全基因組選擇信號解析劍白香豬和從江香豬的遺傳差異
2023-10-09 06:45:26張雨丹張依裕顧曉龍楊紅文
畜牧獸醫學報 2023年9期
袁 巍,畢 歡,張雨丹,張依裕,顧曉龍,楊紅文,陳 偉*
(1.貴州大學動物科學學院 高原山地動物遺傳育種與繁殖教育部重點實驗室,貴陽 550025;2.貴州省動物遺傳育種與繁殖重點實驗室,貴陽 550025;3.貴州大學香豬研究所,貴陽 550025;4.貴州省貴陽德康農牧有限公司,貴陽 550025;5.貴州省畜禽遺傳資源管理站,貴陽 550025)
豬的養殖在我國歷史久遠、分布廣泛、數量眾多。我國復雜的地貌環境、多元化的飲食文化帶來的多樣化選育目標是中國形成多個地方豬種質資源的重要原因,多樣化選育目標致使我國是世界上地方豬種質資源最豐富的國家,約占世界豬種質資源的1/3[1]。許多本地豬品種都有自己獨特的特點,例如抗病性、飼料效率和高生育率。貴州香豬是一種小型豬種,包括從江香豬和劍白香豬兩種類型。在貴州獨特的喀斯特地貌和多民族混居的人文環境的影響下,形成了近交不退化的優勢和基因高度純合的特點。作為貴州地區寶貴的地方豬種遺傳資源,通過全基因組選擇信號解析劍白香豬和從江香豬的遺傳差異,有助于提高對從江香豬和劍白香豬種群的保護狀況、遺傳多樣性、群體結構和和適應性進化的認識,制定更有效的保護策略以維護地方豬的遺傳多樣性和生態系統的穩定性。由于長期以來的豬種雜交和人工選擇,地方豬種的遺傳多樣性正在不斷減少[2],本地豬種和群體大小的數量均大幅減少[3],因此,評估中國本地豬品種的遺傳多樣性和群體結構成為必要的舉措,以便制定更加有效的保護策略,避免品種的基因流失。隨著測序技術和計算方法的快速發展以及測序費用的大幅下降,二代測序技術已被廣泛應用于家畜遺傳多樣性和群體結構的研究中,如豬[4-5]、牛[6]、羊[7-8]、雞[9-10]、鴨[11]等。通過對基因組水平的研究,這些研究篩選出了大量與適應性、表型、重要經濟性狀等相關的候選基因和遺傳標記。這些研究結果不僅有助于人們深刻了解家畜物種的起源、馴化機制、人工選擇和自然選擇,而且還為家畜的遺傳改良和育種提供了基礎。目前,對于從江香豬和劍白香豬的研究主要集中在基因的表達[12]、經濟性狀[13]、生產性能[14]上,在基因組層面上,對劍白香豬和從江香豬的遺傳差異研究較少。因此,了解從江香豬和劍白香豬種群的保護狀況、遺傳多樣性、群體結構和適應性進化等方面的信息非常重要。通過全基因組選擇信號分析,揭示了劍白香豬和從江香豬的遺傳差異,為實現上述目標提供重要的基礎和支持。
1 材料與方法
1.1 樣品采集
隨機選取貴州保種場2歲齡從江香豬和劍白香豬各5頭,采集其耳組織,用DEPC水處理后放入無RNA酶的凍存管中,液氮中冷凍,后轉入-80 ℃的超低溫冰箱儲存,用于提取DNA。從江香豬及劍白香豬如圖1所示。

圖1 從江香豬和劍白香豬照片Fig.1 Photos of Congjiang and Jianbai Xiang pigs
1.2 建庫測序和數據處理
委托北京諾禾致源生物信息科技有限公司完成從江香豬與劍白香豬的總DNA提取和建庫測序,主要測序平臺為Illumina HiSeq 2500,測序策略為PE125,深度為5 x~10 x。用Fsatp v0.19.10軟件[15]對測序原始數據進行過濾,去掉adaptor和低質量序列,將過濾后數據用bwa v0.7.17-r1188軟件比對到豬參考基因組Sscrofa11.1上,并進行排序和去重復處理,獲得bam文件用于變異檢測。
1.3 變異檢測和過濾
1.4 遺傳多樣性分析
群體有效大小(Ne)、多態位點比例(PN)、觀測雜合度(Ho)和期望雜合度(He)是分析種群遺傳多樣性的常用參數。利用SNeP v1.11 軟件[16]計算群體有效大小Ne,使用Plink v1.90b6.21軟件[17]計算多態位點比例(PN)、觀測雜合度(Ho)和期望雜合度(He)。有效群體大小Ne根據不同的SNP數據量和歷史背景進行計算,因此其接近最近世代的程度也會不同。
1.5 群體選擇信號分析
群體間遺傳分化指數(the fixation index,Fst)、核苷酸多樣性 (nucleotide diversity,π)與群體內選擇檢驗(Tajima’s D)均由vcftools v0.1.16軟件基于數據集Data1計算,窗口大小為100 000 bp,滑動距離為1 000 bp,分組信息為從江香豬(coj)和劍白香豬(jab),最后用R包ggplot2可視化。具體參數為--fst-window-size 100 000 --fst-window-step 10 000,--window-pi 100 000 --window-pi-step 10 000,--TajimaD 100000。群體間分歧度(the reduction of diversity,ROD)值公式為1-π1/π2,其中π1為劍白香豬群體平均π值,π2為從江香豬群體平均π值。
1.6 受選擇區域分析
分別取Fst和ROD結果前5%位點,提取其在染色體上位置等信息,用R編寫腳本合并為Fst和ROD是否同時滿足前5%位點信息表。將Fst閾值線設置為0.2,ROD閾值線設置為0.55,利用R包ggplot2繪制點圖。并將同時滿足Fst和ROD前5%的位點使用bedtools v2.30.0合并重疊窗口位點,然后根據合并位點提取候選基因信息。具體參數為“bedtools intersect -F 0.1 -b gff3,bedtools merge” 。
1.7 注釋與通路分析
將已鑒定到的豬特有家族基因和顯著收縮與擴張基因與KEGG、GO數據庫比對,進行通路分析。KEGG通過R包clusterProfiler v4.0[18]調用KEGG數據庫官網(https://www.kegg.jp/)API直接用于KEGG分析,并通過R包內置函數dotplot進行可視化分析;GO通過R包org.Ss.eg.db v3.16.0(http://bioconductor.org/packages/release/BiocViews.html#___OrgDb)進行分析,并使用clusterProfiler內置函數dotplot進行可視化分析;最后通過在線網站基迪奧生信云工具(https://www.omicshare.com/tools/Home/Soft/cog)進行可視化。
2 結 果
2.1 SNP及INDEL檢測結果
從江香豬和劍白香豬中平均檢測到2 100萬個SNPs位點,其中從江香豬23 021 994個,劍白香豬20 433 662個。同樣平均檢測到460萬個INDELs位點,其中從江香豬4 949 456個,劍白香豬4 389 623個。在外顯子區域,均檢測到的SNPs位點數約為17萬個,其中從江香豬188 674個,劍白香豬164 383個;平均檢測到的INDELs位點數約為10萬個,其中從江香豬11 841個,劍白香豬10 613個。這些外顯子區域上的SNP和INDEL變異可能會影響蛋白質的結構、功能、表達和穩定性,從而影響生物體的表型。利用R包Venn Diagram根據SNPs位置信息繪制的韋恩圖顯示,從江香豬和劍白香豬群體共有的SNPs位點數為16 158 002個,從江香豬特有的為6 863 992個,劍白香豬特有的為4 275 660個(圖2A);共有的INDEls位點數為3 275 375個,從江香豬特有的為1 674 081個,劍白香豬特有的為1 114 248個(圖2B)。
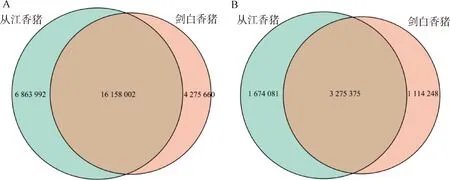
圖2 從江香豬和劍白香豬SNP(A)及INDEL(B)檢測結果韋恩圖Fig.2 The Venn diagrams depicting the SNP (A) and INDEL (B) detection results of Congjiang and Jianbai Xiang pigs
2.2 從江香豬和劍白香豬群體的遺傳多樣性分析
利用plink軟件分析群體有效大小(Ne)、多態位點比例(PN)、觀測雜合度(Ho)和期望雜合度(He),結果如表1所示。通過plink計算的劍白香豬和從江香豬的最近世代群體有效大小為18頭,最近世代為13。從江香豬的多態性位點比例為0.875 7,高于劍白香豬的多態性位點比例0.813 2。另外,從江香豬和劍白香豬各自群體內的觀測雜合度均大于期望雜合度,說明二者群體內部都存在較高的遺傳多樣性。選擇壓力等因素導致群體內的雜合子數量增加,有利于個體對環境的適應和進化。
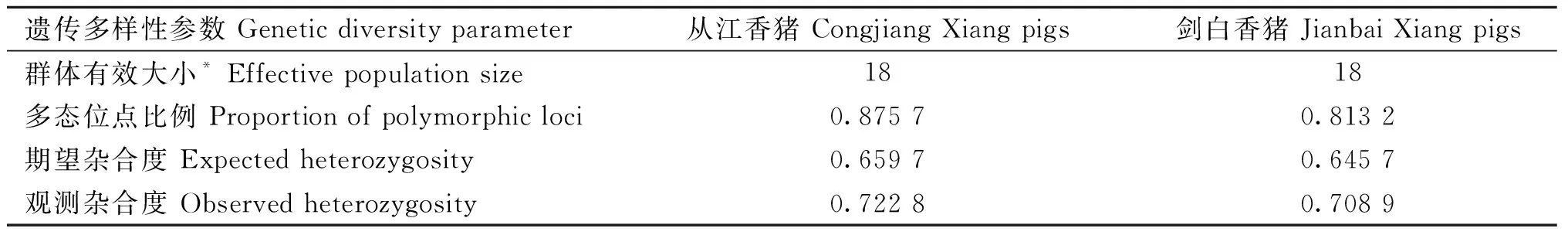
表1 從江香豬和劍白香豬遺傳多樣性參數Table 1 The genetic diversity parameters of Congjiang and Jianbai Xiang pigs
2.3 劍白香豬和從江香豬群體選擇分析
以從江香豬作為參照組,對劍白香豬進行群體壓力分析。通過滑動窗口計算從江香豬(coj)和劍白香豬(jab)群體的核苷酸多樣性π值(圖3A,B)、群體遺傳分化指數Fst(圖3C)。統計從江香豬和劍白香豬每條染色體上高于0.01閾值的核苷酸位點數量(圖3D),結果顯示從江香豬的每條染色體上均存在高核苷酸多態性位點,而劍白香豬的chr4上不存在高核苷酸多態性位點。從江香豬的總核苷酸多態性位點數為1 288個,高于劍白香豬的1 035個。Fst圖中,每條染色體均存在大量高于閾值線的多態性位點,表明劍白香豬和從江香豬之間在每個染色體上都存在遺傳分化情況。
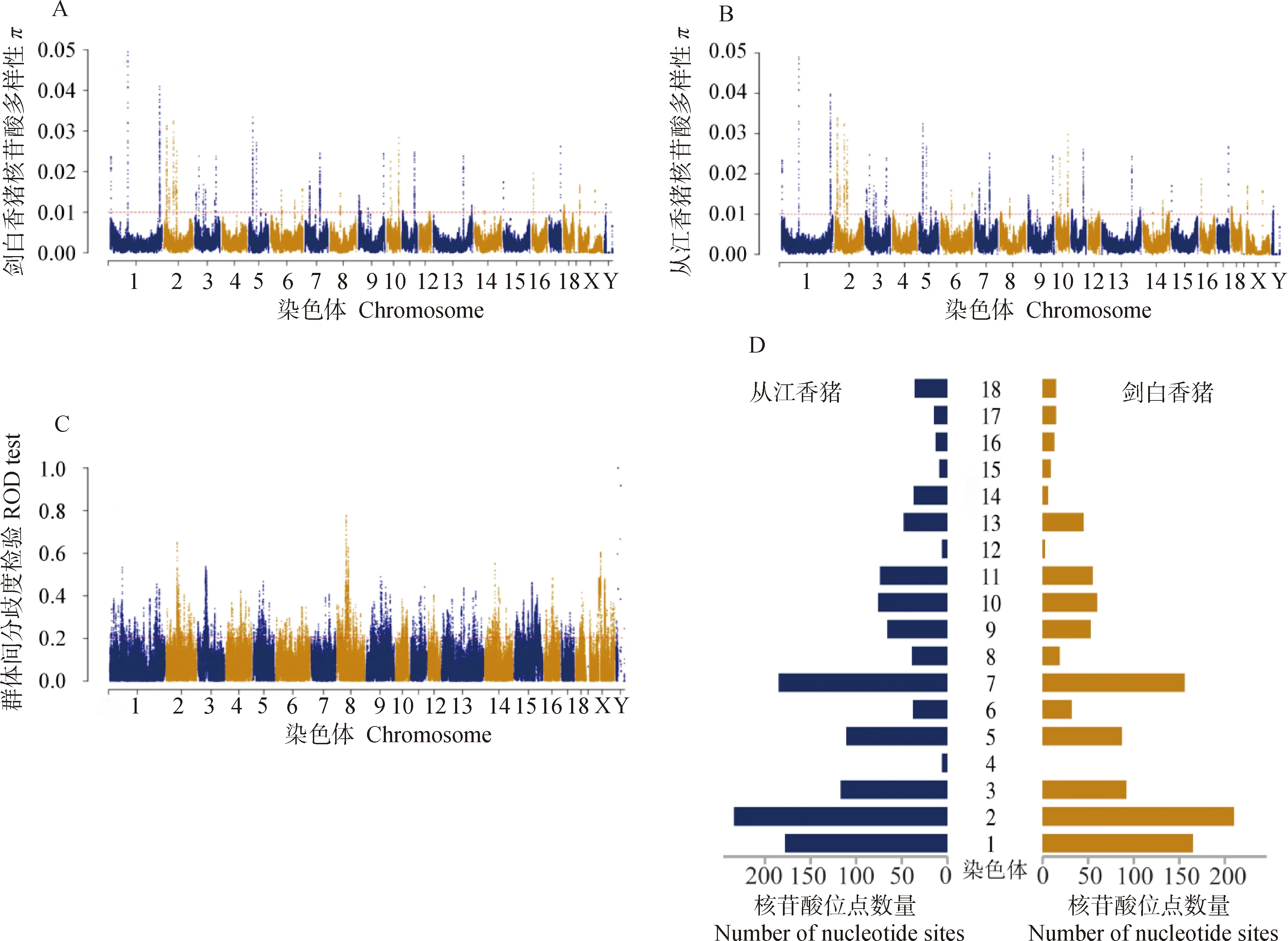
圖3 從江香豬和劍白香豬的π值、Fst值以及核苷酸多態性在染色體上的分布Fig.3 The distribution of π values, Fst values, and nucleotide polymorphisms along the chromosomes of the Congjiang and Jianbai Xiang pig breeds
2.4 從江香豬和劍白香豬分化水平
將從江香豬和劍白香豬基因組中每條染色體上的核苷酸多樣性π值、群體間分歧度檢驗Fst、群體分歧度ROD以及群體內選擇檢驗Tajima’sD的計算結果取均值后繪制成圖4。結果表明,從江香豬和劍白香豬的核苷酸多樣性均小于0.5%,且群體內選擇檢驗Tajima’sD值均大于0。此外,劍白香豬和從江香豬的ROD值為0.553 1,Fst值為0.736 2。
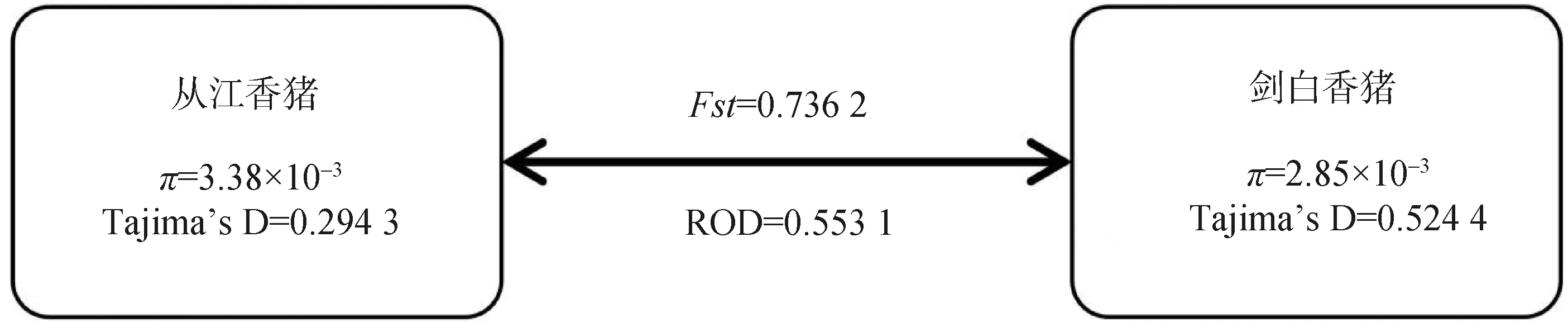
圖4 從江香豬和劍白香豬分化水平參數Fig.4 The differentiation level parameters of Congjiang and Jianbai Xiang pigs
2.5 劍白香豬群體受選擇區域
對于受選擇區域的篩選,一般考慮群體間分歧度高及群體分化率高的區域。以從江香豬為對照組,結合群體間分歧度ROD和群體間分歧度檢驗Fst的計算結果,篩選出劍白香豬基因組中前5%受到強烈選擇作用、高度分化的區域,并從中識別出了共計775個受選擇區域基因,其分布情況如圖5紅色區域所示。
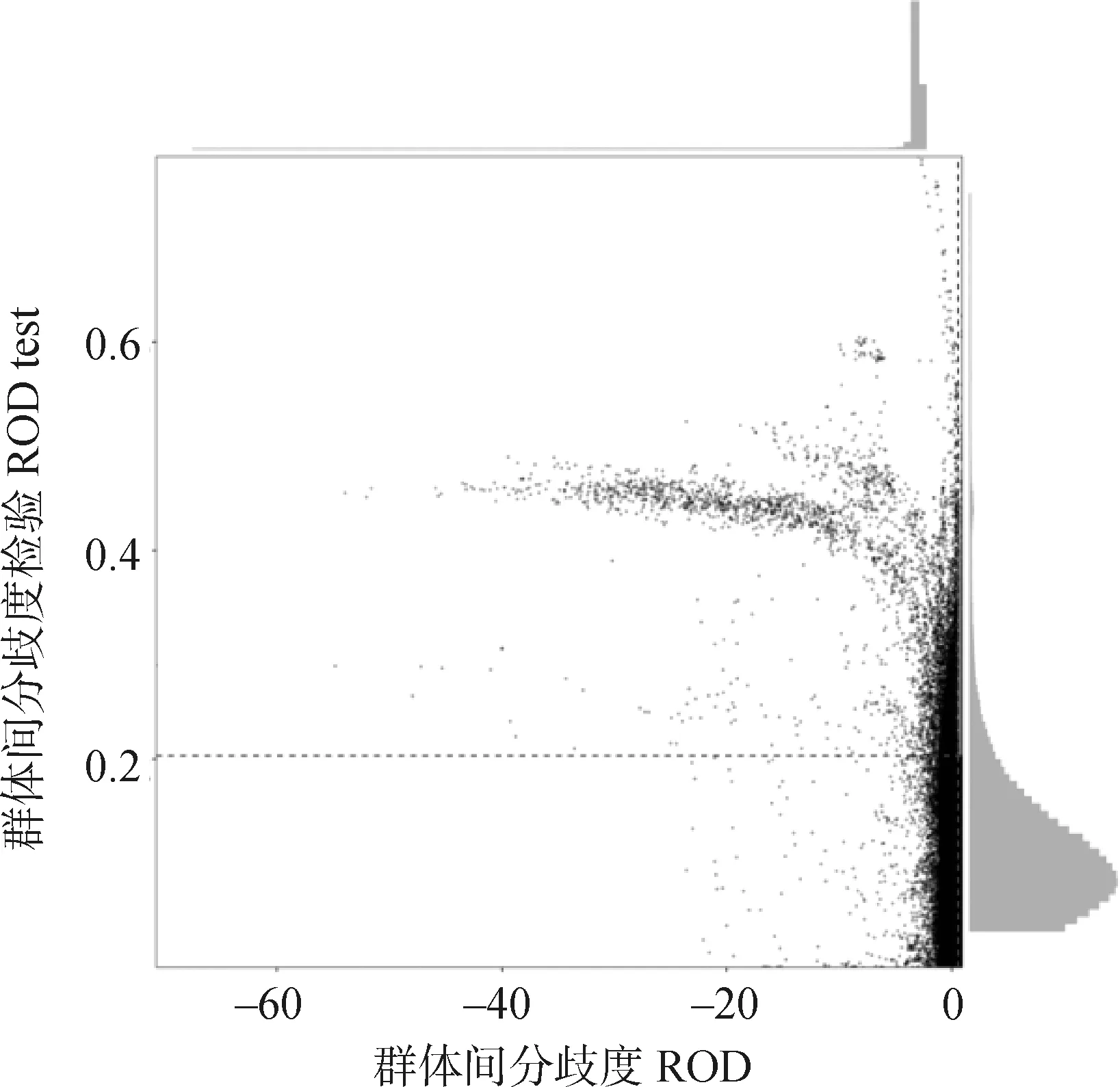
圖5 劍白香豬Fst-ROD的受選擇區域Fig.5 The Fst-ROD regions under selection in Jianbai Xiang pigs
2.6 劍白香豬群體受選擇區域基因富集分析
對受選擇區域基因進行了KEGG富集分析(圖6A),結果表明前20種KEGG富集通路涉及軸突引導、ECM與受體的相互作用、局部粘附、PI3K-Akt信號通路、硫辛酸代謝、cGMP-PKG信號通路、MAPK信號傳導途徑、核苷酸切除修復、阿佩林信號通路、心律失常性右心室心肌病等通路。
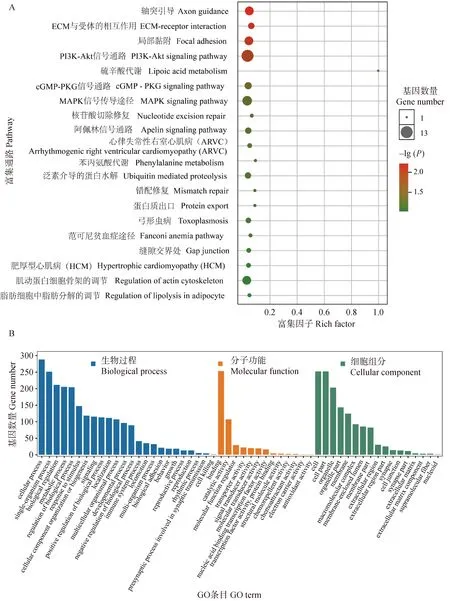
圖6 劍白香豬群體受選擇區域基因KEGG(A)和GO分析(B)結果Fig.6 The KEGG (A) and GO analysis (B) results of the genes located in the selected regions in the population of Jianbai Xiang pigs
同時,對受選擇區域基因進行了GO富集分析(圖6B),發現在從江香豬和劍白香豬的775個候選基因中,有367個基因顯著富集于751個GO條目(P<0.05),其中包括113個分子功能類,553個生物過程類,85個細胞組分類。在分子功能中,受選擇區域基因主要富集在結合和催化活性;在生物過程中,受選擇區域基因富集最集中的GO條目在細胞過程、單一生物體過程、生物調節、新陳代謝過程和生物過程的調節上;在細胞組分中,細胞、細胞部分、細胞器條目上的基因富集較多。
3 討 論
遺傳分化由長期自然選擇和人工選擇造成,受環境、基因漂變、基因流動和遺傳漂變等因素的影響。ROD基于兩個群體間的核酸多態性π的差異識別信號,被引入用于測量兩群體間的群體多態性損失,ROD平均值越接近1,群體多態性損失越大。本研究中,從江香豬和劍白香豬的ROD值為0.553 1,表示從江香豬和劍白香豬之間存在一定的群體多態性損失,群體分歧度較高。Fst值主要受遺傳漂變和遷移等因素的影響,種群中對于特定環境經歷的適應性進化會增大種群分化水平,當Fst值大于0.25時表示群體間遺傳分化程度較大,Fst越接近1,表明兩種群間越存在明顯的種群分化[19]。尤桂爽等[20]對四川盆地6個地方雞群體的遺傳關系研究發現,豐巖烏骨雞和巖水雞黑羽群體之間具有較高的遺傳分化(Fst=0.183)。本研究中,從江香豬和劍白香豬的Fst值為0.736 2,結果顯示從江香豬種群和劍白香豬種群之間存在著明顯的種群分化。群體內選擇檢驗Tajima’s D用于檢驗正選擇效應[21],本研究中從江香豬和劍白香豬的Tajima’s D值均大于0,說明二者基因組內存在大量的中等頻率等位基因,可能是由于群體瓶頸效應,群體結構或者平衡選擇等原因所致。
群體有效大小越小意味著該群體的基因多樣性越低,本研究發現,從江香豬和劍白香豬在13個世代之前的群體有效大小為18頭,明顯低于商品豬群體有效大小[22],同時也低于目前報道的梅山豬(50~65頭)[23]、陸川豬(35頭)[24]和撒壩豬(21頭)[25],高于涼山豬(15頭)[26]等中國其他地方豬種群體有效大小。多態位點比例與群體中等位基因的數目和頻率有關,用來表示群體中某一位點多態性的程度,以評估群體的遺傳多樣性[27],本研究發現從江香豬和劍白香豬群體的多態位點比例分別為0.875 7和0.813 2,稍低于西方豬種0.90~0.98的多態位點比例范圍[28],劍白香豬的多態位點比例稍低于里岔黑豬(0.827)[29]。說明從江香豬和劍白香豬相比其他群體遺傳多樣性較低,這對一個種群的長期生存和變化環境的適應是不利的。期望雜合度是指理論計算得出的雜合度,觀測雜合度是指隨機抽取的兩個樣本的等位基因不相同的概率。若觀測雜合度高于期望雜合度,表明群體內存在較高的遺傳多樣性,群體可能受到遷徙或者基因流的影響[30]。本研究中,從江香豬和劍白香豬的期望雜合度均高于觀測雜合度,說明二者群體可能出現過遷徙事件或者有外來血緣的引入,這與杭豬[31]、青峪豬[32]、馬身豬[27]、通城豬[33]、丫杈豬[34]群體的研究結果相似。
雖然劍白香豬與從江香豬同為小型豬種,但本研究中的5個劍白香豬樣品中有3個經過14代自交培育,相比之下,劍白香豬種群更為純正,遺傳背景更為穩定。根據KEGG分析結果,劍白香豬和從江香豬的受選擇區域基因中差異表達的通路涉及了多個生物學過程和信號通路,其中包括涉及軸突引導、ECM與受體的相互作用、局部粘附、PI3K-Akt信號通路、硫辛酸代謝、cGMP-PKG信號通路、MAPK信號傳導途徑、核苷酸切除修復、阿佩林信號通路、心律失常性右室心肌病等通路。從這些通路中可以初步推斷出劍白香豬在某些方面的優勢。PI3K-Akt 信號通路和MAPK 信號通路等與生長和發育[35]、代謝調節[36]、細胞存活和凋亡[37]等生物學過程有關,因此劍白香豬在這些方面可能表現出更好的生物學特性。硫辛酸代謝也與肝臟解毒、生物體對藥物的代謝、膽固醇代謝、維生素代謝相關[38],因此劍白香豬在物質代謝方面也可能有優勢。豬由于其在體型、解剖結構、生理、營養物質代謝、藥物代謝和疾病發展方面與人類相似,所以被廣泛作為人類疾病研究的生物模型[39],Fanjul等[40]就使用小鼠和豬模型來研究衰老對心臟代謝疾病的影響,本研究KEGG分析結果中,劍白香豬群體受選擇區域基因富集到心律失常性右室心肌病通路,可以考慮將劍白香豬作為研究心律失常性右心室心肌病的生物醫學模型。根據GO富集分析結果,受選擇區域基因富集在結合和催化活性條目,這說明劍白香豬在分子功能方面具有更多的活性相關基因,這可能意味著該品種在代謝、生長和發育等方面具有更強的能力;而在細胞過程、單一生物體過程、生物調節、新陳代謝過程和生物過程調節的富集,表明劍白香豬在生物過程和生物調節方面可能具有更為豐富的功能;細胞、細胞部分、細胞器條目的富集,表明劍白香豬在細胞結構和細胞器的功能方面具有更多的相關基因。
4 結 論
綜上所述,4個統計數據(Ne、PN、Ho、He)共同表明,需要提高從江香豬和劍白香豬群體內的遺傳多樣性,減少外來血緣引入導致純種度降低的風險。群體全基因組選擇信號分析表明,劍白香豬和從江香豬之間存在一定的群體多態性損失,群體分歧度較高,兩種群間存在明顯的種群分化,而二者群體受選擇區域基因富集分析表明,劍白香豬在代謝、生長和發育等方面具有更強的能力。自交培育的劍白香豬可以用作研究心律失常性右室心肌病的動物模型。這些發現有助于更深入地了解貴州地方香豬品種的遺傳特征和潛在用途,科學制定對貴州地方香豬的保護和利用策略,以保障其種質資源的可持續利用。
