IGF2BP2介導的m6A修飾調(diào)控動物脂肪沉積的研究進展
2023-10-09 06:45:22韓皓哲帖子航龐衛(wèi)軍
畜牧獸醫(yī)學報 2023年9期
韓皓哲,帖子航,龐衛(wèi)軍,蔡 瑞
(西北農(nóng)林科技大學動物科技學院 陜西省動物遺傳育種與繁殖重點實驗室,楊凌 712100)
我國是一個養(yǎng)殖大國,近年來畜禽肉產(chǎn)品總產(chǎn)量保持穩(wěn)中有升,基本能夠滿足消費市場需求。然而隨著人們生活水平的提高,高端優(yōu)質(zhì)肉產(chǎn)品深受消費市場青睞,但目前高端優(yōu)質(zhì)肉的市場供應并不能滿足消費者的需求。脂肪沉積是影響肉品質(zhì)的重要因素,動物體脂分布在軀體皮下、腹腔、肌間和肌內(nèi)等不同部位,其中肌內(nèi)脂肪含量的高低是影響肉質(zhì)的關(guān)鍵因素之一,其與風味、多汁性和嫩度等密切相關(guān),對肉質(zhì)性狀的形成發(fā)揮著重要作用,因此探究脂肪沉積的分子機理對解析動物經(jīng)濟性狀形成的規(guī)律具有重要意義。脂肪沉積受到動物品種、營養(yǎng)水平及表觀遺傳等多方面因素影響。近年來研究發(fā)現(xiàn),DNA甲基化和非編碼RNA調(diào)控等表觀遺傳修飾在動物脂肪沉積中發(fā)揮著關(guān)鍵作用[1-3],表觀遺傳修飾是調(diào)控脂肪沉積的關(guān)鍵因素,而m6A甲基化修飾是真核生物中含量最豐富的一種RNA表觀遺傳修飾,探究其調(diào)控作用與分子機制對脂肪生物學和動物性狀改良具有重要作用[4-5]。因此,本文將綜述有關(guān)m6A修飾、IGF2BP2及其對動物脂肪沉積影響的研究進展,并展望該領(lǐng)域未來的研究方向與發(fā)展趨勢。
1 IGF2BP2介導的m6A修飾概述
1.1 RNA的m6A甲基化修飾過程
RNA是實現(xiàn)遺傳信息在蛋白質(zhì)上表達、促進遺傳信息向表型轉(zhuǎn)化過程的橋梁。絕大多數(shù)生物體的RNA都可以接受多種化學修飾,其中m6A是真核生物mRNA中最豐富的化學修飾之一[6]。m6A是mRNA中腺嘌呤第6位N原子上的甲基化修飾,其修飾位點常存在于mRNA的CDS區(qū)和3′UTR區(qū),主要發(fā)生在RRACH序列上,mRNA上可以含有一個或多個m6A修飾位點[7]。m6A修飾可以影響mRNA的運輸、降解、翻譯以及代謝等,在細胞增殖、分化、凋亡及免疫等多種生物學過程中發(fā)揮重要調(diào)控作用[8-10]。
RNA的m6A修飾涉及3種類型的蛋白質(zhì):甲基轉(zhuǎn)移酶 (writer)、去甲基酶 (eraser)和m6A閱讀蛋白 (reader)。在細胞核內(nèi)存在由多個亞基組成的writer復合體[11-12],此種復合體主要由METTL3、METTL14和Wilms腫瘤相關(guān)蛋白(WTAP)三個必不可少的亞基組成。METTL3充當重要的催化亞基[13-14],而METTL14則提供RNA結(jié)合支架,激活并增強METTL3的催化活性。WTAP與METTL3和METTL14相互作用,從而調(diào)節(jié)RNA的m6A修飾水平。writer復合體的各個亞基相互作用共同執(zhí)行m6A修飾的寫入功能[15]。m6A修飾處于動態(tài)調(diào)控之中,目前已知有兩種不同的酶參與m6A去甲基化過程:FTO和ALKBH5[16-17],FTO是第一個被發(fā)現(xiàn)的m6A mRNA去甲基化酶,FTO被敲除后m6A峰升高[18-19],其調(diào)控脂肪相關(guān)基因的表達,并影響脂質(zhì)代謝多個過程[20-21]。當mRNA進入細胞質(zhì)后會與特定reader蛋白結(jié)合,不同的m6A reader蛋白有著不同的功能,如具有YTH結(jié)構(gòu)域YTHDF1和YTHDF3通過協(xié)同作用提高mRNA的翻譯效率,YTHDC1對于m6A介導的選擇性剪接有一定的作用[22]。m6A reader蛋白除了參與脂肪細胞mRNA的穩(wěn)定、翻譯和降解等過程外(圖1),還參與調(diào)控腫瘤發(fā)生、血細胞生成、病毒復制和免疫應答等生物學過程[23-24]。

圖1 m6A甲基化修飾的作用機制Fig.1 The mechanism of m6A methylation modification
1.2 m6A修飾新閱讀蛋白IGF2BP2
IGF2BP蛋白(包括IGF2BP1/2/3)作為m6A閱讀蛋白的一個新家族,通過識別一致的GG(m6A)C序列靶向數(shù)千個mRNA轉(zhuǎn)錄本[25]。與含有YTH結(jié)構(gòu)域的蛋白家族促進mRNA衰變的功能相反,IGF2BPs蛋白成員在正常和應激條件下以m6A依賴的方式促進靶mRNA的穩(wěn)定性和翻譯,從而影響基因表達豐度[26]。IGF2BP1/2/3可以參與RNA生命周期調(diào)控,每個IGF2BP結(jié)合蛋白都能結(jié)合多種RNA,包括長鏈非編碼RNA (lncRNA)和mRNA,以調(diào)控它們的剪接、運輸、翻譯和穩(wěn)定性[27]。研究表明這三種IGF2BP家族基因在小鼠胚胎中協(xié)同表達,開始于胚胎發(fā)育第10.5天,峰值為第12.5天。IGF2BP1和IGF2BP3的表達在出生后基本消失,而IGF2BP2在出生后廣泛表達,因此研究IGF2BP2對于性狀表觀遺傳機制的調(diào)控尤為重要[28]。IGF2BP2是一種參與調(diào)節(jié)多種生物過程的RNA結(jié)合蛋白,且高度保守,由兩個RNA識別基序結(jié)構(gòu)域和4個hnRNP K同源結(jié)構(gòu)域組成[29],其中K同源結(jié)構(gòu)域是IGF2BP2識別m6A的必要結(jié)構(gòu)域,通過與m6A修飾的相互識別可以提高m6A修飾mRNA的穩(wěn)定性[30]。研究表明IGF2BP2影響糖尿病[31-32]、肥胖癥和脂肪肝等多種疾病[33-34],且關(guān)于癌癥的研究較多[35-37]。在肌肉生物學領(lǐng)域,有研究表明lncMyoD可以直接與IGF2BP2結(jié)合,負調(diào)控IGF2BP2介導的骨骼肌增殖基因N-Ras和c-Myc的翻譯,表明IGF2BP2可以阻止肌細胞增殖以促進成肌分化[38],但對于IGF2BP2是否會依賴m6A或與其它lncRNAs及功能基因mRNA互作來調(diào)控脂肪沉積目前尚不清楚,有必要進行進一步研究。
2 m6A修飾對RNA的影響
2.1 m6A修飾影響RNA前體的剪接過程
真核生物在轉(zhuǎn)錄過程中形成的RNA并不能直接翻譯成蛋白,須經(jīng)過一個剪接過程,剪接過程涉及對mRNA中內(nèi)含子的識別、精準切除以及被切除后的外顯子mRNA連接產(chǎn)生成熟mRNA,此過程中需要剪接因子參與。研究發(fā)現(xiàn),WTAP作為m6A writer復合體成員之一,可以發(fā)揮剪接因子的作用[39],敲除WTAP或METTL3將會產(chǎn)生不同的mRNA亞型。在m6A去甲基化酶FTO敲除的前體脂肪細胞中,m6A修飾水平發(fā)生改變,導致剪接調(diào)控蛋白SRSF2對m6A mRNA進行可變剪接,產(chǎn)生不同的剪接產(chǎn)物,對脂肪沉積產(chǎn)生不同的影響[40]。因此,m6A修飾會影響mRNA的剪接過程。
2.2 m6A修飾調(diào)控RNA的出核
RNA經(jīng)剪接后進入到細胞質(zhì),出核的過程受到m6A修飾的調(diào)控。研究表明細胞中存在一種TAP-P15復合體[41],而ASF/SF2的磷酸化水平?jīng)Q定這種復合體與mRNA的結(jié)合能力,當m6A去甲基化酶ALKBH5缺失時,m6A修飾水平增強使ASF/SF2去磷酸化程度上升,導致TAP-P15復合體與mRNA的結(jié)合能力上升,mRNA的核輸出能力增強[42]。DEAD-box(DDX)解旋酶對RNA的識別和新陳代謝至關(guān)重要。DDX46通過其特有的螺旋結(jié)構(gòu)域招募ALKBH5使與之結(jié)合的信號分子RNA去甲基化,導致其RNA不能出核[43]。因此,m6A修飾可以調(diào)控mRNA的出核過程。
2.3 m6A修飾調(diào)控RNA的穩(wěn)定性和降解
遺傳信息通過表達蛋白實現(xiàn)其生物學功能,mRNA是用于翻譯蛋白質(zhì)的直接模板。大多數(shù)m6A修飾富集于外顯子區(qū)和終止密碼子區(qū)域[44-45]。在前體mRNA被剪接后形成成熟mRNA的過程中,m6A修飾被保留下來存在于成熟mRNA中。在調(diào)控mRNA穩(wěn)定性方面,IGF2BP采用一種m6A依賴性方式來提高目標mRNA的穩(wěn)定性[23]。HUR作為另外一種RNA結(jié)合蛋白,在各種組織中普遍表達[46],其包含核質(zhì)穿梭序列(HNS),HNS的轉(zhuǎn)錄后修飾控制HUR在細胞核和細胞質(zhì)之間的移位。HUR的功能依賴于動態(tài)的亞細胞定位,一旦HUR從細胞核穿梭到細胞質(zhì),導致HUR增加了目標RNA的穩(wěn)定性和翻譯[47-49]。當m6A與含有YTH結(jié)構(gòu)域的YTHDF2結(jié)合后會導致m6A修飾的mRNA降解[50-51]。此外,有研究發(fā)現(xiàn)敲低胚胎干細胞中METTL3或METTL14后,其靶mRNA的穩(wěn)定性降低[52-53]。因此,m6A修飾會影響m6A mRNA的穩(wěn)定性與降解。
3 IGF2BP2介導的m6A修飾調(diào)控動物脂肪沉積
3.1 調(diào)控白色脂肪沉積
白色脂肪是脂肪沉積的主要場所,其廣泛分布在動物機體皮下組織和內(nèi)臟周圍,其中皮下脂肪含有較高的飽和脂肪酸,其攝入量過高會導致膽固醇、甘油三酯以及低密度脂蛋白升高,增加患各種心血管疾病的風險[54];而肌內(nèi)脂肪含量與肉品質(zhì)以及胰島素敏感性等緊密相關(guān)[55]。因此,研究白色脂肪沉積不僅對于動物肉質(zhì)的改善有重要作用,且對人類健康也具有一定意義。在白色脂肪的成脂過程中,m6A表觀遺傳修飾發(fā)揮著不可或缺的作用。lncRAP2的表達具有很高的組織特異性,其能誘導白色脂肪細胞的分化,lncRAP2主要存在于細胞質(zhì)中,不與核糖體和RNA直接結(jié)合,但可以與RNA結(jié)合蛋白IGF2BP2結(jié)合形成復合體,影響RNA穩(wěn)定性或降解(圖2)。通過分析白色脂肪細胞中IGF2BP2靶向的mRNAs,發(fā)現(xiàn)其選擇性地與編碼成脂調(diào)節(jié)因子和能量消耗相關(guān)的mRNAs結(jié)合,如瘦素、脂聯(lián)素及白介素等[56-58],表明這些成脂調(diào)控因子轉(zhuǎn)錄本上可能存在m6A修飾位點。此外,研究發(fā)現(xiàn)lncRNAHilnc與IGF2BP2相互作用調(diào)節(jié)PPAR途徑介導的肝脂質(zhì)沉積[59](圖2)。全身敲除IGF2BP2抵抗高脂誘導的肥胖和脂肪肝發(fā)生,而在肝組織特異性過表達IGF2BP2會導致肝的脂肪變性[60]。肝細胞特異性IGF2BP2敲除小鼠導致PPARα、PPARγ和CPT1A等基因的表達水平的顯著下調(diào),使體內(nèi)的甘油三酯的運轉(zhuǎn)速率加快,抑制肝脂肪沉積。因此,IGF2BP2通過m6A依賴的方式識別成脂或脂代謝因子以調(diào)控靶mRNA的穩(wěn)定性影響脂肪沉積[61-63]。
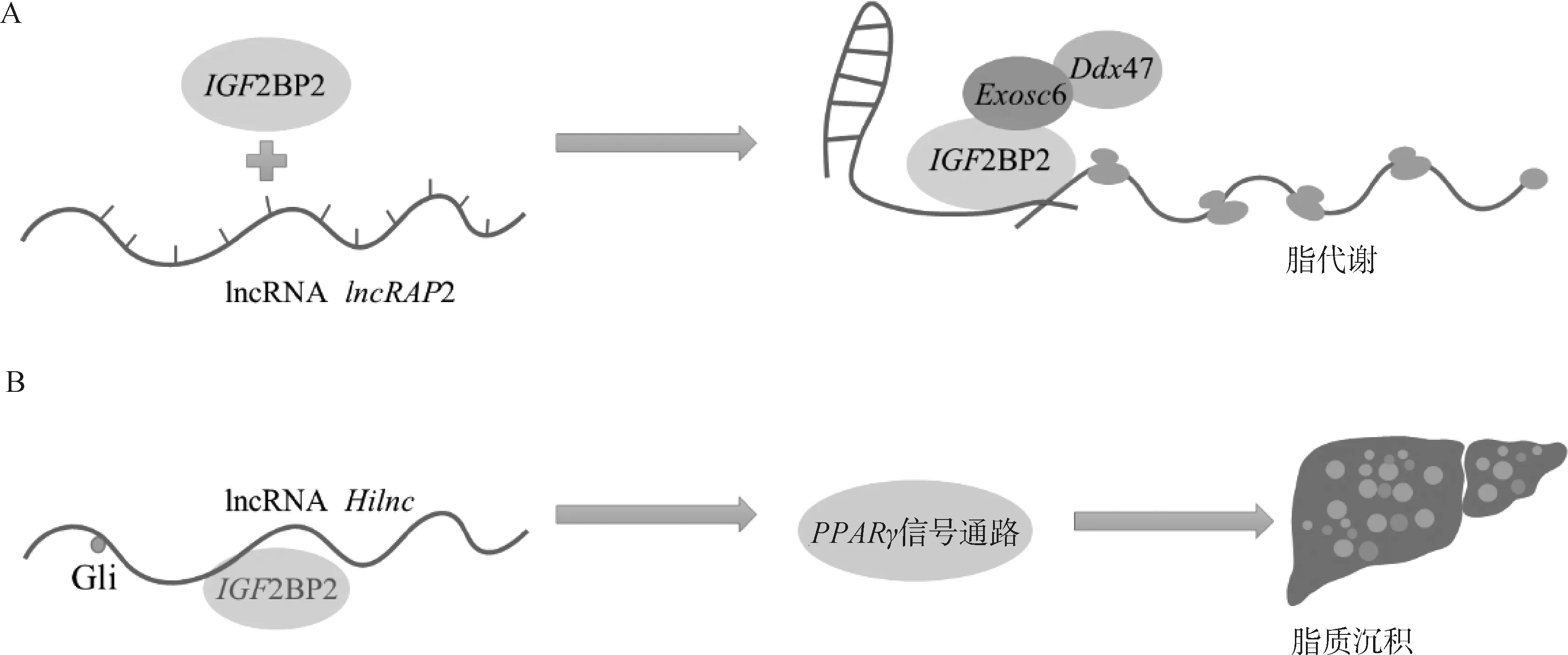
A. IGF2BP2與lncRNA lncRAP2結(jié)合通過招募Exosc6和Ddx47調(diào)控脂代謝基因表達;B. IGF2BP2與lncRNA Hilnc互作通過PPARγ調(diào)控肝脂質(zhì)積累A. IGF2BP2 regulates lipid metabolism by binding to lncRNA lncRAP2 and recruiting Exosc6 and Ddx47; B. The interaction between IGF2BP2 and lncRNA Hilnc regulates hepatic lipid accumulation through PPARγ圖2 IGF2BP2與lncRNA互作調(diào)控脂質(zhì)代謝的作用機制Fig.2 The mechanism of interaction between IGF2BP2 and lncRNA regulating lipid metabolism
在豬脂肪沉積方面,發(fā)現(xiàn)IGF2BP2等RNA結(jié)合蛋白與選擇性剪接事件區(qū)域內(nèi)的核心剪接體相互作用,進行前體mRNA剪接,并對脂質(zhì)代謝造成影響[64-65]。此外,研究表明IGF2BP2蛋白與IGF2具有較強的親和力[66],可調(diào)控IGF2的生理作用,而IGF2基因可作為豬脂肪沉積的關(guān)鍵基因[67]。以上研究表明IGF2BP2可能對豬脂肪沉積發(fā)揮重要調(diào)控作用。在羊脂肪沉積方面的研究發(fā)現(xiàn)IGF2BP2在未成年山羊的肌肉和脾組織表達量最高,腎、卵巢、胃和大腦次之,肺、心和肝表達量相對較低;而在成年山羊中,IGF2BP2表達量最高的組織是脂肪,輸卵管、卵巢、心和肺次之,腎、肌肉、脾和肝表達量相對較低,推測IGF2BP2在山羊個體不同發(fā)育時期發(fā)揮著重要的作用,特別是在脂肪沉積過程中[68]。
3.2 調(diào)控棕色脂肪產(chǎn)熱
棕色脂肪組織是動物體內(nèi)呈棕色樣的脂肪組織,其脂肪細胞體積較小,胞質(zhì)中有多個較小的脂滴,并有較多的線粒體[69]。棕色脂肪的功能是促進適應性產(chǎn)熱、ATP產(chǎn)生和底物氧化的解偶聯(lián),通過非顫抖產(chǎn)熱的方式來維持體溫變化。因此,棕色脂肪細胞能夠通過氧化其自身儲存的脂質(zhì),從而產(chǎn)生熱量并增加代謝率[70]。UCP1是一個很重要的解偶聯(lián)基因,激活該基因可顯著抑制飲食誘導的肥胖。研究發(fā)現(xiàn)IGF2BP2對UCP1基因具有轉(zhuǎn)錄抑制作用,在小鼠上敲除IGF2BP2導致棕色脂肪的UCP1蛋白豐度增加,造成棕色脂肪產(chǎn)熱能力增強,并對飲食誘導的肥胖和脂肪肝具有更強的抵抗力,以及更高的葡萄糖耐受性、胰島素敏感性和更好的御寒能力[71]。因此,IGF2BP2介導的m6A修飾與棕色脂肪產(chǎn)熱息息相關(guān)。
4 總結(jié)與展望
綜上所述,IGF2BP2主要通過m6A依賴的方式影響脂肪沉積靶基因的穩(wěn)定性,影響其RNA的豐度,進而調(diào)控動物脂肪沉積,但IGF2BP2介導的m6A修飾調(diào)控動物脂肪沉積的分子機制仍需進一步深入探究。因此,后續(xù)研究可借助多組學測序技術(shù)以及新的分子生物學方法挖掘IGF2BP2結(jié)合的m6A修飾下游靶基因,明確IGF2BP2介導的m6A修飾調(diào)控動物脂肪沉積的表觀遺傳機制,為畜禽遺傳改良提供關(guān)鍵基因,并加快畜禽性狀遺傳改良進程,從而改善畜禽肉類品質(zhì),增加經(jīng)濟效益,滿足國民對優(yōu)質(zhì)肉產(chǎn)品的需求。
