小麥新品系的赤霉病抗性及分子標記分析
2023-10-11 08:50:38王紫檀李浩陽趙文莎宋鵬博孫道杰張玲麗
麥類作物學報 2023年10期
王紫檀,李浩陽,趙文莎,宋鵬博,孫道杰,馮 毅,張玲麗
(西北農林科技大學農學院,陜西楊凌 712100)
小麥赤霉病(Fusarium head blight, FHB)是由禾谷鐮孢菌(FusariumgraminearumSchw.)等引起的一種真菌病害。禾谷鐮孢菌在小麥開花期侵染穗部小花,在籽粒灌漿成熟過程中沿穗軸擴展,產生并積累脫氧雪腐鐮孢菌烯醇(deoxynivalenol, DON)等毒素,致使籽粒干癟,嚴重時整穗死亡,對小麥的產量和品質造成負面影響;被毒素污染的籽粒還嚴重威脅人畜健康[1-4]。小麥赤霉病的高發區過去主要集中在長江中下游冬麥區和東北春麥區,近年來伴隨氣候變化和耕作方式的改變,發病區域逐步延伸至黃淮麥區[5]。生產上,防治赤霉病主要采用化學防治和種植抗病品種的方法,化學防治會造成嚴重的環境污染,因此種植抗病品種是防治小麥赤霉病最經濟有效的措施[3]。在生產上,能直接利用的抗赤霉病小麥品種較少,選育抗赤霉病品種是當前黃淮麥區小麥育種的重要任務之一[6]。
小麥赤霉病抗性是由多基因控制的數量性狀,目前正式命名的抗病基因有Fhb1~Fhb7,只有Fhb1[7]和Fhb7[8]被克隆,其中Fhb1被證實是效應最強且最穩定的抗病基因,在不同遺傳背景下均可大幅減輕赤霉病病害[9]。Fhb1最早在我國地方小麥品種蘇麥3號及其衍生品系寧7840中被發掘,已被國內外廣泛用于小麥抗赤霉病育種[10-12]。蘇麥3號是全球普遍公認的高抗赤霉病品種[12],已發現其攜帶有多個抗赤霉病基因/QTLs,其3BS染色體臂上的基因Fhb1既抗赤霉病菌擴展又抗赤霉毒素DON的積累[13],其6BS染色體臂上的基因Fhb2是一個抗擴展的主效基因[14],其5AS染色體臂上的基因Fhb5是一個抗侵染的主效位點[15],其2DL染色體臂上的QFhs.crc-2DL也具有一定的抗病貢獻率[16]。蘇麥3號赤霉病抗性強、穩定且持久,但其春性較強,易遭凍害,植株較高且莖稈纖細易倒伏,重感條銹病、葉銹病和白粉病等病害,產量水平較低,籽粒小且呈粉質,品質較差,對黃淮南部冬麥區的生態環境適應性較差,作為育種親本材料在黃淮南部冬麥區很難被直接利用[17]。當前,我國黃淮南部冬麥區的小麥生產上急需兼抗赤霉病和條銹病的新品種,育種上迫切需要兼抗赤霉病和條銹病的優異親本材料。小麥新品系979-5是本課題組從黃淮南部冬麥區大面積推廣種植的品種西農979中系選而成,屬弱冬性,株高較低(75 cm左右),莖稈粗壯,抗倒伏能力強,中抗條銹病和葉銹病,籽粒大長呈角質,為優質強筋高產類型,但中感赤霉病[18]。為創制適宜黃淮南部冬麥區育種親本材料,本課題組于2012年組配了蘇麥3號與979-5的雜交組合,利用分子標記輔助選擇和田間接種鑒定等技術,創制出20個攜帶有Fhb1、Fhb2及Fhb5基因的優異育種親本材料[19]。2016年起,利用這些優異親本材料,分別與本課題組育成的高產優質新品種西農235、西農282和西農226組配雜交組合,從其后代中選育出13個遺傳穩定的抗赤霉病新品系。目前部分新品系正在參加黃淮南部冬麥區的多點適應性試驗。本研究以這些小麥新品系為材料,在鑒定其赤霉病和條銹病抗性及主要農藝性狀的基礎上,分析其抗赤霉病的遺傳基礎,為小麥新品系的研究和利用提供理論依據。
1 材料與方法
1.1 試驗材料
參試的13個抗赤霉病的小麥新品系(表1),是本課題組前期以高抗赤霉病的親本材料k10、k12和k13[19]為母本,分別與優質高產新品種西農235、西農282和西農226雜交,綜合利用SSR分子標記輔助選擇、赤霉病麥粒菌地表接種和單花滴注法接種鑒定、條銹病菌田間接種鑒定、大群體系譜選擇等技術選育而成。對照材料分別為矮抗58(高感赤霉病,高感條銹病)、979-5(中感赤霉病,中抗條銹病)和蘇麥3號(高抗赤霉病,高感條銹病)。
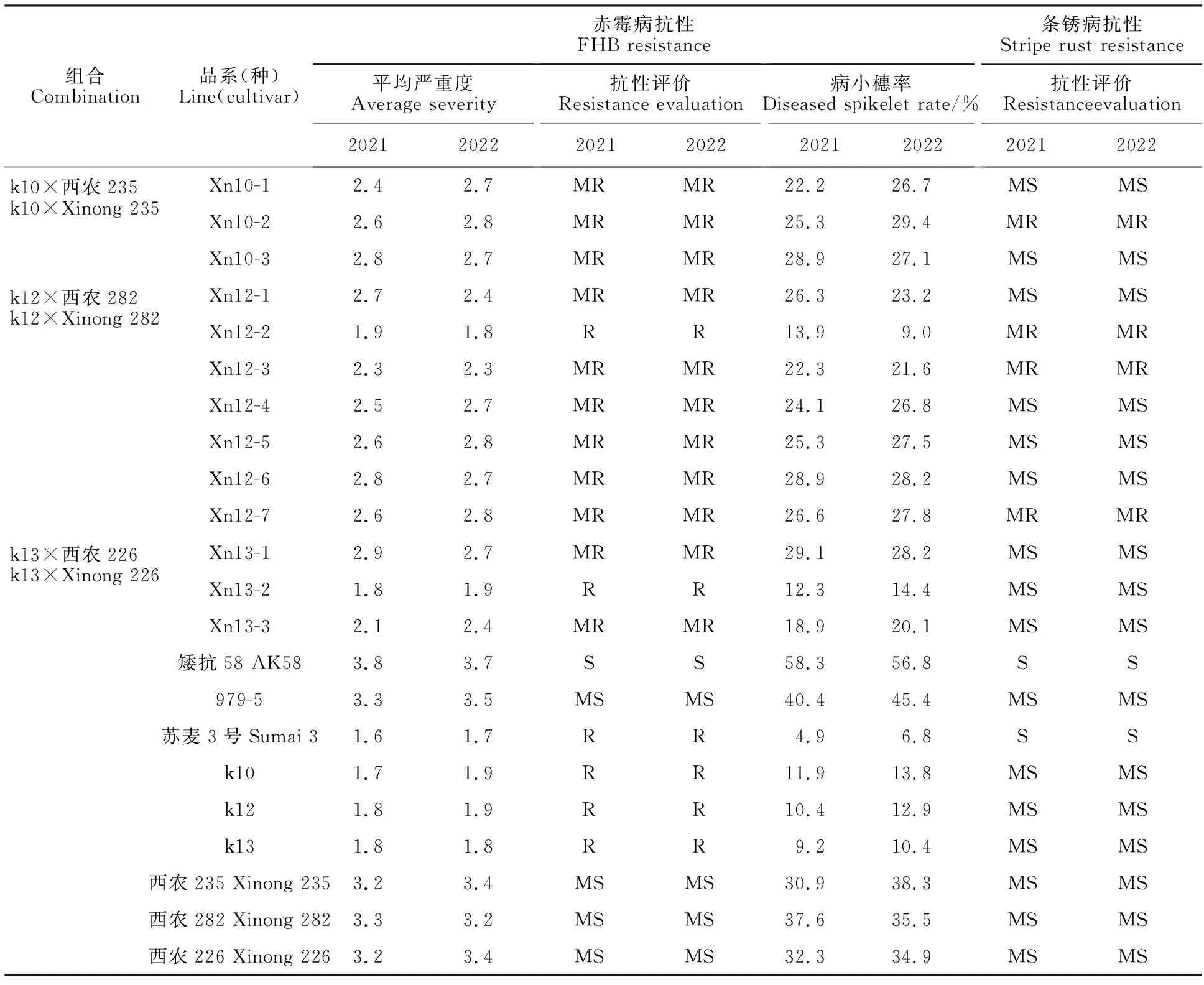
表1 參試材料的赤霉病和條銹病抗性Table 1 Resistance to FHB and stripe rust in tested materials
所有參試材料均進行連續兩年的試驗,分別于2020年10月12日和2021年10月11日播種于西北農林科技大學小麥育種試驗田。每個品系(種)種植3行,株距0.07 m,行距0.27 m,行長2.0 m。在小麥整個生長期間除不進行病害防治外,其余管理措施同當地大田。
1.2 小麥赤霉病田間接種和鑒定
試驗所用的赤霉病4個強致病力菌株(F0980、F1312、F0609和F0301)與國家小麥新品種區域試驗鑒定所用相同,由河南農業大學殷貴鴻研究員惠贈。采用單花滴注法接種:將預先培養的4個菌株孢子液混合并稀釋孢子濃度為1×105·mL-1,在小麥揚花期取10 μL的懸浮液注入小麥穗中上部第4個小穗最外側的小花內,隨即噴濕接種穗,并套袋保濕72 h。每個品系接種20個單穗,兩個重復。
在接種21 d后調查每個接種穗的發病小穗數和總小穗數,計算平均病小穗率和平均嚴重度。病小穗率=發病小穗數/總小穗數×100%。記載標準參照《中華人民共和國農業行業標準NY/T 2954-2016:小麥區域試驗品種抗赤霉病鑒定技術規程》[20],依據平均嚴重度劃分抗病等級:抗病(R)(0<平均嚴重度<2.0),中抗(MR)(2.0≤平均嚴重度<3.0),中感(MS)(3.0≤平均嚴重度<3.5),感病(S)(平均嚴重度≥3.5)。
1.3 小麥條銹病田間接種和鑒定
分別于2020年3月20日和2021年3月15日接種條銹病菌,所用3個條銹病強致病力菌株CYR32、CYR33和CYR34,與國家小麥新品種區域試驗鑒定所用相同,由甘肅省農科院提供。于田間空氣濕度較大的傍晚進行接種,將條銹菌混合小種與淀粉以1∶1 000比例混勻,采用小麥葉心抖粉法,于每個材料的第一行接種20株。當條銹病進入盛發期時,即感病對照品種的病害最大嚴重度達到90%~100%,對發病情況進行田間調查,參照《小麥抗條銹病評價技術規范》[21]分級為:抗病(R,葉片有壞死斑,有零星的孢子),中抗(MR,葉片褪綠,有中等大小的孢子堆),中感(MS,葉片大片褪綠,有大片孢子堆),感病(S,葉片不褪綠,且出現大片孢子堆)。
1.4 農藝性狀調查
在小麥生長發育的關鍵時期,按農藝性狀的記載標準,分別調查幼苗的耐寒性、開花期、株高、穗長、穗粒數、籽粒大小(包括千粒重、粒長、粒寬)等主要農藝性狀。除抗寒性和開花期按群體性狀記載外,其余性狀每個材料調查10個單株,取其平均值。
1.5 抗性基因檢測
采用CTAB法[22]提取苗期小麥鮮葉基因組DNA。
選用已報道的與蘇麥3號抗赤霉病基因Fhb1、Fhb2、Fhb5和QFhs.crc-2DL分別緊密連鎖的分子標記進行檢測,引物及擴增程序同相應的參考文獻(表2)。引物由陜西中科羽瞳生物科技有限公司合成。PCR反應體系為20 μL:10 μL 2×Taq Master Mix for PAGE(諾唯贊,南京),上、下游引物各1 μL(10 μmol·L-1),1 μL模板(50 ng·μL-1),7 μL ddH2O。除標記TaHRC-GSM的PCR擴增產物用2.0 %瓊脂糖凝膠外,其余標記的PCR擴增產物均用8 %非變性聚丙烯酰胺凝膠電泳分析。

表2 選用的抗赤霉病基因/QTL連鎖的分子標記Table 2 Selected molecular markers linked to FHB resistance genes/QTL
1.6 數據處理與分析
所有數據為2年平均值,使用SPSS 20.0對數據進行分析。
2 結果與分析
2.1 參試品系的赤霉病抗性分析
由表1可知,13個參試新品系的赤霉病抗性水平均高于對照中感品種西農979-5,其中2個品系(Xn12-2和Xn13-2)的表現為抗,平均病小穗率為11.5%、13.4%;平均嚴重度均為1.9,抗性水平同蘇麥3號;其余11個品系的赤霉病抗性表現均為中抗,平均病小穗率均小于30%。
2.2 參試品系的條銹病抗性分析
13個參試品系中4個品系表現為中抗條銹病,其余9個品系表現為中感條銹病。其中,Xn12-2抗赤霉病,中抗條銹病;Xn10-2、Xn12-3和Xn12-7中抗赤霉病,中抗條銹病(表1)。這些兼抗赤霉病和條銹病的新品系可作為黃淮麥區優異育種親本材料。
2.3 參試品系的主要農藝性狀分析
從表3可以看出,參試品系均具有耐寒、矮稈、長穗、高粒重的特點。其中,株高為67~82 cm;除品系Xn10-3開花期特別早外(與蘇麥3號相同),其余參試品系的開花期基本與西農979相同。在籽粒性狀中,參試品系的千粒重為39.2~50.2 g,顯著高于蘇麥3號;粒寬3.40~3.64 mm,粒長在6.44~7.53 mm之間,品系間差異較為名顯(圖1)。以上結果表明,參試部分品系不僅具有良好的赤霉病和條銹病抗性,其主要農藝性狀能滿足育種目標要求。

表3 新品系的主要農藝性狀Table 3 Main agronomic traits of the new lines

a和b圖分別為部分材料10個粒籽的長度和寬度。a and b show the length and width of 10 grains of partial lines, respectively.圖1 部分材料的籽粒大小Fig.1 Grain size of the partial lines
2.4 參試品系Fhb1基因分析
利用已報道的與抗赤霉病基因Fhb1緊密關聯的5個分子標記TaHRC-GSM、Xgwm493、Xgwm533、Xbarc133和Xbarc147對參試品系的親本進行了多態性分析發現,親本材料k10、k12和k13與親本材料西農235、西農282和西農226在Xgwm493位點擴增片段大小一致,表現為單態,其余4個標記表現為多態。利用4個具有多態性的標記對參試品系進行分析發現,在TaHRC-GSM標記位點,11個品系檢測到蘇麥3號的特異條帶,Xn10-3和Xn12-1兩個品系中未檢測到蘇麥3號的特異條帶(表4,圖2a);在Xbarc147標記位點,10個品系中檢測到蘇麥3號的特異條帶;在Xbarc133標記位點,3個品系中檢測到蘇麥3號的特異條帶;在Xgwm533標記位點,6個品系中檢測到蘇麥3號的特異條帶。這表明,這些抗赤霉病新品系中,84.6%具有蘇麥3號Fhb1基因的TaHRC-GSM標記,76.9%具有蘇麥3號Xbarc147標記位點,9個品系同時具有蘇麥3號TaHRC-GSM和Xbarc147標記位點。

a和b圖分別為分子標記TaHRC-GSM和Xbarc147的檢測結果;箭頭指示蘇麥3號的特征條帶。a and b show the detection results of molecular marker TaHRC-GSM and Xbarc147, respectively. Arrows indicate the specific bands for Sumai 3.圖2 部分參試材料3BS染色體臂上的分子標記檢測結果Fig.2 Molecular marker detection on 3BS chromosome of some test materials

表4 參試材料的抗赤霉病基因/QTL連鎖分子標記檢測結果Table 4 Detection results of gene/QTL linked molecular markers for FHB resistance in the tested materials
2.5 參試品系Fhb2基因位點的標記分析
利用與抗赤霉病基因Fhb2緊密連鎖的3個分子標記分析發現,親本材料k10、k12和k13與西農235、西農282和西農226在Xgwm133和Xgwm644位點擴增片段大小一致,表現為單態,在標記Xbarc101位點表現為多態。在Xbarc101位點,2個品系(Xn10-1和Xn13-1)檢測到蘇麥3號的特異條帶(表4,圖3a),其余品系中未檢測到蘇麥3號的特征帶譜。

a、b和c圖分別為分子標記Xbarc101、Xgwm154和Xgwm539的檢測結果。箭頭指示蘇麥3號的特征條帶。a, b and c show the detection results of molecular marker Xbarc101, Xgwm154 and Xgwm539, respectively. Arrows indicate the specific bands for Sumai 3.圖3 部分參試材料分子標記檢測結果Fig.3 Molecular marker detection results of some test materials
2.6 參試品系Fhb5基因位點的標記分析
利用與抗赤霉病基因Fhb5緊密連鎖的4個分子標記分析發現,親本材料k10、k12和k13與西農235、西農282和西農226在Xgwm293和Xbarc186位點擴增片段大小一致,表現為單態,在Xgwm304和Xgwm154位點表現為多態。進一步分析發現,在Xgwm304標記位點,Xn12-5、Xn12-6兩個品系中檢測到蘇麥3號的特異條帶;在Xgwm154標記位點,5個品系中檢測到蘇麥3號的特異條帶(表4,圖3b)。
2.7 參試品系QFhs.crc-2DL基因位點的標記分析
利用與抗赤霉病基因QFhs.crc-2DL緊密關聯的2個分子標記分析發現,親本材料k10、k12和k13與西農235、西農282和西農226在Xwmc41位點擴增片段大小一致,表現為單態,在Xgwm539位點表現為多態。進一步分析發現,在Xgwm539位點,11個品系中檢測到蘇麥3號的特異條帶(表4,圖3c)。
3 討論
缺乏適于在我國黃淮南部麥區大面積推廣的抗赤霉病品種,是小麥赤霉病在黃淮南部麥區發病逐年加重的重要原因[30],培育抗赤霉病品種已成為該區小麥育種的主要目標之一[31-32]。蘇麥3號是世界公認的抗赤霉病品種,具有多個抗赤霉病基因/QTLs,其中Fhb1、Fhb2、Fhb5和QFhs.crc-2DL等受到廣泛應用[2]。本課題組以蘇麥3號為抗源,采用分子標記輔助選擇和病原菌接種鑒定的方法,創制出適應黃淮南部麥區生態條件的、攜帶有Fhb1抗病基因位點的抗赤霉病育種親本材料[19],2016年以這些親本材料為基礎,分別與我國黃淮南部冬麥區的高產優質廣適新品種西農226、西農282和西農235等組配雜交組合,采用系譜選擇法,自F2~F6代持續進行赤霉病抗性基因分子標記輔助選擇、病原菌田間土表接種和單花滴注法鑒定,育成赤霉病抗性優異且主要農藝性狀優良的13個新品系。
綜合赤霉病和條銹病抗性鑒定、分子標記檢測和主要農藝性狀分析,參試的13個新品系不僅具有較好的農藝性狀、優異的條銹病抗性,且赤霉病抗性水平達抗或中抗。其中,11個品系含有蘇麥3號的TaHRC-GSM位點,可能攜帶抗性基因Fhb1,其中2個品系(Xn12-2和Xn13-2)赤霉病抗性同蘇麥3號,表現為抗。這也進一步證明,TaHRC-GSM是抗性基因Fhb1的可靠有效選擇標記[7]。這11個品系同時還具有蘇麥3號其它位點的優異等位變異,如Xn12-2具有QFhs.crc-2DL的優異等位變異,Xn13-2具有Fhb5和QFhs.crc-2DL的優異等位變異,Xn13-1具有Fhb2和QFhs.crc-2DL的優異等位變異。但中抗赤霉病的品系Xn10-3和Xn12-1卻沒有蘇麥3號的TaHRC-GSM標記位點,僅攜帶蘇麥3號的Xgwm533和Xgwm539位點的標記,其抗性來源有待進一步研究。本研究選用的參試新品系,其赤霉病抗性水平表現好且主要農藝性狀較好的根本原因,可能與本課題組近10余年來一直堅持“自主創新親本材料、強化早代大群體多途徑赤霉病抗性鑒選技術”的應用密不可分:(1)創制適宜黃淮冬麥區的優異抗赤霉病k系列親本材料[19],并選用赤霉病田間抗性較好的育成品種(西農235、西農282、西農226等)組配雜交組合,為優良抗赤霉病新品系(種)選育奠定良好的遺傳基礎;(2)F2大群體(每個組合2 000~3 000株)利用麥粒菌及玉米秸稈結合進行地表接種,苗期條銹病菌接種鑒定,開花期選優株單花滴注法接種,綜合赤霉病和條銹病抗性進行優良單株選擇;(3)中選單株的F3-5株系群,根據收獲的種子量盡可能保證較大的群體,繼續上述的抗性鑒選,中選單株繼續赤霉病抗性基因/QTLs的分子標記分析,了解其抗性遺傳背景。持續兩年的田間種植觀察發現,個別參試品系還中抗葉銹病(田間自然發病下),但葉尖枯現象較重,可能影響籽粒產量,因此還需要多年多點產量比較試驗以進一步改良。
