鎘脅迫下地膚根際與非根際土壤微生物群落結構及多樣性
2023-10-10 06:49:14肖銳譚璐吳亮張皓郭佳源楊海君
中國農業科技導報 2023年8期
關鍵詞:植物
肖銳, 譚璐, 吳亮, 張皓, 郭佳源, 楊海君*
(1.湖南農業大學資源環境學院,長沙 4 101281; 2.長沙經開區管委會,長沙 410100;3.核工業二三〇研究所,長沙 410007)
土壤微生物是土壤生態系統的重要組成部分,對重金屬污染土壤的植物生長以及重金屬的生物地球化學循環起到重要作用[1-2]。在所有土壤微生物中,根際微生物被定義為“受根周圍土壤區域影響的微生物”,其與植物根系的直接接觸而影響著植物根際活動[3]。實際上,每種植物在其根際都有特定的微生物群落,這在促進宿主植物生長和重金屬解毒等方面均起著重要作用[4]。在重金屬污染的土壤中,土壤微生物通過不斷調整自身群落結構以及代謝能力以適應不同的生長環境[5],例如:土壤微生物可通過增加碳氮固定化微生物的豐度以及代謝酶的活性來增強植物對重金屬毒性的抵抗能力[6],同時,一些根際細菌可以通過細胞內積累[7]、細胞外沉淀[8]、重金屬離子的吸附或轉化[9]來降低重金屬對植物的毒性并防止其遷移。因此,土壤根際微生物群落對植物健康、養分循環和植物修復過程至關重要[10]。隨著高通量測序技術在微生物多樣性與功能研究方面的應用[11-13],人們利用高通量測序揭示了土壤中微生物豐度與環境參數相關,鑒定出了對重金屬耐受和敏感的微生物[14]。在植物修復土壤重金屬污染方面,研究者發現隨著重金屬毒性的增強,芒草的修復能力逐漸減弱,且重金屬毒性改變了芒草土壤微生物群落的結構[15]。
地膚(Kochia scopariaL.Schrad.)為藜科地膚屬一年生草本植物,極耐旱、耐鹽堿、耐貧瘠、適生性強、根系發達、生長快、生物量大。目前對地膚的研究集中于其營養成分、耐鹽生理特性[16-17]、栽培與園林應用等方面,也有研究報道了地膚對礦區土壤重金屬的累積特性[18-20]、耐受性和富集能力[21-22],尚未見鎘(Cd)脅迫對地膚根際與非根際土壤微生物群落結構及多樣性影響的報道。本研究以初始土壤pH 6.1、總Cd 2.753 mg·kg-1脅迫下培植150 d的地膚盆栽土壤為供試樣品,采用高通量測序技術研究Cd 脅迫下地膚根際和非根際土壤中微生物群落結構特征,明晰地膚對土壤Cd污染的耐受性與土壤微生物群落構成變化的聯系,以期為地膚修復Cd污染土壤提供理論依據。
1 材料與方法
1.1 試驗設計及樣品采集
2021 年10 月14 日在湖南農業大學耘園基地選取初始pH 6.1、土壤總Cd 2.753 mg·kg-1脅迫下培植150 d 長勢一致的盆栽地膚根際和非根際土壤為研究對象(該處理下地膚對土壤Cd的富集系數和轉運系數均最大[23],分別達4.22、1.71),采集地膚根際和非根際土壤樣品。根際土壤樣品采用抖落法[24]采集,挖取盆中整株地膚,揉捏地膚莖稈,拍打掉大塊土壤,輕抖根部,抖落土壤后,用滅菌的刷子輕刷黏附在地膚根表面的土壤,即為每株地膚的根際土壤(G),將3 盆6 株的根際土壤充分混勻則為1 個根際土壤樣品。輕輕抖落大塊不含根系的土壤后,用陶瓷剪刀取下附著在根系周圍的土壤,作為非根際土壤(F),3 盆的非根際土壤充分混勻為1 個樣本的非根際土壤樣品。將根際和非根際土壤樣品各分成2 份,用無菌自封袋封裝,一份用于土壤理化性質、養分及總Cd測定;另一份置于冷卻泡沫箱中立即帶回實驗室,于超凈工作臺上用2 mL 離心管進行分裝,并立即轉移到-80 ℃超低溫冰箱保存,樣品冰凍寄存送生工生物工程(上海)股份有限公司提取土壤微生物。每個處理設置3個重復,分別命名為G-1、G-2、G-3和F-1、F-2、F-3。
1.2 土壤理化性質、養分及Cd含量
土壤pH 值采用電位法[24]測定;土壤含水量采用烘干法[25]測定;土壤有機質含量采用直接電位滴定法[26]測定;土壤全氮(total nitrogen,TN)含量采用凱氏定氮法[27]測定;堿解氮(alkaline hydrolysis nitrogen,AN)含量利用堿解擴散法[27]測定;全磷(total phosphorus,TP)和速效磷(available phosphorus,AP)含量采用鉬銻抗比色法[27]測定;土壤總Cd含量采用王水—高氯酸消解法[28]測定。
1.3 土壤總DNA提取
采用OMEGA 試劑盒E.Z.N.A? Mag-Bind Soil DNA Kit 提取土壤微生物基因組總DNA,利用synergey HTY 酶標儀對已提取的核酸含量進行檢測,并通過保守區設計引物,再在引物尾端加上測序接頭,進行PCR 擴增。擴增時采用引物314F(5’-CCTACGGGNGGCWGCAG-3’)和805R(5’-GACTACHVGGGTATCTAATCC-3’)擴增細菌V3~V4區;用ITS1F(5’-CTTGGTCATTTAGAGGAAG-TAA-3’)和ITS2(5’-GCTGCGTTCTTCATCGATGC-3’)擴增真菌ITS1 區;后對擴增的細菌和真菌產物分別進行純化、定量和均一化構建文庫。已建成的文庫首先采用Qubit 3.0 定量和文庫檢測合格后,用Illumina Miseq 平臺進行測序。高通量測序所得到的原始圖像數據文件,經過堿基識別分析后轉換為原始序列(raw reads),然后對原始序列進行過濾、拼接、去除嵌合體等得到有效序列(effective reads)。
1.4 數據分析
采用SPSS 25.0 軟件進行數據整理分析。將測序原始數據去除引物接頭序列、各低質量堿基后進行拼接,舍棄長度小于200 bp 的序列,去除非特異性擴增序列及嵌合體后,得到各樣本的有效序列數據。采用Usearch 軟件進行OTUs(operational taxonomic units)聚類和物種信息分析。利用mothur 軟件進行Alpha 多樣性指數分析,包括Ace 指數、Chao 指數、Simpson 指數和Shannon指數;用R軟件繪制稀釋曲線,分析土壤微生物群落在各分類等級的相對豐度;利用Venn 圖統計各樣本中共有及特有的OTUs 數量;運用R 的gplots軟件包進行UPGMA(unweighted pair-group method using arithmetic averages)聚類分析并作樹狀圖;利用R 軟件生成主成分分析(principal component analysis,PCA)。
2 結果與分析
2.1 Cd 脅迫下地膚根際與非根際土壤理化性質、養分及總Cd含量
Cd 脅迫下,地膚根際和非根際土壤的理化性質、養分及Cd 含量如表1 所示。根際和非根際土壤的pH、水分、有機質含量分別為5.01、23.26%、21.71 g·kg-1和5.56、23.76%、19.83 g·kg-1。其中根際土壤pH 及總Cd 含量顯著低于非根際土壤;根際土壤速效磷含量顯著高于非根際土壤;其余指標在根際和非根際土壤間差異不顯著。
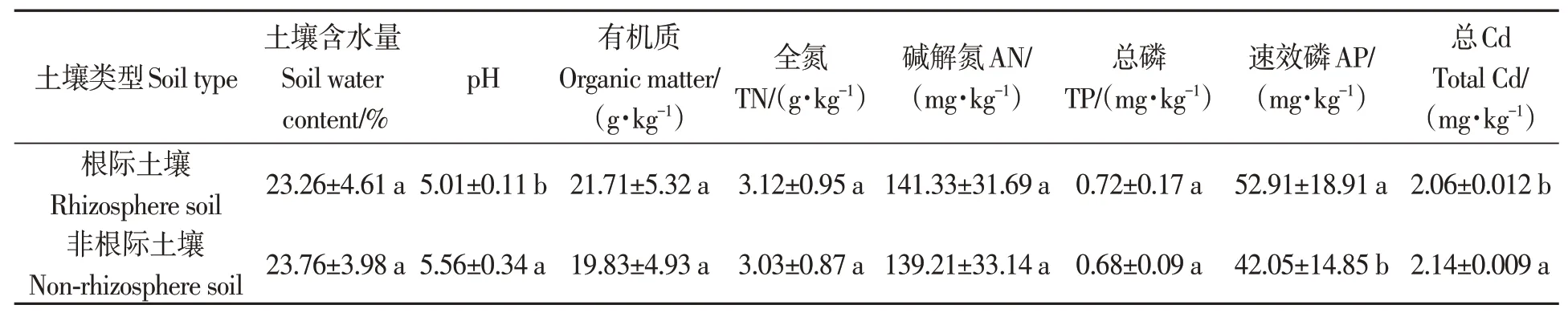
表1 鎘脅迫下地膚根際與非根際土壤理化性質、養分及Cd含量Table 1 Chemical properties, nutrients and total Cd content of rhizosphere and non-rhizosphere soil of Kochia scoparia under Cd stress
2.2 土壤微生物樣品測序結果
根際土壤細菌共得到原始序列351 307條,經拼接、質控過濾后得到有效序列349 700 條,平均序列長度412 bp;根際土壤真菌共得到原始序列233 024 條,經拼接、質控過濾后得到有效序列232 882 條,平均序列長度264 bp。非根際土壤細菌共得到原始序列305 198條,經拼接、質控過濾后得到有效序列303 796條,平均序列長度414 bp;非根際土壤真菌共得到原始序列248 453 條,經拼接、質控過濾后得到有效序列248 334 條,平均序列長度261 bp。稀釋曲線(圖1)表明,當細菌OTUs 數量達到1 500 左右,真菌OTUs 數目達到500 左右,稀釋曲線趨于平穩,隨著數據量增大僅產生較少的新物種,說明此時測序量已基本覆蓋土壤微生物類群,試驗結果科學可靠。
圖1 土壤樣品的稀釋曲線Fig. 1 Dilution curves of the soil samples
2.3 鎘脅迫下地膚根際與非根際土壤微生物群落多樣性和豐富度
2.3.1Alpha 多樣性指數 由表2 可知,地膚非根際土壤細菌的Simpson 指數大于根際土壤;而Shannon 指數小于根際土壤,因地膚非根際和根際土壤細菌的物種豐富度較接近,而均勻度差別較大,采用Simpson指數的效果更好[29],由此表明,地膚非根際土壤細菌的群落多樣性高于根際土壤;非根際土壤細菌的Chao1 指數和ACE 指數均大于根際土壤,表明非根際土壤細菌的豐度高于根際土壤,但差異不顯著。地膚非根際土壤真菌的Shannon 指數和Simpson 指數均大于根際土壤,表明非根際土壤真菌的群落多樣性高于根際土壤,但差異不顯著;非根際土壤真菌的Chao1指數和ACE指數均大于根際土壤,表明非根際土壤真菌的豐度高于根際土壤,但均差異不顯著。此外,在地膚根際和非根際土壤中,除Simpson 指數外,土壤細菌的Shannon 指數、Chao1 指數、ACE 指數均高于真菌,說明地膚根際和非根際土壤中細菌的群落多樣性和豐度均高于真菌。無論是細菌還是真菌,根際土壤的Simpson指數均小于非根際土壤,說明非根際土壤的生物多樣性高于根際土壤。
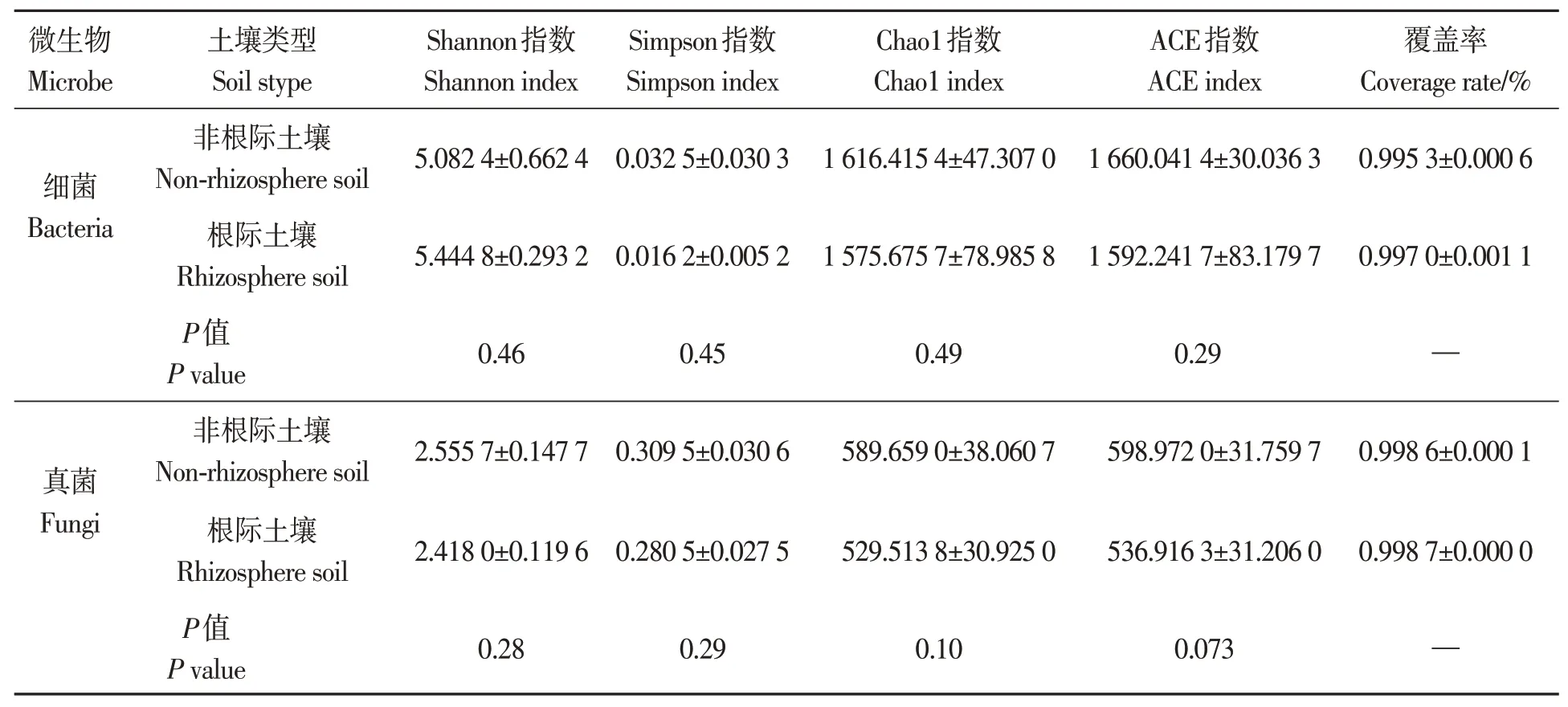
表2 Cd脅迫下地膚根際土壤與非根際土壤中細菌和真菌群落的Alpha多樣性Table 2 Alpha index of the bacterial and fungal communities in Schrad.rhizosphere and non-rhizosphere soil of Kochia scopariaunder Cd stress
2.3.2OTUs豐度 從圖2可知,Cd脅迫下根際與非根際土壤細菌共有OTUs 為1 727 個,其中根際土壤特有OTUs 為829 個,非根際土壤特有OTUs為253 個,分別占總OUT 數量的32%和12%。由此可見,Cd 脅迫下根際土壤細菌特有的OTUs 數目顯著高于非根際土壤,這說明根際土壤的細菌種類明顯較多。Cd 脅迫下,地膚根際和非根際土壤真菌共有OTUs 為593 個,其中非根際土壤特有OTUs 為215 個,根際土樣特有OTUs 為148 個,即非根際土壤真菌特有的OTUs 數量高于根際土壤。綜上,Cd 脅迫下地膚根際和非根際土壤細菌和真菌的共有OTUs 數量明顯多于特有OTUs 數量,這說明Cd脅迫下地膚根際與非根際土壤細菌和真菌的多樣性相似度均較高。
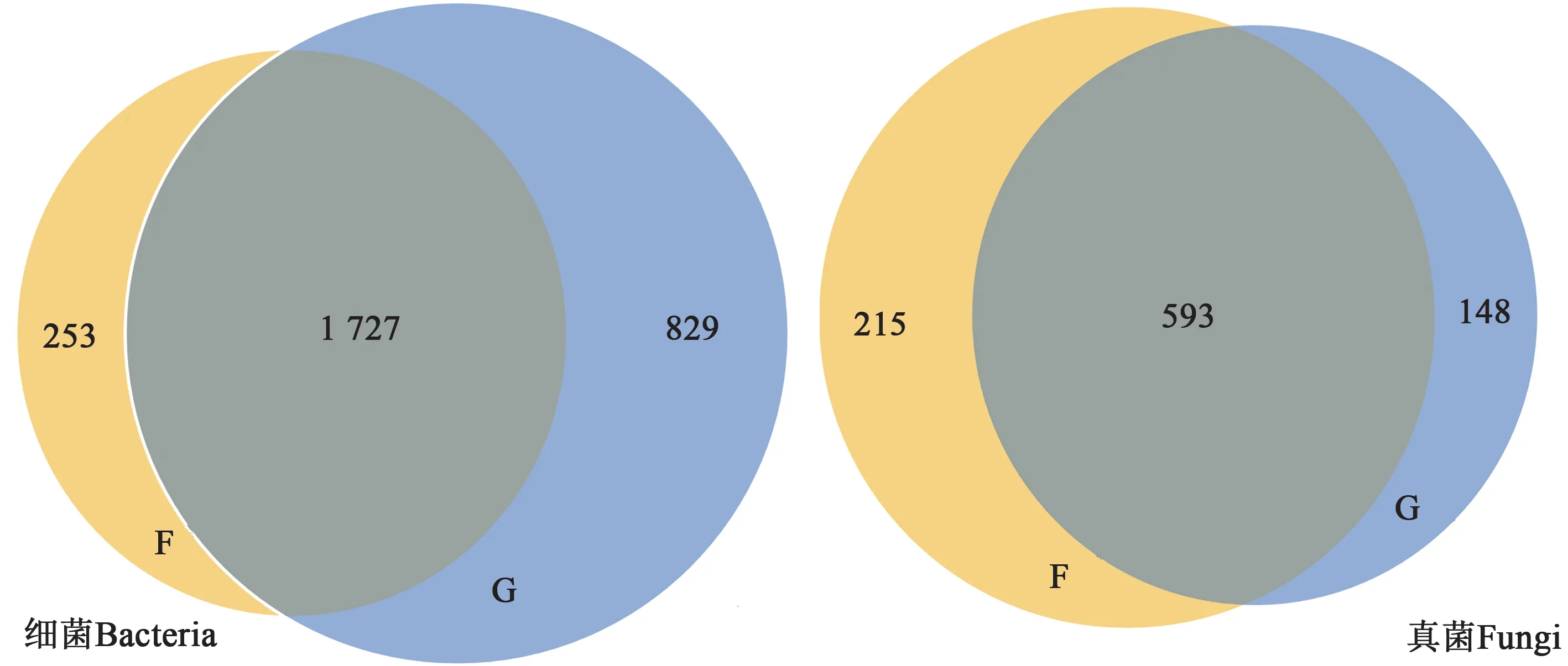
圖2 Cd脅迫下地膚根際和非根際土壤中細菌和真菌的OTUs分布Fig. 2 OTUs of bacteria and fungal in rhizosphere and non-rhizosphere soil of Kochia scoparia under Cd stress
2.4 鎘脅迫下地膚土壤微生物群落結構
2.4.1地膚根際和非根際土壤微生物的群落組成 由圖3 可知,在細菌門水平上,地膚根際和非根際土壤的主要優勢菌門均為綠彎菌門(Chloroflexi)、變形菌門(Proteobacteria)、擬桿菌門(Bacteroidetes)、厚壁菌門(Firmicutes)及其他未知菌,分別占細菌總量的83.22%和70.01%。酸桿菌門(Acidobacteria)、芽單胞菌門(Gemmatimonadetes)、candidate_division_WPS-1、Candidatus_Saccharibacteria 為根際土壤特有細菌門。變形菌門、綠彎菌門在根際和非根際土壤中的相對豐度分別為42.61%、52.41%。在真菌門水平,地膚根際和非根際土壤的主要優勢菌門均為子 囊 菌 門(Ascomycota) 、擔 子 菌 門(Basidiomycota)、壺菌門(Chytridiomycota)、被孢霉門(Mortierellomycota),分別占真菌總量的97.91%和98.55%,其中子囊菌門(Ascomycota)的相對豐度最高,分別為88.78%和83.88%。擔子菌門在根際土壤中的相對豐度高于非根際土壤6.61%。整體來說,Cd 脅迫下地膚根際土壤中的優勢微生物種群的相對豐度大于非根際土壤。
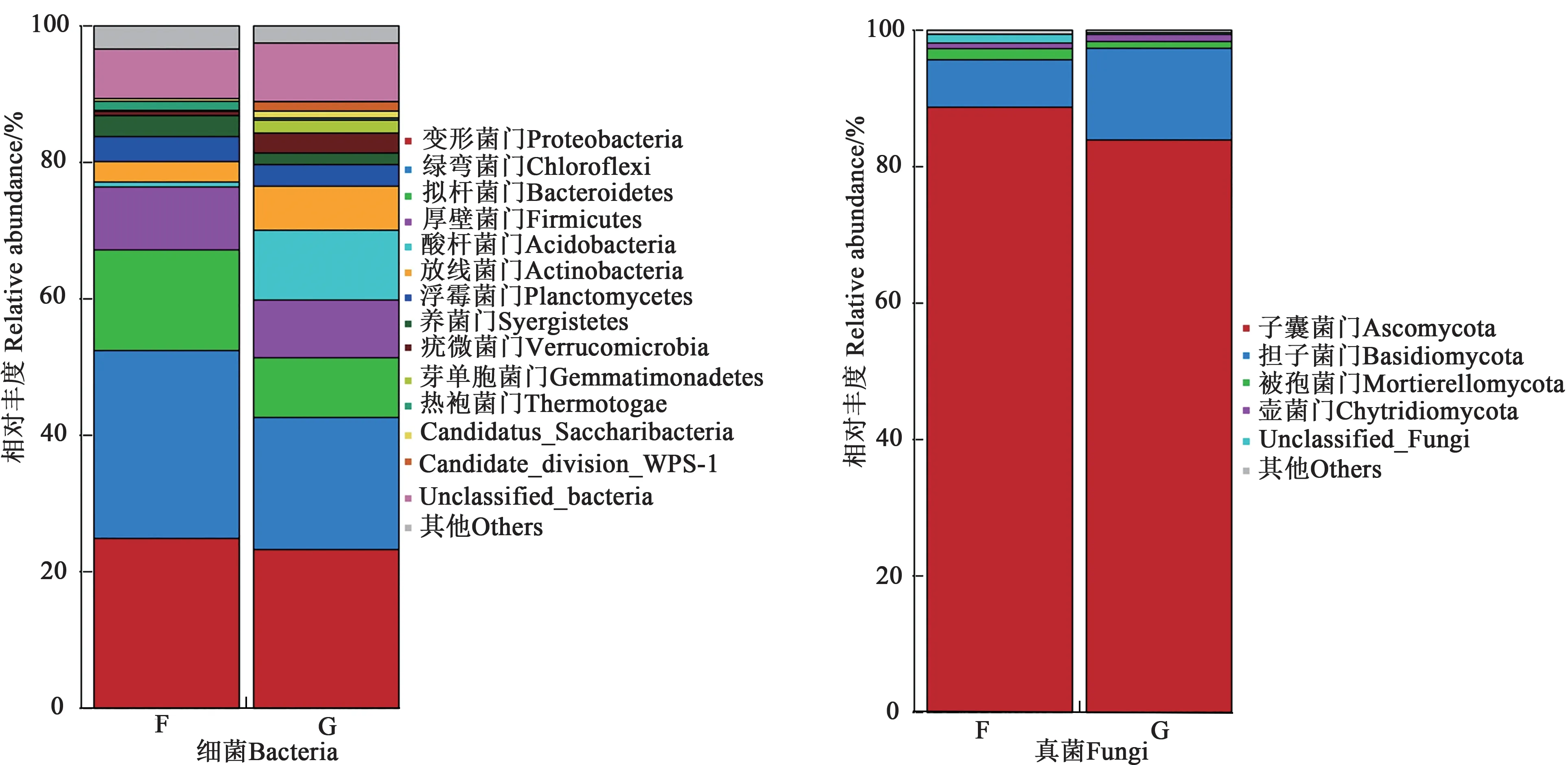
圖3 地膚根際與非根際土壤在門水平上的微生物群落組成Fig. 3 Composition of soil microbial communities in rhizosphere and non-rhizosphere of Kochia scoparia at phylum level
由圖4可知,Cd脅迫下,地膚非根際土壤細菌的主要優勢菌屬為unclassified_Anaerolineaceae、氫噬胞菌屬(Hydrogenophaga)、unclassified_Bacteroidales、未分類菌屬、unclassified_Planctomycetaceae;根際土壤的主要優勢菌屬為unclassified_Anaerolineaceae、未分類菌屬、unclassified_Bacteroidetes、unclassified_Sphingomonadaceae。其中,unclassified_Anaerolineaceae在非根際和根際土壤中的相對豐度均較高,分別為23.28%和17.32%;共有的4 個優勢菌屬分別占非根際和根際土壤細菌總量的40.84% 和33.08%;unclassified_Sphingomonadaceae為根際土壤特有菌屬。地膚非根際土壤真菌的主要優勢菌屬為緣刺盤菌屬(Cheilymenia)、鐮刀菌屬(Fusarium)、念珠菌屬(Candida)、unclassified_Pyronemataceae、unclassified_Sebacinaceae;根際土壤真菌主要優勢菌屬為緣刺盤菌屬、鐮刀菌屬、斜蓋傘屬(Clitopilus)、念珠菌屬、擬盤多毛孢屬(Neopestalotiopsis)。其中,緣刺盤菌屬在非根際和根際土壤中的相對豐度均較高,分別為54.90%和50.39%。由此說明,Cd脅迫下地膚根際和非根際土壤細菌和真菌種群和豐度在屬水平上存在差距。
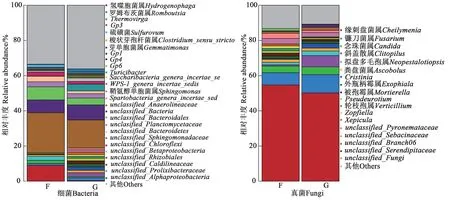
圖4 地膚根際與非根際土壤在屬水平上的微生物群落組成Fig. 4 Composition of soil microbial communities in rhizosphere and non-rhizosphere of Kochia scoparia at genus level
對根際和非根際土壤間進行比較,結果(圖5)表明,在細菌群落中,乳酸桿菌屬(Lactobacillus)和芽孢桿菌屬(Bacillus)在根際土壤中的相對豐度顯著高于非根際土壤;在真菌群落中,unclassified_Sordariomycetes、擬盤多毛孢屬、unclassified Botryosphaeriaceae、厚垣輪枝孢菌屬(Metacordyceps)在根際土壤中的相對豐度顯著高于非根際土壤;柄孢殼菌屬(Podospora)、Plectosphaerella、Gibellulopsis、 彎 孢 霉 屬(Curvularia) 、Gongronella、Myxocephala、維希尼克氏酵母菌屬(Vishniacozyma)、Pseudeurotium在非根際土壤中的相對豐度顯著高于根際土壤。
由聚類分析(圖6)可知,非根際土壤細菌群落中的優勢菌屬主要集中于unclassified_Anaerolineaceae、未分類菌屬、unclassified_Bacteroidales、氫噬胞菌屬(Hydrogenophaga)、硫磺屬(Sulfurovum)等11個屬;根際土壤細菌群落中優勢菌屬主要集中于芽單胞菌屬(Gemmatimonas)、Gp1、Gp3、unclassified_Sphingomonadaceae、unclassified_Betaproteobacteria等16 個屬。其中鞘氨醇單胞菌屬(Sphingomonas)、Gp3、WPS-1_genera_incertae_sedis等在根際土壤中的相對豐度顯著高于非根際土壤,說明非根際土壤與根際土壤細菌群落組成差異較大。
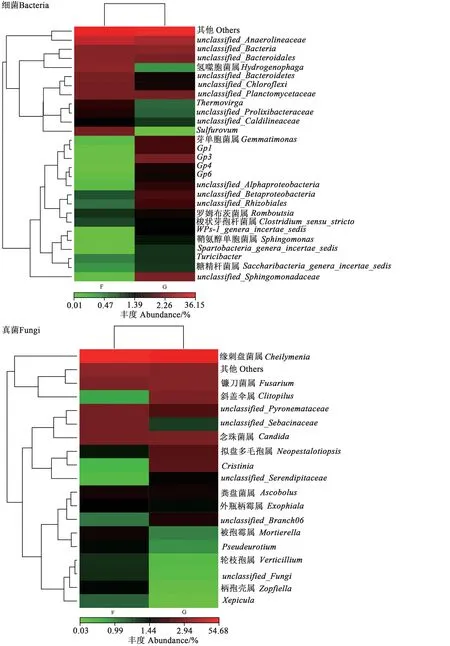
圖6 地膚根際與非根際土壤微生物群落聚類熱圖Fig. 6 Cluster heat map of microbial communities of rhizosphere and non-rhizosphere soil of Kochia scoparia
非根際土壤真菌群落中的優勢菌屬主要集中于緣刺盤菌屬、鐮刀菌屬、念珠菌屬、unclassified_Pyronemataceae、unclassified_Sebacinaceae等6 個屬;根際土壤真菌群落中的優勢菌屬主要集中于緣刺盤菌屬、鐮刀菌屬、斜蓋傘屬、unclassified_Pyronemataceae、念珠菌屬、擬盤多毛孢屬、Cristinia等8個屬(圖6)。由此說明,非根際與根際土壤的真菌群落組成存在較大差異。
2.4.2PCA分析 PCA分析可對原有的復雜數據進行降維,揭示隱藏在復雜數據背后的結構[30]。通過分析不同樣本的微生物群落組成可以反映樣本間的差異性及相似性,如圖7 所示。對于細菌群落結果,主成分1 和主成分2 的貢獻率分別為62.51%和23.96%,即分別解釋了細菌群落結構62.51%和23.96%的變異;地膚根際和非根際土壤的細菌群落在組內分布均較為分散,組間差異性較大,即根際和非根際土壤細菌群落的相似性較低。對于真菌群落組成,主成分1 和主成分的貢獻率分別為41.81%和23.55%;地膚非根際土壤不同重復間距離較近,群落組成相似,而根際土壤不同重復間較為分散,說明群落組成差異較大。上述結果表明,地膚根際和非根際土壤微生物的群落結構有較大差異。
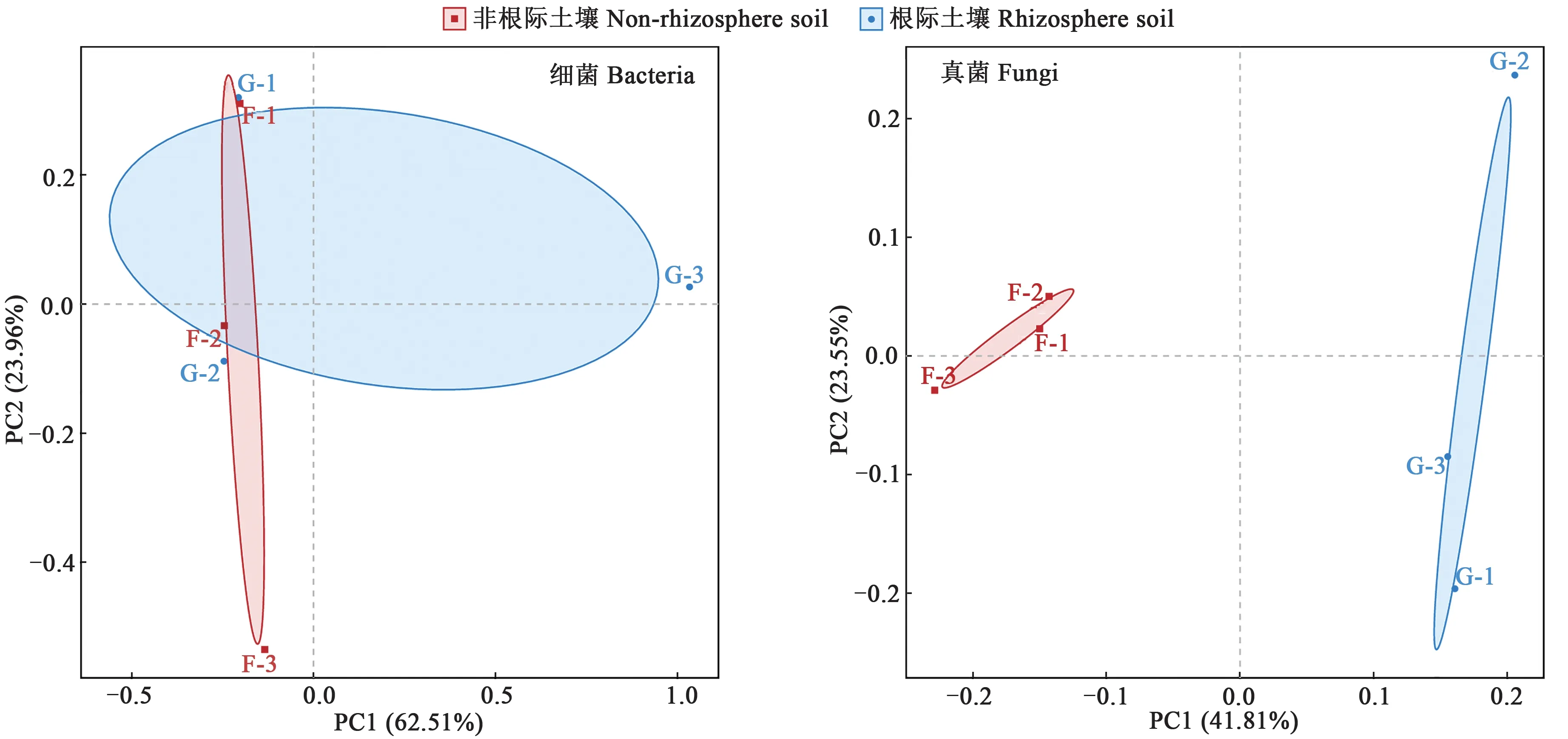
圖7 地膚根際土壤與非根際土壤微生物群落結構主成分分析Fig. 7 Principal component analysis of microbial community structure in rhizosphere soil and non-rhizosphere soil of Kochia scoparia
3 討論
根際是植物-土壤-微生物與其環境條件相互作用最為活躍的場所。根際微生物可通過促進礦物礦化、催化土壤生物化學過程等方式提高土壤中植物可利用形態營養元素的含量及植物對逆境的適應性[31]。植物根際土壤存在大量具有植物促生特性的菌群,對植物的生長、耐受逆境脅迫等方面起重要作用[32]。因此,研究植物根際土壤微生物多樣性的變化有利于揭示植物的生長發育規律,對植物尤其是用作重金屬污染土壤修復的植物十分有意義。目前,利用高通量測序技術研究植物根際土壤微生物多樣性已有較多報道。Liu等[33]研究了三葉植物修復過程中根際土壤微生物的群落結構和組成,發現在修復后,其Shannon 指數上升,Simpson 指數下降,表明種植三葉植物提高了土壤微生物群落的豐富度和均勻性,且修復后的微生物—植株系統對重金屬的抗性得到增強。金裕華等[34]研究了在重金屬污染環境下木本植物根際微生物群落多樣性,發現其修復區土壤細菌、放線菌及總菌群數量均顯著高于對照區土壤,而對照區土壤的真菌數量顯著高于木本植物修復區土壤;且木本植物修復區土壤微生物群落種類多、分布均勻,利用木本植物修復土壤可提高土壤微生物群落功能多樣性。卞方圓等[35]研究了重金屬復合污染下毛竹純林、毛竹與伴礦景天間作林2 種不同種植模式對土壤微生物群落的影響,發現毛竹純林土壤微生物的Shannon指數和Chao指數最高,表明種植修復植物有利于增加土壤細菌群落多樣性。本研究通過對地膚根際和非根際土壤細菌16S rRNA V3~V4 序列和真菌 ITS1~ITS2 區進行測序,共獲得4 536 個細菌OTUs、1 549 個真菌OTUs。其中,根際土壤細菌和真菌檢測到2 556和741 個OTUs,非根際土壤細菌檢測到1 980 和808 個OTUs;根際細菌和真菌特有的OTUs 占比分別為32.43%和19.97%,非根際細菌和真菌特有OTU 占比分別為12.78%和26.61%。由此表明,根際土壤細菌擁有較多特有的OTUs,非根際土壤真菌擁有較多獨有的OTUs;且地膚根際土壤的微生物多樣性高于非根際土壤,這與吳秋芳等[36]研究結果一致。
本研究中,地膚根際和非根際土壤細菌隸屬于35門、83綱、131目、249科、473屬。在門水平上,根際與非根際土壤細菌共有的菌門主要是綠彎菌門、變形菌門、擬桿菌門、厚壁菌門、浮霉菌門、放線菌門、養菌門、疣微菌門和降氨酸菌門。Kaplan 等[37]研究發現,上述這些細菌也存在于重金屬污染的農田土壤中。其中綠彎菌門、變形菌門、擬桿菌門和厚壁菌門在地膚根際和非根際土壤中均占主導地位,表明它們是根與微生物相互作用正反饋的主要促進因素,這與Yang 等[38]的研究結果一致。這一現象可能與細菌的生命策略(富營養、貧營養或兼性貧營養)有關,因為富營養細菌比貧營養細菌在碳資源方面更具競爭力[39-40]。變形菌門是所有菌群中最為豐富的菌門,對土壤中有機質、氮、磷、硫的循環有顯著促進作用[41]。變形菌門是重金屬高污染環境中的主要細菌門,包括礦山沉積物、地表水和被重金屬污染的土壤[42]。綠彎菌門是土壤生態循環的重要組成部分,它能通過光合作用將光能轉化為化學能來適應貧瘠的環境,且在重金屬污染土壤中占有優勢地位[43]。放線菌門中包含許多可產抗生素、酶和有機酸等的細菌,其豐度的高低可作為土壤健康狀況的評價指標[41]。本研究發現,地膚根際土壤中放線菌門的相對豐度是非根際土壤的2倍左右,這表明放線菌在重金屬污染土壤修復中起重要作用。放線菌對植物生長及營養獲取、重金屬解毒、減輕生物和非生物脅迫起重要作用[44]。由于對重金屬具有抗性,放線菌總是存在于重金屬污染的土壤中,此菌群可以取代高敏感性真菌作為土壤中的主要分解者[45]。本研究表明,酸桿菌門、芽單胞菌門、candidate_division_WPS-1、Candidatus_Saccharibacteria 為根際土壤特有細菌門。Barns 等[46]研究證明,酸桿菌門、浮霉菌門、candidate_division_WPS-1 是典型的慢生長型貧營養細菌,酸桿菌門不僅具有抵抗滲透脅迫的能力,還具有抵抗重金屬污染和其他極端環境的能力,且可以在土壤濕度起伏大的環境中生存。據報道,芽單胞菌屬和鞘脂單胞菌屬是促進植物生長的有益細菌[47-48];Gp1、Gp3和Gp6可以幫助植物吸收養分,提高植物生產力[49]。本研究表明,芽單胞菌屬、Gp1、Gp3、unclassified_Sphingomonadaceae為根際土壤的優勢菌屬,這表明地膚根際土壤具有促進植物生長和重金屬解毒的優勢微生物種群,該結果為研究地膚在Cd 脅迫下健康生長的機理奠定了理論基礎。
地膚根際和非根際土壤真菌隸屬于13 門、47綱、116目、247科、431屬。在門水平上,根際和非根際土壤中的真菌優勢菌群主要是子囊菌門、擔子菌門、被孢霉門和壺菌門。其中子囊菌門在非根際土壤和根際土壤中均占絕對優勢,相對豐度分別為88.78%和83.88%。子囊菌是世界上最豐富和分布最廣泛的土壤真菌[50],它的繁殖能力極強,能產生大量的無性孢子,且生長迅猛,使其在數量上占顯著優勢,這可能是子囊菌在很多植物根際中占比較大的原因[51]。子囊菌是生態系統碳氮循環的重要驅動因素,其在土壤穩定性、植物生物量分解中發揮作用,還可與植物形成共生關系[52]。Lin 等[53]研究發現,子囊菌門、擔子菌門、壺菌門與重金屬污染的相關性較強,可通過增強對環境壓力的防御生存,即使在各種重金屬環境中也是如此[54],擔子菌門具有轉移重金屬的能力[53]。本研究發現,地膚根際土壤真菌中擔子菌門的相對豐度較非根際土壤提高6.61%;而根際土壤的總Cd 含量(2.06 mg·kg-1)低于非根際土壤(2.14 mg·kg-1),由此表明,擔子菌在重金屬水平較低的環境中數量顯著增加。這表明擔子菌更適合在重金屬污染水平相對較低的環境中發揮作用,與Tian 等[55]的研究結果一致。被孢霉門真菌是土壤中較為常見的真菌,具有促進植物根系吸收養分、抑制病原菌等功能[56]。在屬水平上,緣刺盤菌屬和鐮刀菌屬為地膚根際和非根際土壤真菌的2 大優勢菌屬,累計相對豐度均在60%以上。研究發現,重金屬污染區域中鐮刀菌是繼青霉菌的第二大高豐度真菌,且分離出的鐮刀菌屬真菌對培養基中金屬離子水平均十分敏感,這表明鐮刀菌屬真菌對重金屬有一定耐受性[57],但對高水平重金屬污染的抵抗力較差[58]。此外,在地膚根際與非根際土壤中,均有較大比例的未知菌群,這些未知菌群的生態學功能有待進一步研究。
本研究發現,在地膚根際土壤中促進植物生長和重金屬解毒的微生物菌群占優勢,且根際土壤微生物的多樣性高于非根際土壤,包括放線菌門、疣微菌門、降氨酸菌門、擔子菌門、壺菌門以及unclassified_Betaproteobacteria、鐮刀菌屬、斜蓋傘屬、念珠菌屬、擬盤多毛孢屬、unclassified_Branch06等,它們在根際土壤中的相對豐度高于非根際土壤。由此表明,種植地膚可以增加土壤微生物豐富度,進而增強土壤微生物群落結構的穩定性,減輕Cd污染土壤給農業生產帶來的危害。但是,由于本研究未對空白樣品(未種植地膚的Cd 污染土壤)進行分析,僅比較了地膚根際和非根際土壤微生物群落的變化情況,無法比較種植地膚對土壤微生物群落結構和多樣性的影響,雖得出了地膚根際土壤較非根際土壤中促進植物生長和重金屬解毒的微生物菌群更占優勢,但究竟是哪種優勢菌群對地膚在Cd 污染土壤中的健康生長起決定作用目前尚無法確定,有待進一步研究。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
