固態食醋自然發酵醋醅中生淀粉酶產生菌的篩選及初步應用
2023-10-10 12:43:18康雪梅羅雯郭建孫群
中國調味品 2023年10期
康雪梅,羅雯,郭建,孫群
(1.四川大學 生命科學學院,成都 610064;2.千禾味業食品股份有限公司,四川 眉山 625000)
中國傳統谷物醋是以生谷物為原料,大曲等為發酵劑,在開放式環境中發酵而成,其中涉及多菌種及多酶系的相互作用,多菌種混合發酵賦予谷物醋豐富且協調的風味與口感[1-4]。固態食醋發酵過程中,微生物菌群的代謝網絡表明曲霉(Aspergillus)、根霉(Rhizopus)、紅曲(Monascus)是食醋大曲中主要的真菌,釀酒酵母(Saccharomycescerevisiae)和漢遜酵母(Hansenulaanomala)是酒精發酵階段的主要酵母菌,醋酸菌屬(Acetobacter)、乳桿菌屬(Lactobacillus)、乳球菌屬(Lactococcus)、假單胞菌屬(Pseudomonas)、葡糖醋桿菌屬(Gluconacetobacer)和芽孢桿菌屬(Bacillus)是食醋風味物質產生的核心功能菌群,在醋的優勢風味物質形成中起著重要作用[5-7]。
選育獲取優良性能的功能微生物并進行強化生產,可促進食醋發酵生產提高生產效率,降低成本,改善品質。在食醋醋醅中添加A.pasteurianusG3-2、L.brevis4-22、L.fermentumM10-3和L.buchneriF2-5,可以原位增強食醋菌群的產乙偶姻功能[6]。在醋酸發酵開始時添加內源巴氏桿菌,可以增強鎮江香醋的風味,縮短了醋酸發酵周期,提高了總酸、2,3-丁二醇和川芎嗪的積累[8]。在保寧醋生產中,使用黑曲霉進行強化制曲,可使原料利用率及有機酸含量得到提高[9-10]。將食醋醋醅中非常豐富的物種KomagataeibactereuropaeusJNP1篩選出來并強化應用到食醋發酵中,可使食醋總酸提升14.78%,還原糖降低40.38%,促進食醋出醋率的提升[11]。
本文從自然發酵醋醅中篩選獲取同時產生淀粉酶及蛋白酶的菌株,驗證其在酸性條件下的生長性能及酶活性能,并將其初步強化應用到食醋發酵過程中,為固態食醋發酵生產的降本增效及風味改善提供了新的改善思路。
1 材料和方法
1.1 材料與試劑
千禾味業食品股份有限公司食醋生產車間發酵周期為1~24 d的自然發酵狀態下的固態醋醋醅樣品共24個,用無菌采樣袋采樣后存放于4 ℃冰箱中備用。
18種氨基酸標品(均為色譜純):購自Sigma公司;7種有機酸標品(均為色譜純):購自J &K公司、Aladdin等試劑官網;其他化學試劑:均為分析純。
1.2 培養基
富集培養基:可溶性淀粉20 g/L,酵母粉5 g/L,蛋白胨10 g/L,Na2HPO40.5 g/L,K2HPO40.5 g/L,溶劑為蒸餾水。
初篩培養基:富集培養基內添加瓊脂20 g/L。
復篩培養基:生玉米淀粉20 g/L,酵母粉5 g/L,蛋白胨10 g/L,Na2HPO40.5 g/L,K2HPO40.5 g/L,瓊脂20 g/L,溶劑為蒸餾水,其中生玉米淀粉單獨稱取到稱量瓶中,107 ℃烘箱干熱滅菌2 h后,在使用前加入到滅菌后的培養基中,混勻后傾倒平板。
醬油培養基:醬油原油50 g/L(氨基酸態氮大于1.2 g/dL),葡萄糖 20 g/L,溶劑為蒸餾水。
1.3 實驗方法
1.3.1 生淀粉酶產生菌篩選
稱取100 g醋醅樣品于1 000 mL 0.85%的生理鹽水中混合均勻并過濾得到清液,取1 mL至富集培養基中,30 ℃、150 r/min培養2~3 d,并進行3輪連續富集培養。將富集完成的菌液梯度稀釋后涂布于初篩培養基平板,35 ℃培養12~48 h后,挑選具有明顯水解透明圈生成的菌落,得到淀粉酶產生菌,進行保存。同時點種至復篩培養基平板,35 ℃培養12~48 h后,挑選透明圈直徑與菌落直徑比較大的菌落進行保存,得到生淀粉酶產生菌。
1.3.2 酶活力測定
待測菌株使用LB液體培養基活化培養48 h后,10 000 r/min、4 ℃離心10 min所得上清液即為粗酶液。
淀粉酶(或生淀粉酶)酶活定義:在40 ℃條件下,以可溶性淀粉(或玉米生淀粉)為底物,以醋酸-醋酸鈉(50 mmol/L,pH 5.8)為緩沖液,每1 min釋放1 mg還原糖(以葡萄糖計)所需酶量為一個酶活力單位,記為1 U。測定方法:取5 mL底物溶液(可溶性淀粉或玉米淀粉)于試管內,在40 ℃下保溫5 min,加入40 ℃保溫5 min的酶液 30 μL,200 r/min反應30 min,用200 μL 200 mmol/L的NaOH終止反應,以200 r/min反應5 min,得反應液。對照反應添加滅活后酶液進行相同處理。用DNS法測定反應液與對照反應液中的葡萄糖生成量,計算相應的酶活。
pH對酶活的影響測定方法:使相同濃度底物溶液溶解于不同pH的緩沖溶液中(50 mmol/L),測定并計算酶活,其中緩沖溶液為不同pH條件下的NaH2PO4-Na2HPO4(50 mmol/L)緩沖液或醋酸-醋酸鈉(50 mmol/L)緩沖液。
采用GB/T 23527—2009中福林-酚法測定中性蛋白酶及酸性蛋白酶酶活。
1.3.3 目標菌株鑒定
以目標菌株的總DNA為模板,利用引物P1:5′-AGAGTTTGATCCTGGCTCAG-3′和P2:5′-AAG-GAG
GTGATCCAGCCGCA-3′擴增菌株的16S rDNA基因,委托生工生物工程(上海)股份有限公司對該菌的16S rDNA擴增及測序,得到該菌株的16S rDNA序列后,在NCBI網站上用BLAST檢索GenBank中相關菌株的16S rDNA基因序列,并進行同源性比對,構建進化樹,確定其種屬。
1.3.4 菌株培養及耐酸性實驗
菌株培養:使用食品級LB(酵母粉 5 g/L,蛋白胨 10 g/L,NaCl 10 g/L)培養基進行種子培養及菌株生長耐酸性能驗證。使用醬油培養基進行菌株的擴大培養,得到發酵菌株培養液,用于發酵生產。
耐酸性實驗:配制LB培養基(pH 6.0~3.0,乳酸調節),滅菌后接種種子液在35 ℃、200 r/min進行培養,觀察菌株的生長狀況。
1.3.5 菌株產乙偶姻功能驗證
LB液體培養基進行液態培養48 h得到的菌株培養液,以10 000 r/min離心,分離得到上清液備用。以未接種菌株的空白培養基作為對照,采用肌酐比色法測定乙偶姻含量[6]。
3)噴嘴煙氣流速:再循環煙氣在爐內需起到充分攪拌及降低爐溫的作用,不同爐型、不同處理規模的焚燒爐對應的最低風速要求均不相同。噴嘴煙氣流速需根據爐膛流場模擬進行設計,并結合運行情況進行優化。
1.3.6 目標菌株在固態食醋釀造中的初步應用
將目標菌株用LB斜面培養后,于醬油培養基中進行種子培養,獲得發酵菌種,并進一步轉接至醬油培養基進行擴大培養,獲得發酵培養液。將獲得的發酵培養液按0.25%的接種量接種至液化糖醪液中,靜置培養過夜后,下水拌醅進行食醋發酵,每輪選擇車間3個樓層共6個發酵池作為實驗組,對照組為分別與實驗組同樓層的未接種目標菌株的正常進行下水發酵生產的食醋。發酵過程中檢測總酸、氨氮的生成情況,發酵結束后進行淋醋,所得原醋進行總酸、氨氮、有機酸、氨基酸及風味物質檢測。連續進行3輪實驗后,計算出醋率。
1.4 分析方法及數據處理
實驗結果均為3次實驗的平均值,使用GraphPad Prism 9進行數據統計分析及繪圖,P<0.05表示顯著差異。
食醋總酸、氨氮、有機酸、氨基酸、風味物質等的測定參照文獻[9,12]的方法。
2 結果與分析
2.1 生淀粉酶產生菌的篩選
2.1.1 生淀粉酶產生菌株的篩選
采用透明圈法篩選出透明圈直徑與菌落直徑比大于2.5的菌株共78株,即為產淀粉酶菌株。為滿足食醋生料發酵需要,進一步復篩得到具有透明圈且透明圈直徑與菌落直徑比大于2的菌株共16株,進行進一步的分離純化,獲得生淀粉酶產生菌,見圖1。

圖1 生淀粉酶產生菌
2.1.2 生淀粉酶產生菌酶活測定及目標菌株的確定
復篩所得生淀粉酶產生菌通過16S rDNA序列對比分析進行鑒定,結果見表1,編號為k4、k5、k6、k7、k13的菌株鑒定為巴氏醋桿菌巴氏亞種(A.pasteurianussubsp.pasteurianus),通過酶活測定,菌株均有淀粉酶、生淀粉酶及中性蛋白酶活性,除k4及k13具有較低的酸性蛋白酶活性外,其他幾株巴氏醋桿菌均無酸性蛋白酶活性。在固態食醋發酵中,巴氏醋桿菌為產酸主要功能性菌株,其同時具有產生淀粉酶活性及蛋白酶活性,將在一定程度上促進原料淀粉、蛋白的分解利用。篩選獲取的菌株除巴氏醋桿菌外,其他鑒定結果顯示均為芽孢桿菌屬(Bacillus),包括死谷芽孢桿菌(B.vallismortis)、解淀粉芽孢桿菌(B.amyloliquefaciens)、貝萊斯芽孢桿菌(B.velezensis)、暹羅芽孢桿菌(B.siamensis)、枯草芽孢桿菌堆肥亞種(B.subtilissubsp.stercoris)、中村芽孢桿菌(B.nakamurai),酶活測定結果顯示其均具有較好的生淀粉酶、淀粉酶、中性蛋白酶、酸性蛋白酶活性。
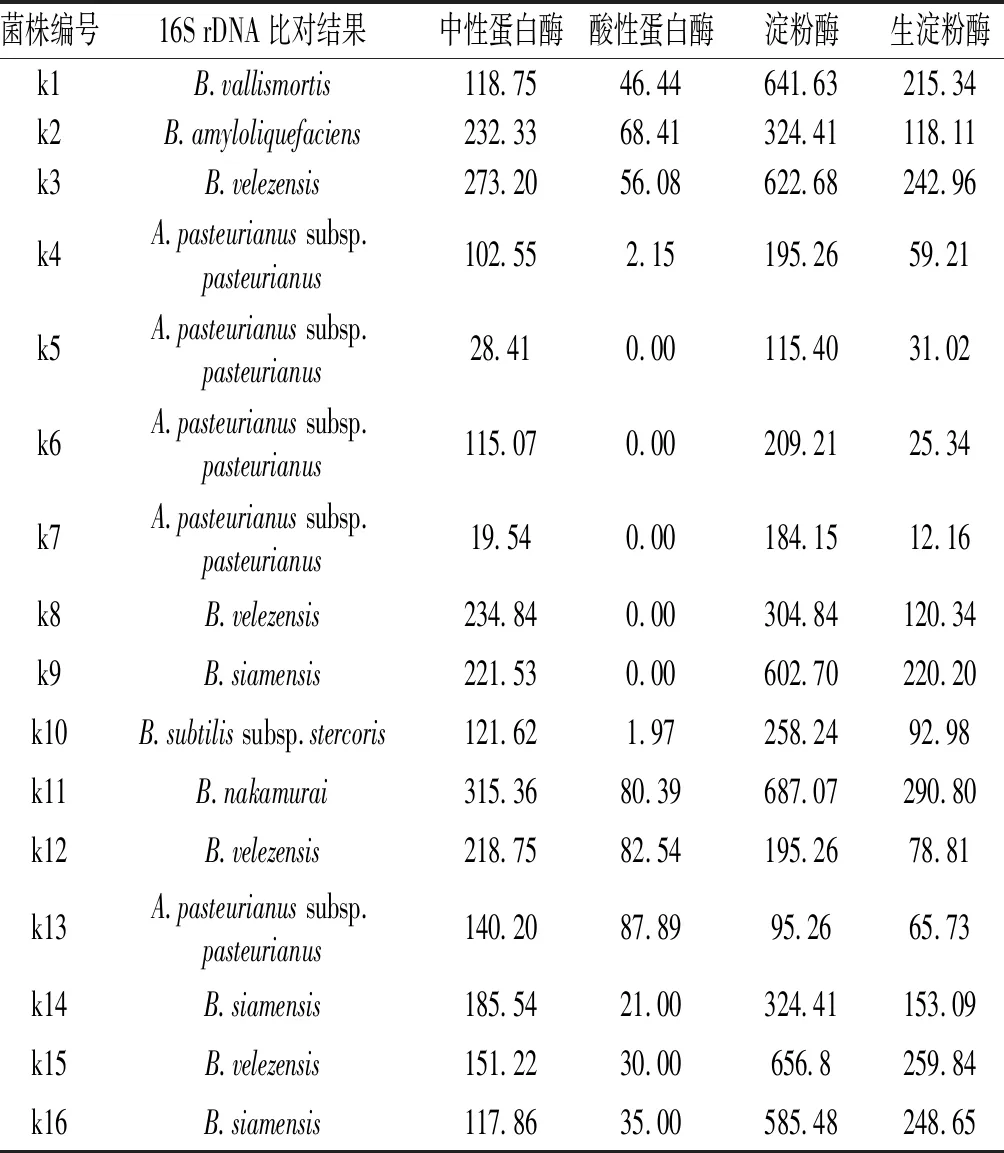
表1 菌株產酶酶活測定
芽孢桿菌抗逆性強,部分對乙酸、高溫具有耐受性,同時也被認為是一種食品級的安全菌株,具有豐富的酶系,能參與多種催化反應,具有較高的商業價值。篩選所得菌株在不同pH條件下培養的結果見表2。k4、k5、k6、k7、k13均能在酸性條件下生長,并且在pH 3.0時均能生長,接種液體培養基12 h內均可形成渾濁菌液。所有芽孢桿菌在pH 4.5條件下均能生長,在pH 4.0時,k2、k12、k15能生長,但是在接種后培養36 h內不能形成渾濁菌液,在pH 3.5時,k3、k8均不能在36 h內形成渾濁菌液,在pH 3.5及pH 3.0時,k1、k9、k16、k11均能正常生長,且能在接種培養36 h 內形成明顯的渾濁菌液。由此可知,在食醋發酵過程中,在低pH環境條件下,死谷芽孢桿菌k1、暹羅芽孢桿菌k9、k16、中村芽孢桿菌k11的生長不會受到影響,為食醋發酵中的又一種可利用的功能菌株。結合表1的酶活測定結果,由于暹羅芽孢桿菌k9缺少酸性蛋白酶酶活,死谷芽孢桿菌k1、暹羅芽孢桿菌k16、中村芽孢桿菌k11可作為優勢菌株進行進一步研究及利用。
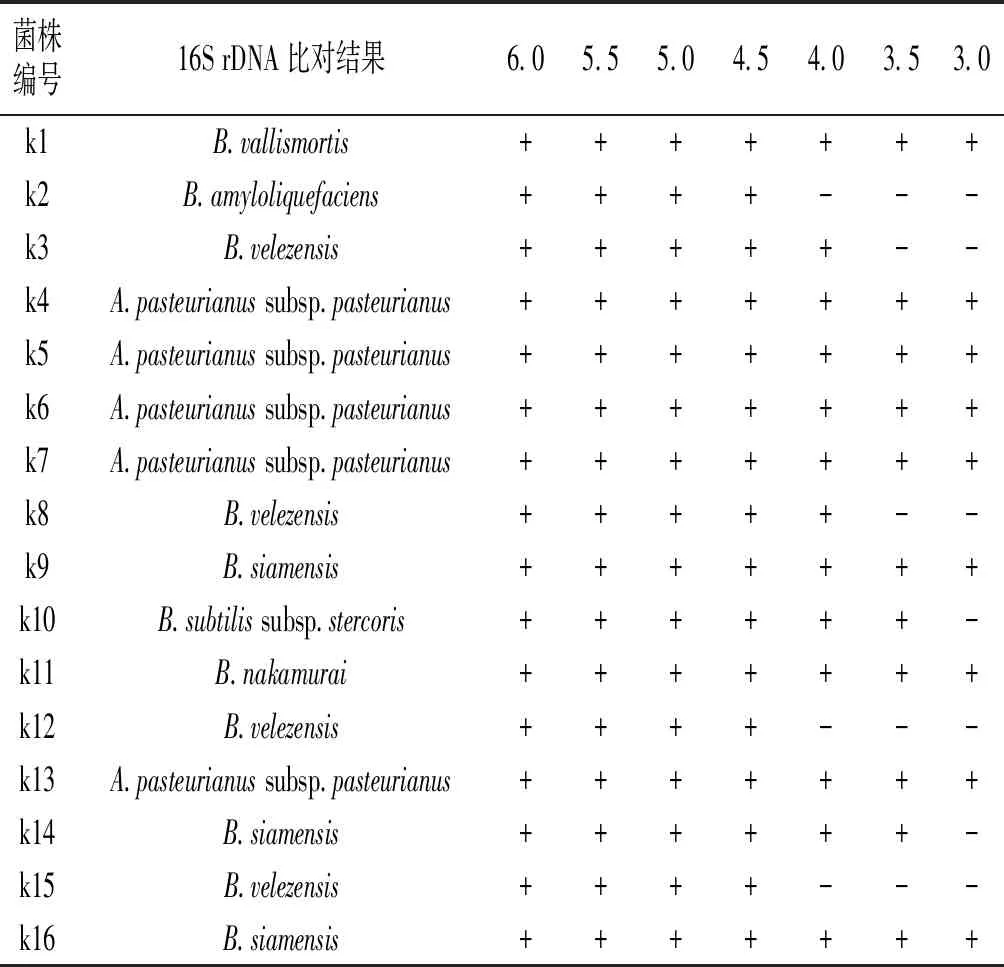
表2 pH對菌株生長的影響
2.1.3 pH對目標菌株產酶酶活的影響
考察在酸性條件下菌株產酶酶活的變化情況,進一步評估目標菌株在食醋發酵過程中的適用性。由圖2可知,k1、k16、k11所產生的生淀粉酶最佳pH分別為6.5,7.0,6.5。在堿性條件下,酶活隨著pH的升高而升高,在酸性條件下,酶活隨著pH的降低而降低,在pH 3.0時,菌株殘余酶活為最佳酶活的50%以上,其中,中村芽孢桿菌k11所產生的生淀粉酶酶活為最佳酶活的81.15%,保留了最大的酶活。死谷芽孢桿菌k1、暹羅芽孢桿菌k16的殘余酶活分別為初始酶活的52.84%及65.95%。中村芽孢桿菌k11為食醋發酵生產中的最佳生淀粉酶產生菌,可作為目標菌株進行在食醋生產中的應用實驗。
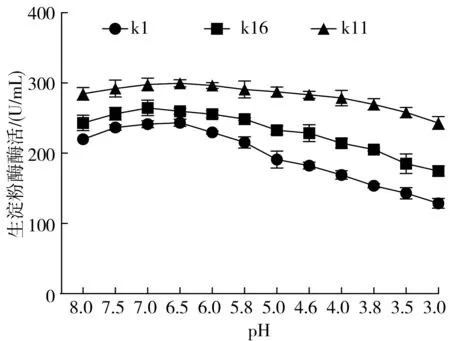
圖2 pH對酶活的影響
2.1.4 目標菌株產乙偶姻功能驗證
乙偶姻是食醋中一種重要的風味物質,能夠提供濃郁的奶油香,賦予食醋香甜美味。同時,乙偶姻也是功能性物質四甲基吡嗪的重要前體物質[13-15]。
由圖3可知,篩選獲取菌株均具有產乙偶姻功能,其中,菌株k5、k7、k13所產乙偶姻均明顯低于其他菌株,分子生物學鑒定顯示其均為巴氏醋桿菌,研究表明巴氏醋桿菌具有乙偶姻生成功能,但將其與乳酸菌進行共培養,乙偶姻的積累更明顯[6]。同時,芽孢桿菌也可促進食醋中乙偶姻的積累,如在保寧醋發酵過程中使用強化接種解淀粉芽孢桿菌的大曲,將使發酵所得食醋乙偶姻含量大幅提升[16]。所篩選菌株中,k11、k14、k15的乙偶姻含量均較高,分別為45.71,45.21,45.38 mg/L,可作為產乙偶姻功能性菌株進一步應用到固態食醋發酵過程中。結合篩選菌株在酸性條件的生長情況及其所產生淀粉酶在pH 3.0~4.0之間的酶活情況,綜合考慮,選取中村芽孢桿菌k11作為目標菌株進行固態食醋發酵的應用。
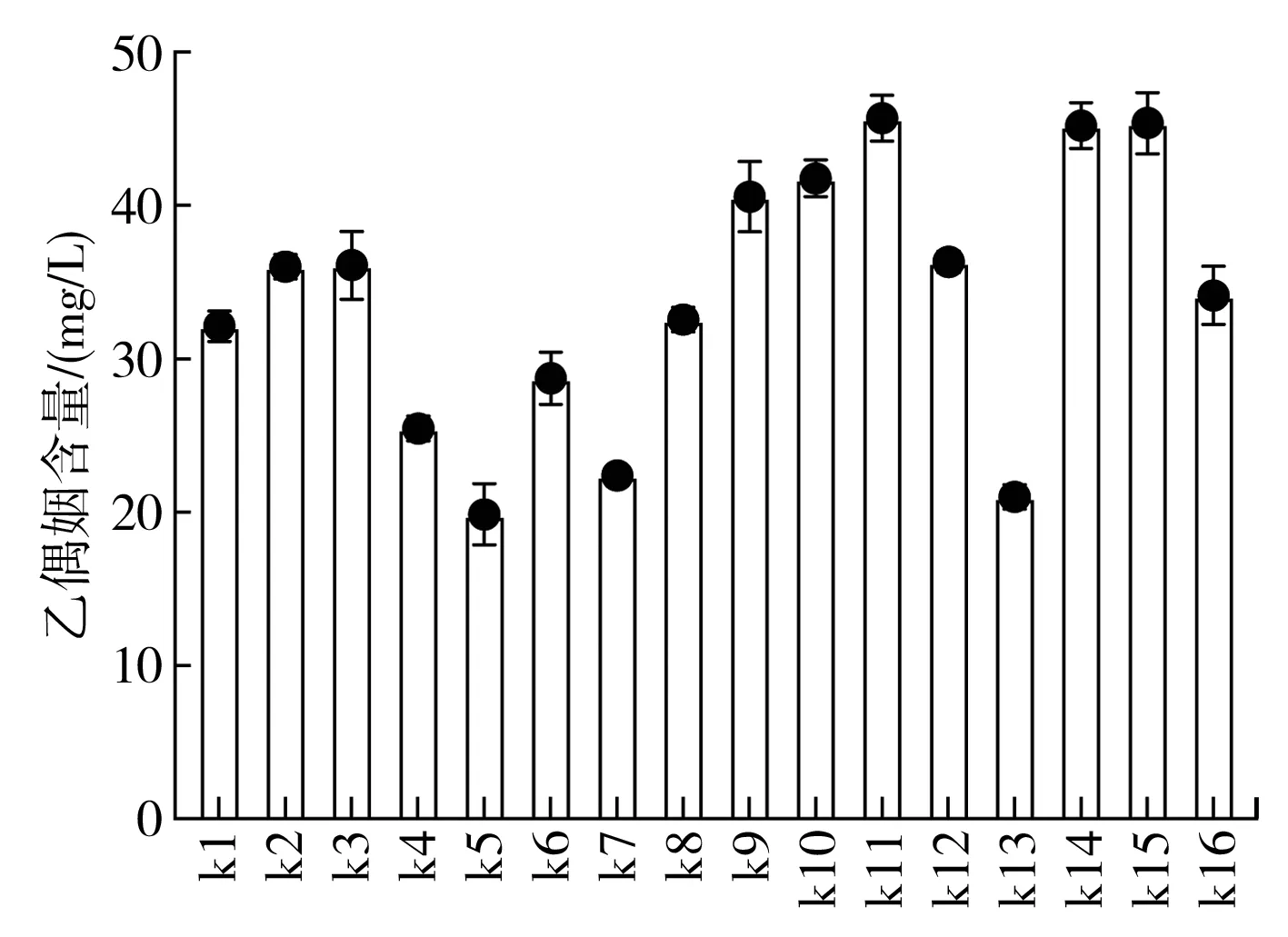
圖3 菌株產乙偶姻濃度測定
2.2 生淀粉酶產生菌在固態食醋發酵過程中的初步應用
2.2.1 發酵過程中理化指標隨發酵周期的變化情況
測定了發酵過程中發酵池底鹵汁的理化指標,結果見圖4。
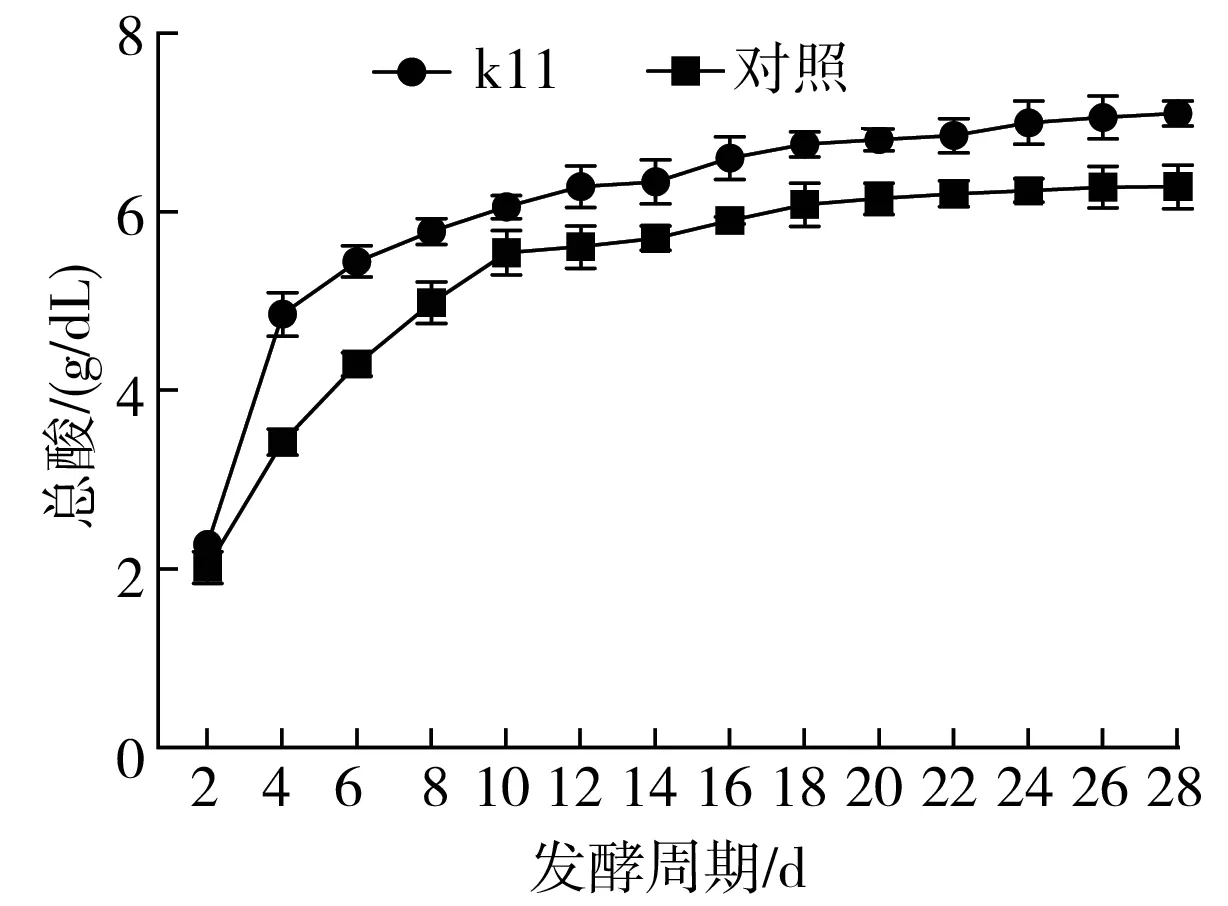
圖4 總酸、不揮發酸、氨氮、酒精度、還原糖、發酵溫度隨發酵周期的變化
實驗組與對照組在發酵過程中,總酸含量不斷升高,實驗組在發酵過程中總酸、氨氮等均高于對照組,且實驗組總酸的升高速度快于對照組,可見k11產生的淀粉酶、蛋白酶等對原料中淀粉、蛋白的水解促進了還原糖即氨基酸的生成,促使酒精發酵、醋酸發酵更快速地進行,促進總酸的生成,氨基酸的生成使氨氮的含量升高。發酵結束時實驗組的鹵汁總酸為(7.10±0.14) g/dL,對照組為(6.28±0.25) g/dL,提高了15.9%。發酵結束時發酵池鹵汁中氨氮含量明顯高于對照組,實驗組為(0.65±0.01) g/dL,對照組為(0.592±0.01) g/dL。食醋發酵過程中整體不揮發酸含量均先升高后降低,在發酵開始到接近發酵中期,鹵汁中不揮發酸濃度逐漸升高,在發酵第10~12天,不揮發酸達到最高值,隨著發酵進行,不揮發酸開始逐漸降低,但實驗組整體不揮發酸濃度均高于實驗組。
發酵開始到發酵周期第4天,實驗組與對照組鹵汁中酒精度均表現出明顯且迅速地增高,第4天之后,酒精度逐漸降低,發酵后期,酒精度緩慢下降,到發酵后期,實驗組與對照組鹵汁中酒精度均低于0.5 %。發酵過程為淀粉糖化、酒精發酵、醋酸發酵同時進行,在發酵前期,淀粉糖化與酒精發酵速度高于醋酸發酵速度,酒精含量總體呈升高趨勢,隨著醋酸菌的生長與代謝逐漸旺盛,酒精產生的速度在一定程度上降低,同時酒精被快速消耗,醋酸快速生成,隨著發酵周期的進行,淀粉糖化、酒精發酵與醋酸發酵均在一定程度上進行,保持著一種微妙的平衡關系,直至發酵結束,酒精含量降低,醋酸含量達到最大值。在發酵過程中,酶解糖化淀粉質所提供的還原糖為酒精發酵及醋酸發酵所需的底物,同時也為其他微生物菌群提供了能源,因此在整個發酵過程中,無還原糖的累積階段,還原糖一直處在生成與被消耗利用的過程中。隨著發酵的進行,酸度升高,pH降低,長期的酸性環境使大部分不耐酸的微生物死亡,僅留下耐酸高滲微生物存在于發酵后期。隨著發酵的進行,在發酵前期,酵母產酒及醋酸菌等其他微生物的生長代謝,使發酵醋醅中溫度急劇升高,通過翻醅等手段可在發酵前期將醋醅溫度控制在42 ℃以下,防止燒醅,保證發酵的正常進行,發酵中后期,隨著酸度升高,微生物代謝減弱,發酵醋醅溫度也逐漸降低,在發酵中后期維持在32~34 ℃較穩定的溫度范圍內。
2.2.2 強化菌株對食醋出醋率的影響
連續進行3輪中村芽孢桿菌k11的強化實驗,每輪6個不同樓層的發酵池為實驗組,分別與實驗組同樓層同日下水制醅的6個不接種中村芽孢桿菌k11的發酵池為對照組,每2個發酵池套淋得到一批原醋,每輪3批醋高溫滅菌后放置于同一個儲存罐內。3次強化實驗的出醋率數據見圖5。中村芽孢桿菌k11強化組3次實驗的出醋率分別為(58.50±0.63)%、(58.08±0.55)%、(58.19±0.21)%,對照組的出醋率分別為(55.50±0.06)%、(55.29±0.13)%及(53.69±1.35)%,中村芽孢桿菌k11強化實驗組3次實驗結束后通過套淋所得食醋折標(3.5 g/dL)分別為(43.92±0.45),(43.60±0.71),(43.69±0.49) kL,對照組3次實驗結束后通過套淋所得食醋折標產量分別為(41.66±0.39),(41.50±0.39),(40.31±2.26) kL,實驗組3次實驗比對照組分別增加了2.26,2.10,3.38 kL食醋,對應出醋率分別提高了5.4%、5.05%及8.40%,具有顯著效果。中村芽孢桿菌k11具有產生淀粉酶的能力,能在酸性條件下生長且產酶,所產生淀粉酶在酸性條下能保持較好的酶活,在食醋發酵生產中,發酵開始時強化接種中村芽孢桿菌k11,在淀粉質原料所組成的醋醅環境中,優于淀粉、蛋白質等的存在,促進中村芽孢桿菌進一步生長繁殖并產生生淀粉酶、蛋白酶等,作用于原料淀粉及蛋白質,促進淀粉、蛋白質的分解,同時生成的還原糖及氨基酸等又進一步作為碳源、氮源促進微生物細胞的生長繁殖,促進淀粉糖化、酒精發酵、醋酸發酵的進一步進行,提高最終的出醋率,使原料的利用率進一步提高。
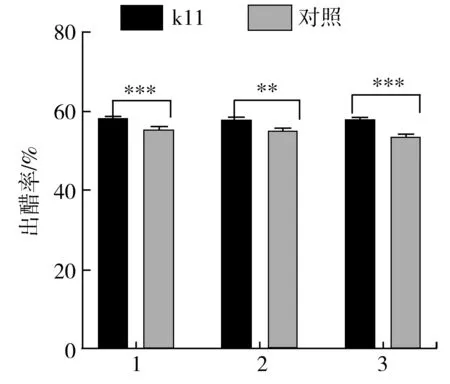
圖5 3輪實驗出醋率分析
2.2.3 食醋有機酸、氨基酸含量分析
3次實驗分別淋取得到3批實驗組原醋,對應酸度分別為(6.67±0.23),(6.23±0.45),(6.44±0.28) g/dL,各22 kL,3批對照組原醋酸度分別為(6.66±0.43),(6.18±0.53),(6.35±0.53) g/dL,各22 kL。取第1次實驗對應的實驗組與對照組原醋,分別測定其中有機酸與氨基酸組成與含量,所得結果見圖6。
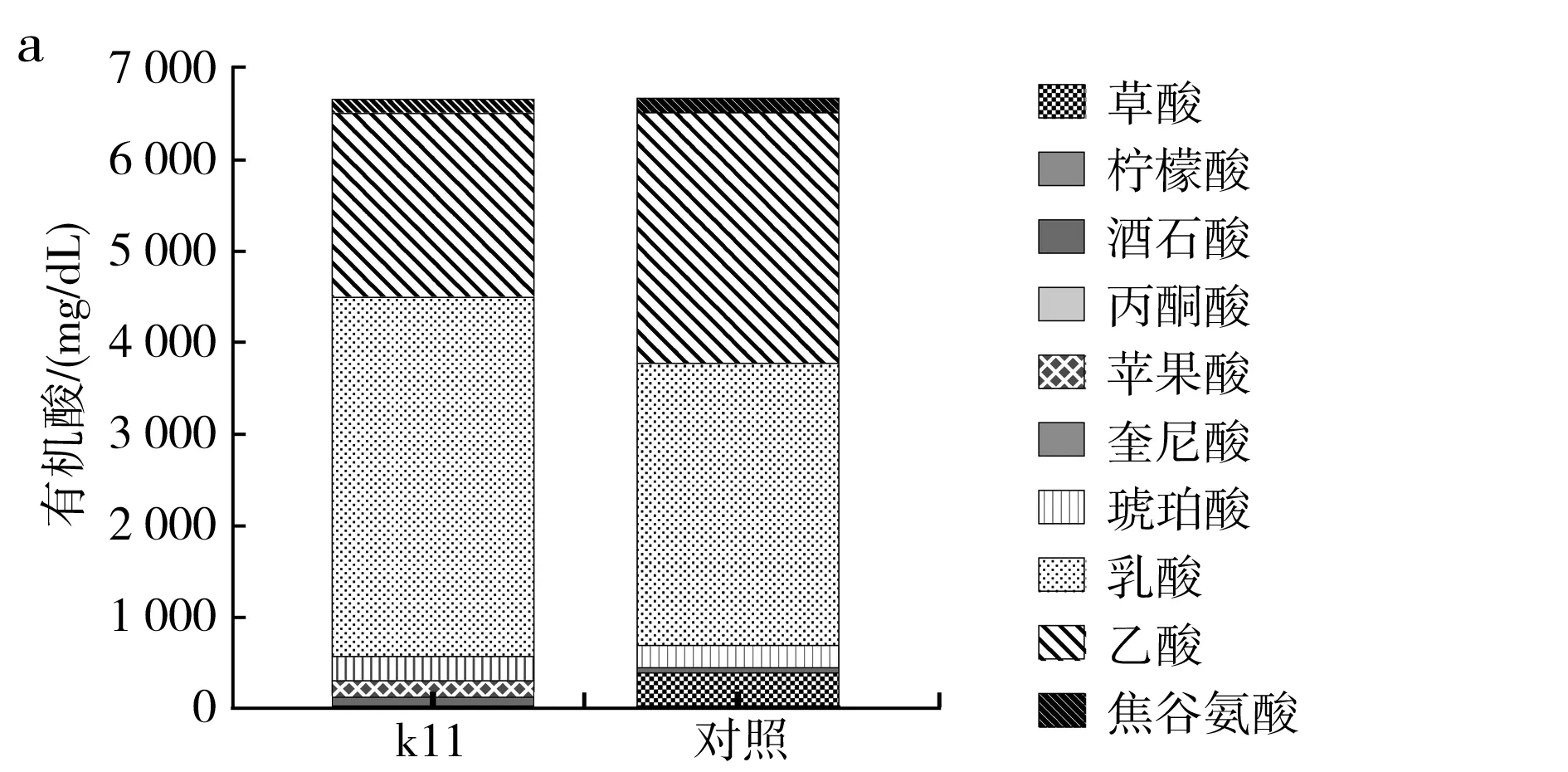
圖6 食醋中有機酸(a)、氨基酸(b)組成及含量分析
由圖6中a可知,中村芽孢桿菌k11強化實驗組乳酸含量明顯高于對照組,乙酸含量明顯低于對照組,且實驗組檢測出了蘋果酸,對照組未檢出蘋果酸,實驗組中草酸含量明顯低于對照組,其他有機酸含量基本保持不變。乳酸、蘋果酸等不揮發酸占比升高,能夠降低食醋的刺激味,使食醋的口感更柔和[2,17]。由圖6中b可知,實驗組總氨基酸含量明顯高于對照組,其中,天冬氨酸、絲氨酸、精氨酸、甘氨酸、蘇氨酸、甲硫氨酸、異亮氨酸、色氨酸、苯丙氨酸和酪氨酸在總氨基酸中的占比基本保持不變,強化接入中村芽孢桿菌k11后,谷氨酸、丙氨酸、纈氨酸、亮氨酸含量明顯降低,賴氨酸含量明顯升高,對照組未檢出半胱氨酸,但強化中村芽孢桿菌實驗組半胱氨酸含量為(214 9±5.76) mg/L,占總氨基酸含量的9.51%。可見中村芽孢桿菌強化接入醋醅后,中村芽孢桿菌所產蛋白酶作用于原料大米、麩皮等,促進了原料中蛋白質分解,中村芽孢桿菌蛋白酶中有作用于半胱氨酸與其他氨基酸所形成的肽鍵基團,使肽鍵水解,釋放出半胱氨酸,同時,與如谷氨酸、丙氨酸、纈氨酸、亮氨酸的相應肽鍵的親和力不如醋醅中其他微生物所釋放的蛋白酶,從而使食醋中谷氨酸、丙氨酸等氨基酸在總游離氨基酸中的占比降低[18]。
2.2.4 食醋風味物質的比較分析
取第一次實驗對應的實驗組與對照組原醋,使用GC-MS進行風味物質掃描分析,所得結果分別按風味物質類別進行統計分析,結果見圖7。
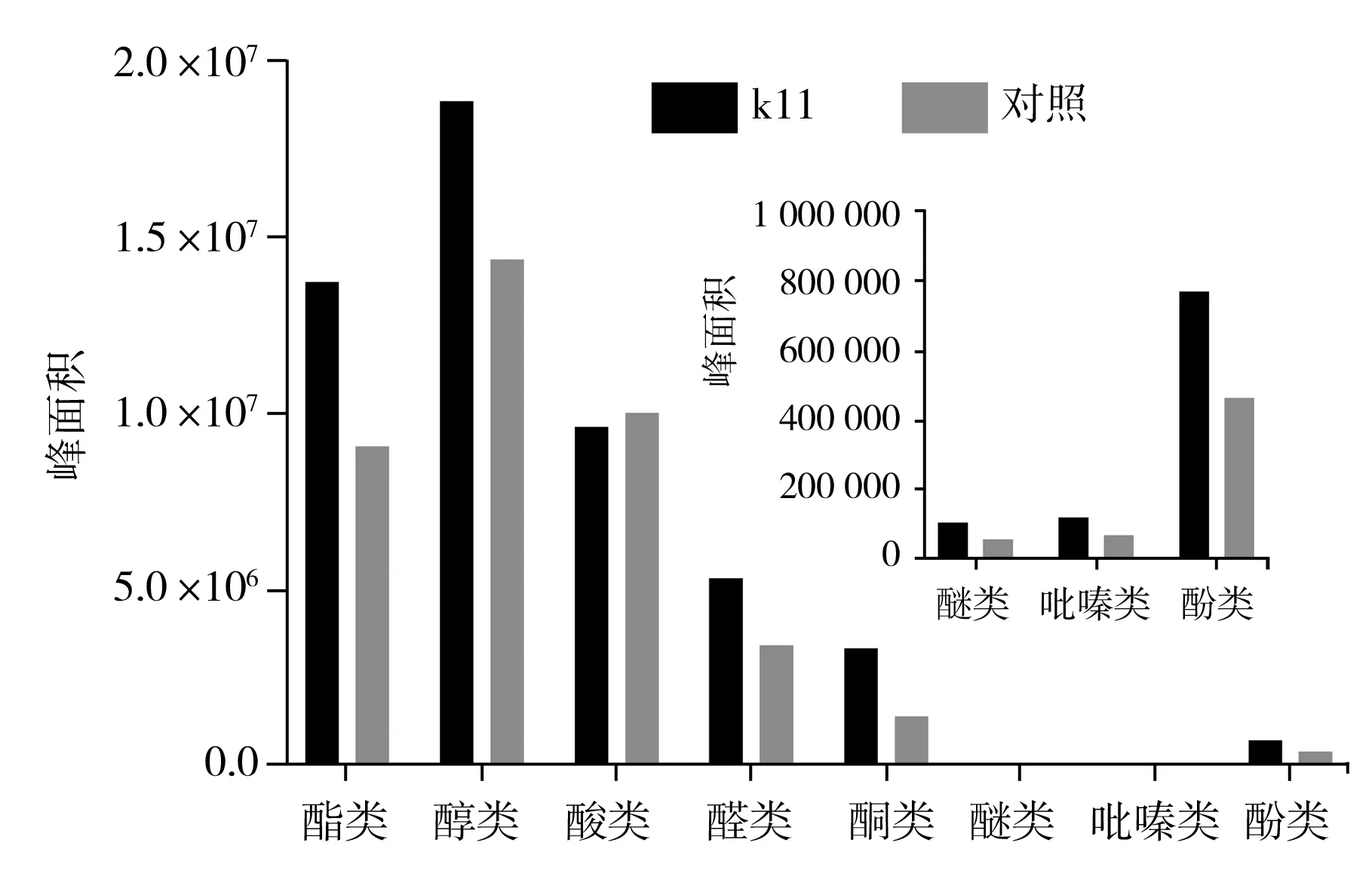
圖7 食醋中風味物質組成分析
由圖7可知,中村芽孢桿菌k11強化組風味物質的總峰面積遠高于對照組,其中,酯類、醇類、酮類、酚類含量明顯增高,酸類含量比對照組偏低。由于中村芽孢桿菌具有產乙偶姻功能,且乙偶姻為食醋中較重要的酮類物質,因此,酮類明顯增加,又因在微生物代謝中,乙偶姻可以與2,3-丁二醇相互轉化,同時也與雙乙酰含量相關,因此,酮類物質、醇類物質總含量均高于對照組[19-20]。由于乙偶姻是四甲基吡嗪的重要前體物質,因此,在中村芽孢桿菌k11強化組中吡嗪類物質顯著高于對照組。將中村芽孢桿菌k11應用到固態食醋釀造過程中,對豐富食醋風味物質具有積極作用。
3 結論
本研究從自然發酵醋醅中篩選獲得生淀粉酶、蛋白酶產生菌,鑒定為中村芽孢桿菌,編號為k11,此菌株能在酸性條件下生長并產酶,所產生淀粉酶在酸性條件下能保存80%以上的酶活,并具有高產乙偶姻功能。將此菌株在固態食醋發酵生產中進行初步應用,在發酵過程中,池底鹵汁的總酸、氨氮、不揮發酸含量等均有明顯提高,通過3輪重復應用實驗,可使食醋出醋率分別提高5.40%、5.05%及8.40%,可在一定程度上降低生產成本。所得食醋乳酸、蘋果酸占比提升,草酸含量降低,可使食醋的口感柔和。游離氨基酸總量提升了71.52%,且增加了其中半胱氨酸的含量,增加了食醋的酸味,且食醋中醇類、酯類、酮類、吡嗪類、酚類物質的含量均有明顯提升。本研究結果表明,將中村芽孢桿菌應用到固態食醋發酵過程中,可提高出醋率,降低生產成本,并在一定程度上提升食醋的口感,對于提升傳統發酵調味品的生產效率及產品品質具有重要意義。
