差異,以期為軟籽石榴在新疆地區的合理引種和抗逆栽培管理提供理論依據。
1 材料和方法
1.1 試驗材料
供試材料為1 年生春季扦插的喀什酸石榴和突尼斯軟籽石榴幼苗(分別引自新疆喀什和河南滎陽)。試驗于石河子大學農學院試驗站溫室和兵團特色果蔬栽培生理與種質資源利用重點實驗室進行。選取生長健壯、無病蟲害和機械損傷且整齊一致的幼苗,移栽于直徑12 cm、高18.5 cm的營養袋中,袋內盛裝草炭、蛭石與珍珠巖復配基質的體積比為1∶1∶1。幼苗統一修剪,保留苗高35~40 cm,正常養護管理,待緩苗結束后轉移至人工氣候箱中進行低溫處理。
1.2 試驗設計
試驗共設置5 個溫度處理,即(1)CK:以25 ℃(晝)/20 ℃(夜)為對照;(2)T1:6 ℃(輕度低溫脅迫);(3)T2:3 ℃(中度低溫脅迫);(4)T3:0 ℃(重度低溫脅迫);(5)T4:-3 ℃(極重度低溫脅迫)。處理前,幼苗均在人工氣候箱(Percival,美國)中進行預培養,培養環境條件均為溫度25 ℃(晝)/20 ℃(夜)、光照周期16 h/8 h、光照度100 μmol ·m- 2 · s- 1、相對濕度70%~80%。然后將4 個低溫脅迫處理按3 ℃·h-1連續降溫的方式進行降溫處理,直至降至目標低溫。每個處理3次重復。于處理2、4、6 d 后測定各項指標。
1.3 測定項目及方法
1.3.1 葉綠素及類胡蘿卜素含量的測定 用95%乙醇浸泡葉片48 h,置于黑暗條件下浸提,至葉片組織完全變白,即為浸提完全,得到色素提取溶液,參照嚴衍祿等[22]方法測定并計算葉綠素a、葉綠素b、類胡蘿卜素及葉綠素總量。
1.3.2 光合參數的測定 試驗以正常25 ℃溫度處理為對照(CK)處理2、4、6 d,于低溫處理相同天數后,使用美國LI-COR公司便攜式光合儀LI-6400在上午10︰00—12︰00測定2個石榴品種幼苗功能葉片(從枝條頂部倒數第7~8 枚完全葉)的凈光合速率(Pn)、氣孔導度(Gs)、胞間CO2濃度(Ci)和蒸騰速率(Tr),并計算水分利用效率(water use efficiency,WUE)=光合速率/蒸騰速率。測定光照度為400 mol·m-2s-1,CO2濃度為400 μmol·mol-1,流速為500 μmol·s-1。每個處理3個重復,每個重復選取1株幼苗的2片功能葉。
1.3.3 葉綠素熒光參數測定 利用脈沖調制式葉綠素熒光成像系統MAX-Imaging-PAM測定葉綠素熒光動力學參數,測定時選取生長一致且受光方向相同的葉片,暗適應30 min 后測定慢速動力學曲線。石榴幼苗葉片的測定參數包括PSⅡ最大光化學效率(Fv/Fm)、實際光化學效率[Y(Ⅱ)]、光化學淬滅系數(qP)、非光化學淬滅系數(non- photochemicalquenching,NPQ)和光系統Ⅱ的絕對電子傳遞速率(Electron transport rate,ETR)。
1.3.4 相對電導率、膜脂過氧化程度和滲透調節物質含量的測定 葉片相對電導率采用電解質外滲量法[23]測定。MDA含量參考硫代巴比妥酸(TBA)顯色法[24]測定。脯氨酸含量參考磺基水楊酸比色法測定,可溶性蛋白質含量采用考馬斯亮藍法測定,可溶性糖含量采用蒽酮比色法[25]測定。
1.4 統計分析
采用Excel 2010 軟件進行數據統計與制圖,采用R 語言(Mango,英國)軟件進行數據分析(p<0.05),采用ANOVA作方差分析和采用Duncan 多重比較法進行比較分析。
2 結果與分析
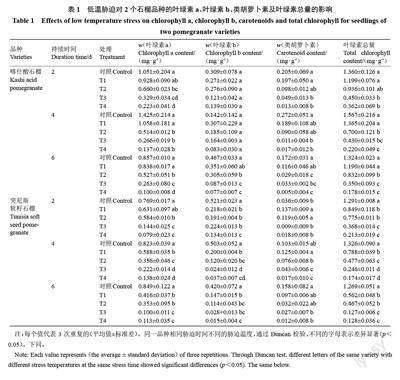
2.1 低溫脅迫對石榴葉片葉綠素含量的影響
由表1 可知,隨低溫脅迫程度的加強和脅迫時間的延長,2 個石榴品種葉片的色素含量均呈不同程度降低的趨勢。T1 處理對在整個處理期間喀什酸石榴的葉綠素a、葉綠素b、類胡蘿卜素及葉綠素總量均無顯著性影響,但顯著降低了突尼斯軟籽石榴的葉綠素a(除持續2 d 處理)、葉綠素b 及葉綠素總量。此外,T2、T3 和T4 處理均顯著降低了2 個石榴品種的葉綠素a、葉綠素b 和葉綠素總量。在同一低溫脅迫時間下,突尼斯軟籽石榴的葉綠素總量下降幅度高于喀什酸石榴,且隨脅迫程度的加劇,葉綠素總量下降幅度越高于喀什酸石榴。這說明低溫影響了葉綠素的合成與降解,使葉綠素總量降低。突尼斯軟籽石榴的色素含量對低溫的響應較喀什酸石榴敏感。
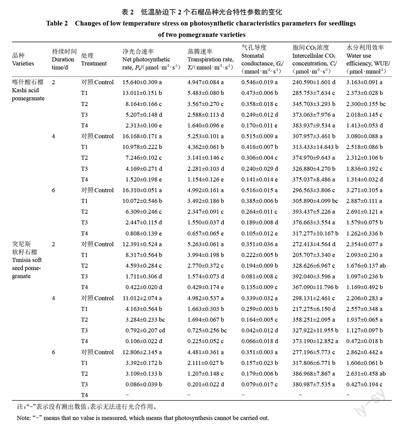
2.2 低溫脅迫對石榴光合特性的影響
如表2所示,與對照相比,相同處理時間的4個低溫處理均導致2個品種石榴葉片的Pn顯著降低,且溫度越低,Pn的降幅越大。同一低溫脅迫下,隨脅迫天數的增加,2 個石榴品種的Pn亦呈下降趨勢。其中,突尼斯軟籽石榴在極重度脅迫(T4 處理)6 d 后未檢測到光合參數,說明極重度低溫處理下時間過長會導致葉片光合功能受到破壞,無法進行光合作用;同一低溫脅迫處理下隨低溫處理時間的延長,不同低溫處理下,2 個石榴品種Tr的變化趨勢與Pn相同,且各處理間差異顯著。突尼斯軟籽石榴Tr的下降幅度大于喀什酸石榴;Ci總體結果上顯示隨溫度下降先升高后降低,同一低溫條件下隨脅迫時間的延長呈升高趨勢;同一脅迫時間不同低溫處理下2個品種石榴葉片的胞間CO2濃度與對照的差異隨處理溫度的降低而呈顯著負相關。WUE是植物生產中單位水分的能量轉化率指標[26],同一脅迫時間隨溫度降低喀什酸石榴的水分利用效率顯著下降,突尼斯軟籽石榴各處理之間變化不顯著。這說明2 個石榴品種的光合氣體交換參數都受溫度和時間互作的影響,且對突尼斯軟籽石榴的抑制效應更大。
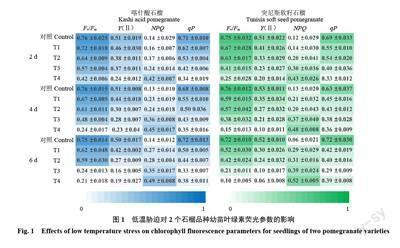
2.3 低溫脅迫對葉綠素熒光參數的影響
由圖1 可知,2 個品種石榴葉片的Fv/Fm隨溫度處理的降低和脅迫時間的延長均呈下降趨勢,其中以突尼斯軟籽石榴的降幅較大。此外,圖2 的Fv/Fm熒光圖像結果顯示,低溫處理下2 個品種石榴葉片藍色面積減少,綠色和紅棕的面積增加的趨勢與圖1 中Fv/Fm值的變化基本一致。在低溫脅迫6 d 后的T3 和T4 處理下,可以明顯看到突尼斯軟籽石榴的芽頭萎縮,葉片外圍失水萎蔫、出現斑塊,顏色變黃。這說明低溫脅迫損害了2 個品種石榴葉片的光系統Ⅱ(PSⅡ),導致光抑制發生,尤其對突尼斯軟籽石榴的光合能力抑制更大。在極重度低溫和脅迫時間過長下,該品種受到了嚴重且不可恢復的損傷。
隨著溫度的降低和脅迫時間的延長,2 個石榴品種的Y(Ⅱ)和qP 均呈下降趨勢,說明該情況下石榴葉片的光化學效率及電子傳遞速率下降;此外,由圖1 可知,2 個石榴品種葉片的NPQ 與低溫脅迫持續時間關系不大,但隨著低溫強度加劇呈上升趨勢;溫度越低,NPQ下降幅度越大,說明低溫脅迫使2 個品種石榴葉片PSⅡ反應中心潛在熱耗能力增強,從而避免植物因吸收過多的光能而造成傷害,NPQ的升高也有可能源于類囊體質子梯度差的增加和葉黃素氧化程度的增強。
2.4 低溫脅迫對2 個石榴品種幼苗滲透調節物質含量的影響
2.4.1 脯氨酸含量 如圖3 所示,與CK相比,處理2 d 后,T3 和T4 處理下喀什酸石榴的脯氨酸含量分別顯著增加了107.71%和215.75%,而T1 和T2 處理下的脯氨酸含量無顯著性變化。脅迫處理4 d 后,4個低溫脅迫處理的脯氨酸含量均較對照顯著增加,分別增加了40.77%、117.8%、156.55%和201.66%。脅迫處理6 d 后,T2、T3 和T4 處理的脯氨酸含量較對照分別顯著增加了126.57%、201.1%、226.49%;4個低溫脅迫處理2、4、6 d 后,突尼斯軟籽石榴葉片的脯氨酸含量均較對照顯著提高,分別增加了127.84% ~173.27% 、212.3% ~324.79% 、252.71% ~373.19%和329.92%~398.75%。2 個品種相比,低溫處理過程中脯氨酸含量均隨脅迫時間延長和脅迫程度加劇而呈現上升趨勢。其中,突尼斯軟籽石榴品種脯氨酸含量的增幅大于喀什酸石榴品種。
2.4.2 可溶性糖含量 由圖4可知,喀什酸石榴在處理2、4、6 d 后各處理的可溶性糖含量較對照分別顯著增加21.6%~57.93%、34.05%~132.56%、17.65%~47.28%,雖然脅迫6 d后的T3、T4處理可溶性糖含量與T2 處理相比有下降趨勢,但與對照相比可溶性糖含量仍有所增加;同一處理時間突尼斯軟籽石榴隨低溫脅迫程度提高呈先升高后降低的趨勢,脅迫處理2、4、6 d 后4 個低溫脅迫處理的可溶性糖含量較對照分別顯著升高了14.72%~48.7% 、39.65%~93.02%、46.92%~71.74%。從整體來看,2 個石榴品種葉片中可溶性糖含量在不同低溫脅迫處理4 d 時的增幅均表現最大。
2.4.3 可溶性蛋白質含量 如圖5 所示,隨著處理溫度的下降,2 個石榴品種葉片的可溶性蛋白質含量均表現出升高的趨勢。喀什酸石榴在脅迫2、4、6 d后,可溶性蛋白質含量(w,后同)分別在1.887~3.425、2.485~4.486 和2.734~4.167 mg· g-1,其中各處理較對照分別增加56.21%~183.53%、110.95%~280.81%和125.96%~244.38%,均與對照呈顯著差異。突尼斯軟籽石榴在低溫脅迫2、4、6 d 后可溶性蛋白質含量在2.826~6.120、4.649~6.762 和3.292~6.568 mg·g-1之間,分別較對比組顯著增加72.74%~274.08% 、180.74%~308.33%和92.51%~284.09% 。從脅迫時間看,2 個品種石榴葉片可溶性蛋白質含量隨脅迫時間的延長,大致表現為先上升再下降的趨勢。其中,突尼斯軟籽石榴品種可溶性蛋白質含量的增幅明顯大于喀什酸石榴品種,且突尼斯軟籽石榴在低溫脅迫后整體可溶性蛋白質含量比喀什酸石榴高。
2.5 低溫脅迫對2 個石榴品種幼苗電解質滲透率和MDA含量的影響
2.5.1 電解質滲透率 由圖6 可知,喀什酸石榴在脅迫2 d 后T1 與對照相比升高7.19%,無顯著差異(p>0.05),在T2、T3、T4 處理后與對照相比分別升高287.96%、364.33%、446.72%,與對照差異顯著;脅迫4 d 后,與對照相比分別顯著升高102.23%、238.25%、298.48%、468.72%;脅迫6 d 后與對照相比分別增加42.85%、78.62%、259.08%、469.86,除T1 處理無顯著性變化外,與其余處理均呈顯著差異。突尼斯軟籽石榴隨低溫脅迫相對電導率也隨之上升,脅迫2、4、6 d 后分別較對照上升87.83%~406.93%、51.43%~388.62%、102.23%~560.14%,在T3 處理的相對電導率較前一處理有所下降,但與對照相比仍是上升的,且除脅迫6 d 后T3 處理外各個處理均與對照相比達到顯著差異。隨溫度的降低,2 個品種石榴葉片相對電導率呈升高趨勢。其中,突尼斯軟籽石榴品種相對電導率在T4 處理時脅迫4、6 d 后的升幅激增,明顯大于喀什酸石榴品種。
2.5.2 MDA含量 如圖7 所示,喀什酸石榴在脅迫2 d 后MDA含量(w,后同)在0.496~0.855 mg· g-1之間,T1、T2 處理時較對照上升9.27%和9.07%,無顯著性差異(p>0.05),T3、T4處理的分別上升75.2%和72.37%,與對照相比差異顯著;脅迫4、6 d后MDA含量分別在0.669~0.973、0.637~1.04 mg·g-1之間,增幅分別為45.43%~111.52%和23.93%~102.33%,除T1處理的脅迫6 d 外其余各處理均與對照差異顯著。突尼斯軟籽石榴與對照相比,處理2、4、6 d 后MDA含量分別在0.552~1.073、0.539~1.403 和1.229~1.198 4 mg·g-1之間,在脅迫2 d 后的T3、T4 處理、脅迫4 d 后的T2、T3、T4 處理和脅迫6 d 后的各處理均與對照呈顯著差異。低溫處理過程中,2 個石榴品種處理T1、T2 和T3 脅迫2 d 的MDA含量無顯著變化,在低溫各處理脅迫4、6 d 后均隨脅迫時間延長和脅迫程度加劇而呈現出上升趨勢。其中,突尼斯軟籽石榴品種MDA 含量的增幅大于喀什酸石榴品種,尤其在脅迫4 d 時T2、T3 和T4 處理和脅迫6 d 時的各處理,MDA含量急劇升高。
3 討論
低溫是植物生長、發育和生存的環境脅迫因子之一,影響植物的光合活性和生理生化過程。光合作用是植物生長最根本的基礎,可為植物提供所需的能量和有機物質。其中,光合參數可直接反映植株長勢和抗性強弱,與生理指標一樣都是評估低溫脅迫程度和植物耐寒能力的可靠依據[27-28]。本研究中,低溫導致2 個石榴品種葉片光合參數受到顯著抑制,隨著低溫脅迫時間的延長Pn、Gs以及Tr持續下降,說明低溫抑制了植物的光合作用,且隨著脅迫時間的延長和脅迫程度的加強而呈加重趨勢。此外,不同低溫處理對突尼斯軟籽石榴的抑制效應更大。突尼斯軟籽石榴在極重度低溫脅迫6 d 后光合參數測不出數值,同時重度低溫脅迫6 d 和極重度低溫脅迫4 d 凈光合速率趨近于0,說明突尼斯軟籽石榴品種較喀什酸石榴品種的低溫耐性弱,在重度低溫和極重度低溫脅迫環境下突尼斯軟籽石榴品種幾乎無光合產物積累。大多數研究學者認為,低溫導致光合速率下降的因素既有氣孔因素也有非氣孔因素。如果Pn、Gs及Tr同時下降,Ci也相應下降,即氣孔因素占主導;如果Pn、Gs及Tr下降的同時Ci升高,則是由非氣孔因素阻礙CO2的利用,造成Ci積累所致。本試驗中,低溫條件下Ci隨溫度降低呈先上升后下降趨勢,表明在低溫脅迫下,石榴葉片Pn下降主要是由非氣孔限制為主作用的結果。這與馮立娟等[29]等在設施條件下測得突尼斯軟籽石榴光合參數的結論一致。
光合色素具有吸收、傳遞和轉化光能的作用,葉綠素含量常作為葉綠體發育、光合能力的指標[30],與光合速率呈正相關,類胡蘿卜素可以起到光能捕獲和光破壞防御的作用[31]。本研究中隨著處理溫度的下降和處理時間的延長,2 個品種葉片葉綠素a 和葉綠素總量較CK相比均顯著降低,喀什酸石榴在輕度脅迫時與對照相比無顯著變化,突尼斯軟籽石榴的葉綠素含量對低溫更加敏感。一方面可能是葉綠素合成酶活性降低使葉綠素合成減少導致光合性能的降低,另一方面可能是過度低溫導致葉綠體超微結構受到破壞,PSⅡ的電子傳遞受阻,位于類囊體上的色素復合體受到影響從而導致葉綠素含量的降低[32]。類胡蘿卜素含量降低表明低溫脅迫下石榴光破壞防御能力的降低,進一步損傷光合機構導致光合能力下降,影響植株生長速度。葉綠素a、葉綠素b 和葉綠素總量與低溫脅迫程度和低溫脅迫時間呈負相關,這與李琦瑤等[33]在低溫脅迫條件下烤煙葉綠素含量變化趨勢的研究結論一致。
葉綠素熒光分析技術是光合作用的靈敏探針,能檢測植物光合生理狀況,反映光能的吸收、傳遞與分配情況,是闡明植物光合機制的重要手段[34]。可變熒光與最大熒光的比值Fv/Fm反映了PSⅡ反應中心的最大光化學效率,Y(Ⅱ)表示實際光化學效率,常用來表示植物光合作用中電子傳遞的量子產額。
低溫脅迫下Fv/Fm明顯下降,可作為低溫脅迫程度的指針和探針[35]。本研究中隨著溫度的降低,2 個品種葉片Fv/Fm和Y(Ⅱ)的數值均呈下降趨勢,不耐寒品種突尼斯軟籽石榴在低溫下葉綠素熒光數值變化更大,可能是低溫脅迫下葉片同化CO2的能力下降,致使葉綠體中ATP 和NADPH的含量下降,引發對PSⅡ反饋氧化還原作用的發生,進而導致光能過剩,引起Fv/Fm下降,同時說明PSⅡ的結構受到低溫脅迫的傷害,產生了光抑制。光化學猝滅系數qP是光化學反應所引起的熒光產額的下降,反映PSⅡ原初電子受體QA的還原狀態及開放的反應中心占總PSⅡ反應中心的比例[36]。本研究中石榴葉片qP 隨低溫持續時間的延長和溫度的降低均呈下降趨勢,這與楊柳青等[37]和王兆等[38]對南美天胡荽和彩葉草葉的研究結論一致,說明PSⅡ開放的反應中心比例降低和固定CO2的電子減少。NPQ 是指PSⅡ天線色素吸收的光能不能用于光合電子傳遞而以熱能的形式耗散掉的光能部分,是一種自我保護機制,對光合系統起一定的保護作用[21]。本研究中,石榴葉片的NPQ與低溫持續時間長短關系不大,但隨著低溫強度加劇NPQ呈較為顯著的上升趨勢,表明植物在受到低溫脅迫時開啟自我保護機制,植物可能通過NPQ的升高來抵御過量光能的傷害,同時部分失活的PSⅡ可能仍在啟動熱耗散的保護機制來抵御過量光能的傷害,這一研究結果與劉蕾蕾等[39]對小麥的研究結果一致。
在植物受到低溫脅迫時體內積累大量滲透調節物質來使細胞膜滲透勢下降從而減輕傷害,可溶性糖、可溶性蛋白和脯氨酸是具有提供能量、提高細胞質濃度和信號轉導的3 種重要調節物質[40]。本研究中,脯氨酸含量隨低溫脅迫時間和低溫的降低均逐漸增加,而可溶性糖和可溶性蛋白含量呈先升后降的趨勢,與謝小玉等[41]的研究結果一致,說明石榴葉片可在一定程度上通過積累滲透調節物質來緩解低溫脅迫。
在低溫脅迫下,植物需要維持細胞的行為和活性,特別是維持具有生物活性的細胞膜和蛋白質結構的穩定性,才能在不利的環境中生存[42]。因此,在細胞水平上的低溫應激往往伴隨著脫水應激,從而導致電解質泄漏的增加和膜脂相的變化。在極端情況下,冰晶可以穿透植物細胞,導致細胞質流出,最終植物死亡。本研究中,隨著低溫時間的延長和脅迫程度的加強,2 個品種石榴的相對電導率均顯著升高,說明低溫導致細胞質膜的穩定性受到一定的破壞,突尼斯軟籽石榴在極重度脅迫4、6 d 后急劇上升,說明低溫持續時間對相對電導率的影響更大。MDA也被認為是植物氧化應激和細胞膜對低溫反應的有機統一性的可靠標志[43]。本研究中,2 個石榴的MDA含量都隨低溫加劇而升高,但脅迫2 d 時,輕度脅迫和重度脅迫對喀什酸石榴無顯著影響,之后隨脅迫時間的延長喀什酸石榴表現出升高趨勢,而突尼斯軟籽石榴是先升高后降低,表明膜脂過氧化程度較嚴重。
4 結論
突尼斯軟籽石榴的低溫耐性顯著低于新疆當地主栽品種喀什酸石榴,具體表現突尼斯軟籽石榴在4 個低溫脅迫處理下電解質滲透率及MDA、可溶性糖、可溶性蛋白和脯氨酸含量均不同程度高于喀什酸石榴。此外,光合色素含量、光合參數和葉綠素熒光參數的降幅亦不同程度高于喀什酸石榴。且在該試驗條件下,突尼斯軟籽石榴不耐較長低溫持續時間的脅迫,重度低溫(0 ℃持續6 d)和極重度低溫(-3 ℃持續4 d)均導致光合作用嚴重受阻,引起低溫凍害,研究結果可為實際生產提供參考。
參考文獻References:
[1] JALIKOP S H,VENUGOPALAN R,KUMAR R. Association
of fruit traits and aril browning in pomegranate (Punica granatum
L.)[J]. Euphytica,2010,174(1):137-141.
[2] TEIXEIRA DA SILVA J A,RANA T S,NARZARY D,VERMA
N,MESHRAM D T,RANADE S A. Pomegranate biology and
biotechnology:A review[J]. Scientia Horticulturae,2013,160:
85-107.
[3] 卓熱木·塔西,木合塔爾·扎熱,盧明艷,帕麗旦·阿布什,吳正
保. 自然干旱條件下2 個石榴品種生長和光合等生理特性的
變化[J]. 干旱地區農業研究,2018,36(6):77-85.
Zhuoremu · Tashi,Muhertar · Zhare,LU Mingyan,Palidan · Abbosh,
WU Zhengbao. Changes in growth and photosynthetic
and other physiological characteristics of two pomegranate cultivars
under drought condition[J]. Agricultural Research in the Arid
Areas,2018,36(6):77-85.
[4] 胡青霞,馮夢晨,陳延惠,郭強,簡在海,史江莉,萬然,鄭先波.
突尼斯軟籽石榴果實生長發育及其品質形成規律研究[J]. 果樹
學報,2022,39(3):426-438.
HU Qingxia,FENG Mengchen,CHEN Yanhui,GUO Qiang,
JIAN Zaihai,SHI Jiangli,WAN Ran,ZHENG Xianbo. Growth
and development of Tunisia soft seed pomegranate and its quality
formation regulation[J]. Journal of Fruit Science,2022,39(3):
426-438.
[5] 柴亞倩,關思慧,崔洪鑫,許靜,朱曉玲,刁明,孔秋生. 水氮互
作對石榴幼苗光合熒光及生理特性的影響[J]. 果樹學報,
2022,39(12):2352-2364.
CHAI Yaqian,GUAN Sihui,CUI Hongxin,XU Jing,ZHU
Xiaoling,DIAO Ming,KONG Qiusheng. Effects of water and
nitrogen interaction on photosynthetic fluorescence and physiological
characteristics of pomegranate seedlings[J]. Journal of
Fruit Science,2022,39(12):2352-2364.
[6] 沈曉燕,揭波,李爽,王幸,李文杰. 倒春寒危害軟籽石榴樹的
溫度及受害情況研究[J]. 落葉果樹,2021,53(2):22-25.
SHEN Xiaoyan,JIE Bo,LI Shuang,WANG Xing,LI Wenjie. Investigation
and research on the effect of late spring coldness on
the growth of soft seed pomegranate trees[J]. Deciduous Fruits,
2021,53(2):22-25.
[7] MORADI S,BANINASAB B,GHOLAMI M,GHOBADI C.
Paclobutrazol application enhances antioxidant enzyme activities
in pomegranate plants affected by cold stress[J]. The Journal
of Horticultural Science and Biotechnology,2017,92(1):65-71.
[8] ZHANG C J,CHEN G X,GAO X X,CHU C J. Photosynthetic
decline in flag leaves of two field-grown spring wheat cultivars
with different senescence properties[J]. South African Journal of
Botany,2006,72(1):15-23.
[9] 杜祥備,王家寶,劉小平,夏家平,韓楊. 減氮運籌對甘薯光合
作用和葉綠素熒光特性的影響[J]. 應用生態學報,2019,30
(4):1253-1260.
DU Xiangbei,WANG Jiabao,LIU Xiaoping,XIA Jiaping,HAN
Yang. Effects of nitrogen fertilizer reduction management on
photosynthesis and chlorophyll fluorescence characteristics of
sweetpotato[J]. Chinese Journal of Applied Ecology,2019,30
(4):1253-1260.
[10] 許永安. 低溫脅迫對甜瓜幼苗光合能力及葉綠素熒光參數的
影響[J]. 中國瓜菜,2020,33(2):22-26.
XU Yongan. Effects of low-temperature stress on photosynthetic
capacity and chlorophyll fluorescence parameters in melon[J].
China Cucurbits and Vegetables,2020,33(2):22-26.
[11] 鄭春芳,劉偉成,魏龍,陳繼濃,張呈念,仇建標,丁文勇,鄭青
松. 外施褪黑素對低溫脅迫下紅樹植物秋茄光合作用和抗壞
血酸-谷胱甘肽循環的調控[J]. 植物生理學報,2019,55(8):
1211-1221.
ZHENG Chunfang,LIU Weicheng,WEI Long,CHEN Jinong,
ZHANG Chengnian,QIU Jianbiao,DING Wenyong,ZHENG
Qingsong. Melatonin regulates photosynthesis and ascorbateglutathione
cycle in a mangrove Kandelia obovata under low
temperature stress[J]. Plant Physiology Journal,2019,55(8):
1211-1221.
[12] 鄒婭,米文博,徐明霞,徐春梅,劉自剛,趙彩霞,米超. 低溫脅
迫下北方強冬性區甘藍型冬油菜的低溫光合生理特征[J]. 甘
肅農業大學學報,2021,56(2):105-113.
ZOU Ya,MI Wenbo,XU Mingxia,XU Chunmei,LIU Zigang,
ZHAO Caixia,MI Chao. Physiological and photosynthetic characteristics
of winter rapeseeds under low temperature stress in
northern strong winterness region[J]. Journal of Gansu Agricultural
University,2021,56(2):105-113.
[13] 羅鑫輝,劉明月,黃科,劉玉兵,彭淼,王軍偉. 不同品種番茄幼
苗低溫適應性評價及光合特性分析[J]. 中國瓜菜,2021,34
(8):49-55.
LUO Xinhui,LIU Mingyue,HUANG Ke,LIU Yubing,PENG
Miao,WANG Junwei. Evaluation of low temperature adaptability
and analysis of photosynthetic characteristics in different tomato
varieties[J]. China Cucurbits and Vegetables,2021,34(8):
49-55.
[14] 胡晨曦,李子恒,張云虹,張林巧,祁建波,張瑛,周如美,張永
泰,張永吉. 低溫弱光對不同品種辣椒幼苗生長和光合特性
的影響[J]. 福建農業學報,2022,37(5):617-625.
HU Chenxi,LI Ziheng,ZHANG Yunhong,ZHANG Linqiao,QI
Jianbo,ZHANG Ying,ZHOU Rumei,ZHANG Yongtai,
ZHANG Yongji. Effects of low temperature and reduced light
on growth and photosynthesis of seedlings of different pepper
cultivars[J]. Fujian Journal of Agricultural Sciences,2022,37
(5):617-625.
[15] 楊鴻基,徐誠,高亞寧,楊建超,楊平,軒正英,阿依買木·沙吾
提. 新疆哈密瓜苗期耐冷性評價[J]. 中國瓜菜,2022,35(2):
90-94.
YANG Hongji,XU Cheng,GAO Yaning,YANG Jianchao,
YANG Ping,XUAN Zhengying,Ayimaimu · Shawuti. Cold tolerance
evaluation of Hami melon seedlings in Xinjiang[J]. China
Cucurbits and Vegetables,2022,35(2):90-94.
[16] BAN Q Y,WANG X W,PAN C,WANG Y W,KONG L,JIANG
H G,XU Y Q,WANG W Z,PAN Y T,LI Y Y,JIANG C
J. Comparative analysis of the response and gene regulation in
cold resistant and susceptible tea plants[J]. PLoS One,2017,12
(12):e0188514.
[17] 逯久幸,苗潤田,王司琦,趙鵬飛,張開明,李永華. 低溫脅迫
下秋菊葉片光系統特性分析[J]. 植物生理學報,2022,58(2):
425-434.
LU Jiuxing,MIAO Runtian,WANG Siqi,ZHAO Pengfei,
ZHANG Kaiming,LI Yonghua. Analysis of photosystem features
in autumn chrysanthemum leaves under low temperature
stress[J]. Plant Physiology Journal,2022,58(2):425-434.
[18] 郭雁君,吉前華,杜鵬飛,黃火金,許冰玲. 低溫脅迫對2 種主
要砧穗組合砂糖橘幼樹抗寒性的影響[J]. 果樹學報,2022,39
(5):784-799.
GUO Yanjun,JI Qianhua,DU Pengfei,HUANG Huojin,XU
Bingling. Effect of two main rootstock species on cold resistance
of Shatangju mandarin saplings under low temperature
stress[J]. Journal of Fruit Science,2022,39(5):784-799.
[19] 楊軍,黃淑娥,郭建平,李翔翔,李亞靖,丁美萍. 不同夜間低
溫條件對臍橙光合參數和形態特征的影響[J]. 果樹學報,
2022,39(11):2088-2098.
YANG Jun,HUANG Shue,GUO Jianping,LI Xiangxiang,LI
Yajing,DING Meiping. Effects of different low night temperature
conditions on the photosynthetic parameters and morphological
characteristics in navel orange[J]. Journal of Fruit Science,
2022,39(11):2088-2098.
[20] 王洋洋,賀利,任德超,段劍釗,胡新,劉萬代,郭天財,王永
華,馮偉. 基于主成分-聚類分析的不同水分冬小麥晚霜凍害
評價[J]. 作物學報,2022,48(2):448-462.
WANG Yangyang,HE Li,REN Dechao,DUAN Jianzhao,HU
Xin,LIU Wandai,GUO Tiancai,WANG Yonghua,FENG Wei.
Evaluations of winter wheat late frost damage under different
water based on principal component-cluster analysis[J]. Acta Agronomica
Sinica,2022,48(2):448-462.
[21] 張隴艷,程功敏,魏恒玲,王寒濤,蘆建華,馬峙英,喻樹迅. 陸
地棉種子萌發期對低溫脅迫的響應及耐冷性鑒定[J]. 中國農
業科學,2021,54(1):19-33.
ZHANG Longyan,CHENG Gongmin,WEI Hengling,WANG
Hantao,LU Jianhua,MA Zhiying,YU Shuxun. Chilling tolerance
identification and response to cold stress of Gossypium hirsutum
varieties (lines) during germination stage[J]. Scientia Agricultura
Sinica,2021,54(1):19-33.
[22] 嚴衍祿,劉心生. 葉綠素測定方法的研究[J]. 北京農業大學學
報,1982,8(2):53-67.
YAN Yanlu,LIU Xinsheng. Studies on determination chlorophylls
a and b[J]. Journal of China Agricultural University,
1982,8(2):53-67.
[23] 鄒琦. 植物生理學實驗指導[M]. 北京:中國農業出版社,2000.
ZOU Qi. Experimental instruction of plant physiology[M]. Beijing:
China Agriculture Press,2003.
[24] 李合生. 植物生理生化實驗原理和技術[M]. 北京:高等教育
出版社,2000.
LI Hesheng. Principles and techniques of plant physiological biochemical
experiment[M]. Beijing:Higher Education Press,2000.
[25] 高俊鳳. 植物生理學實驗指導[M]. 北京:高等教育出版社,
2006.
GAO Junfeng. Experimental guidance for plant physiology[M].
Beijing:Higher Education Press,2006.
[26] 鐘信念,梁其干,李有忠,趙建華,馬海霞,李志博. 蕾期低溫脅
迫對棉花光合特性的影響[J]. 新疆農業科學,2022,59(7):
1573-1581.
ZHONG Xinnian,LIANG Qigan,LI Youzhong,ZHAO Jianhua,
MA Haixia,LI Zhibo. Effects of low temperature stress in bud
stage on photosynthetic characteristics of cotton[J]. Xinjiang Agricultural
Sciences,2022,59(7):1573-1581.
[27] 項洪濤,鄭殿峰,何寧,李琬,王曼力,王詩雅. 植物對低溫脅迫
的生理響應及外源脫落酸緩解脅迫效應的研究進展[J]. 草業
學報,2021,30(1):208-219.
XIANG Hongtao,ZHENG Dianfeng,HE Ning,LI Wan,WANG
Manli,WANG Shiya. Research progress on the physiological response
of plants to low temperature and the amelioration effcectiveness
of exogenous ABA[J]. Acta Prataculturae Sinica,2021,
30(1):208-219.
[28] GOHARRIZI K J,MERU G,KERMANI S G,HEIDARINEZHAD
A. Short- term cold stress affects physiological and biochemical
traits of pistachio rootstocks[J]. South African Journal
of Botany,2021,141:90-98.
[29] 馮立娟,楊雪梅,王嘉艷,史作亞,尹燕雷. 設施條件下兩個石
榴品種光合熒光特征參數差異與變化[J/OL]. 分子植物育種,
2021:1-13[2022-09-28]. http://kns.cnki.net/kcms/detail/46.1068.
s.20210918.1620.022.html.
FENG Lijuan,YANG Xuemei,WANG Jiayan,SHI Zuoya,
YIN Yanlei. Differences and changes of photosynthetic fluorescence
characteristic parameters of two pomegranate varieties
under protected conditions[J/OL]. Molecular Plant Breeding,
2021:1-13[2022-09-28]. http://kns.cnki.net/kcms/detail/46.1068.
s.20210918.1620.022.html.
[30] LING Q H,HUANG W H,JARVIS P. Use of a SPAD-502 meter
to measure leaf chlorophyll concentration in Arabidopsis thaliana[
J]. Photosynthesis Research,2011,107(2):209-214.
[31] 張振花,孫勝,劉洋,李靖,袁宏霞,鄭金英,邢國明. 增施CO2
對溫室番茄結果期葉片光合特性的影響[J]. 生態學雜志,
2018,37(5):1398-1402.
ZHANG Zhenhua,SUN Sheng,LIU Yang,LI Jing,YUAN
Hongxia,ZHENG Jinying,XING Guoming. Effects of CO2 enrichment
on photosynthetic characteristics of greenhouse tomato
during fruiting stage[J]. Chinese Journal of Ecology,2018,37
(5):1398-1402.
[32] 岳海,李國華,李國偉,陳麗蘭,孔廣紅,梁國平. 澳洲堅果不
同品種耐寒特性的研究[J]. 園藝學報,2010,37(1):31-38.
YUE Hai,LI Guohua,LI Guowei,CHEN Lilan,KONG Guanghong,
LIANG Guoping. Studies on cold resistance of different
Macadamia cultivars[J]. Acta Horticulturae Sinica,2010,37(1):
31-38.
[33] 李琦瑤,陳愛國,王程棟,劉光亮,周培祿,鄭璇,曾文龍,陳志
厚,王樹聲. 低溫脅迫對烤煙幼苗光合熒光特性及葉片結構
的影響[J]. 中國煙草學報,2018,24(2):30-38.
LI Qiyao,CHEN Aiguo,WANG Chengdong,LIU Guangliang,
ZHOU Peilu,ZHENG Xuan,ZENG Wenlong,CHEN Zhihou,
WANG Shusheng. Effects of low temperature stress on photosynthetic
fluorescence characteristics and leaf tissue structure of
flue-cured tobacco seedlings[J]. Acta Tabacaria Sinica,2018,24
(2):30-38.
[34] 張利霞,常青山,薛嫻,劉偉,張巧明,陳蘇丹,鄭軼琦,李景林,
陳婉東,李大釗. 酸脅迫對夏枯草葉綠素熒光特性和根系抗
氧化酶活性的影響[J]. 草業學報,2020,29(8):134-142.
ZHANG Lixia,CHANG Qingshan,XUE Xian,LIU Wei,
ZHANG Qiaoming,CHEN Sudan,ZHENG Yiqi,LI Jinglin,
CHEN Wandong,LI Dazhao. Effects of acid stress on chlorophyll
fluorescence characteristics and root antioxidant activity
of Prunella vulgaris[J]. Acta Prataculturae Sinica,2020,29(8):
134-142.
[35] 邵怡若,許建新,薛立,張柔,吳彩瓊,盧廣超. 低溫脅迫時間
對4 種幼苗生理生化及光合特性的影響[J]. 生態學報,2013,
33(14):4237-4247.
SHAO Yiruo,XU Jianxin,XUE Li,ZHANG Rou,WU Caiqiong,
LU Guangchao. Effects of low temperature stress on physiological-
biochemical indexes and photosynthetic characteristics of
seedlings of four plant species[J]. Acta Ecologica Sinica,2013,
33(14):4237-4247.
[36] CONSTANT S,PEREWOSKA I,ALFONSO M,KIRILOVSKY
D. Expression of the psbA gene during photoinhibition and recovery
in Synechocystis PCC 6714:Inhibition and damage of
transcriptional and translational machinery prevent the restoration
of photosystem Ⅱ activity[J]. Plant Molecular Biology,
1997,34(1):1-13.
[37] 楊柳青,張柳,廖飛勇,吳紅強,劉志昂,曾紅,朱小青. 低溫脅
迫對南美天胡荽光合作用和葉綠素熒光的影響[J]. 經濟林研
究,2014,32(4):99-102.
YANG Liuqing,ZHANG Liu,LIAO Feiyong,WU Hongqiang,
LIU Zhiang,ZENG Hong,ZHU Xiaoqing. Effects of low temperature
stress on photosynthesis and chlorophyll fluorescence
parameters in Hydrocotyle vulgaris[J]. Nonwood Forest Research,
2014,32(4):99-102.
[38] 王兆,劉曉曦,鄭國華. 低溫脅迫對彩葉草光合作用及葉綠素
熒光的影響[J]. 浙江農業學報,2015,27(1):49-56.
WANG Zhao,LIU Xiaoxi,ZHENG Guohua. Effects of low temperature
stress on the photosynthesis and chlorophyll fluorescence
of Solenostemon scutellarioides[J]. Acta Agriculturae Zhejiangensis,
2015,27(1):49-56.
[39] 劉蕾蕾,紀洪亭,劉兵,馬吉鋒,肖瀏駿,湯亮,曹衛星,朱艷. 拔
節期和孕穗期低溫處理對小麥葉片光合及葉綠素熒光特性的
影響[J]. 中國農業科學,2018,51(23):4434-4448.
LIU Leilei,JI Hongting,LIU Bing,MA Jifeng,XIAO Liujun,
TANG Liang,CAO Weixing,ZHU Yan. Effects of jointing and
booting low temperature treatments on photosynthetic and chlorophyll
fluorescence characteristics in wheat leaf[J]. Scientia Agricultura
Sinica,2018,51(23):4434-4448.
[40] DING Y L,SHI Y T,YANG S H. Advances and challenges in
uncovering cold tolerance regulatory mechanisms in plants[J].
New Phytologist,2019,222(4):1690-1704.
[41] 謝小玉,侯爽,郭金格,汪芬芬,崔凱東,楊奇,陳錦芬,顧開元.
烤煙伸根期對低溫脅迫的生理響應[J]. 中國煙草學報,2022,
28(5):47-55.
XIE Xiaoyu,HOU Shuang,GUO Jinge,WANG Fenfen,CUI
Kaidong,YANG Qi,CHEN Jinfen,GU Kaiyuan. Physiological
responses of flue-cured tobacco to low temperature stress at rootextending
stage[J]. Acta Tabacaria Sinica,2022,28(5):47-55.
[42] BAYSAL F. Effects of low-temperature acclimation on morphological
and physiological indices of banana seedlings[J]. Emirates
Journal of Food and Agriculture,2022,34(5):446-454.
[43] HE W D,GAO J,DOU T X,SHAO X H,BI F C,SHENG O,
DENG G M,LI C Y,HU C H,LIU J H,ZHANG S,YANG Q S,
YI G J. Early cold-induced peroxidases and aquaporins are associated
with high cold tolerance in dajiao (Musa spp.‘Dajiao) [J].
Frontiers in Plant Science,2018,9:282.
