
油莎豆種質資源低溫脅迫生理響應與耐寒性評價
2023-10-15 06:00:10鐘鵬苗麗麗王建麗劉杰王曉龍
中國農業科技導報 2023年9期
鐘鵬, 苗麗麗, 王建麗, 劉杰, 王曉龍
(1.黑龍江省農業科學院畜牧獸醫分院,黑龍江 齊齊哈爾 161005; 2.黑龍江省黑土保護利用研究院,哈爾濱 150086; 3.黑龍江省農業科學院草業研究所,哈爾濱 150086)
中美貿易摩擦不斷升級使我國食用油和飼料安全問題日益凸顯,替代進口大豆、降低食用油和飼料對外依存度成為公眾關注的熱點和焦點。解決油料需求增長與耕地面積有限之間的矛盾,必需尋找單產和品質均高于大豆的食用油新油源作物開發利用。油莎豆被證明是能夠部分替代進口大豆、最為現實可行的糧油飼兼用型作物[1]。油莎豆屬莎草科一年生植物,耐旱、耐貧瘠、防風固沙能力強,比較適宜沙質土壤和沙化邊際土地生長,不與糧爭地[2-4]。在我國東北農牧交錯區適時發展油莎豆產業,將是補充優質食用油供給、保護農牧交錯區生態環境、補齊農業發展短板的有效途徑。
低溫是植物生長發育的限制因子之一,低溫冷害會抑制植物生長發育、破壞細胞結構并影響正常生理代謝[5]。玉米、水稻、花生及果菜等耐低溫脅迫研究較多,已經在耐低溫性品種鑒定、低溫脅迫生理響應及外源調節劑作用機理等方面取得一定進展[6-10]。盡管油莎豆引進我國已經70余年,但對其研究相對滯后。目前,國內外關于油莎豆的研究主要集中在栽培技術、營養成分分析、加工工藝、功能性食品開發等方面[11-14],國內關于油莎豆萌發期及苗期耐低溫全面系統的評價尚未見報道。黑龍江省十年九春寒,且近幾年來倒春寒現象頻發,溫度成為限制該地區油莎豆種植的主要因素。篩選和評價油莎豆耐低溫種質資源,挖掘油莎豆耐低溫基因對東北農牧交錯區水土流失治理、黑土地保護利用及鹽堿地改良具有重要意義。本研究在低溫脅迫下對不同生態區油莎豆種質資源耐低溫性進行鑒定和評價,以期明確油莎豆萌發期和苗期耐低溫的主要鑒定指標,并篩選出耐低溫較強的油莎豆種質資源,為后續培育耐低溫品種、克隆耐低溫基因及功能驗證提供材料。
1 材料與方法
1.1 試驗材料
試驗材料為2010 年以來引進國內外不同生態區的18 份油莎豆種質資源材料,由黑龍江省農業科學院農村能源研究所能源植物資源利用研究室提供。由于國內油莎豆只有1 個登記品種中油莎1 號(ZYS01),其余均為種質材料,均用代號表示。18 份油莎豆種質材料如下:HLJ105、RY126、BY156、HLJ106、RY102、BY141、RY114、NM103、XJ101、JL154、LN181、SX100、HB122、HN162、ZYS01、XY109、GD187和QY186。
1.2 試驗設計
大田試驗在黑龍江省大慶市杜爾伯特蒙古族自治縣新店林場試驗基地(46°32′ N,124°30′ E)進行,該區屬中溫帶大陸季風氣候。2018—2020年均于5 月1 日播種,每穴1 粒,隨機區組設計,3次重復,小區4.5 m×10 m,行株距45 cm×20 cm。倒春寒規定及等級標準參照文獻[15-16]。
室內試驗在黑龍江省農業科學院農村能源研究所進行。油莎豆種子萌發試驗使用人工氣候箱,光照時間為6:00—18:00,濕度80%。將種子置于直徑為20 cm 的培養皿中,45 ℃溫水浸泡吸脹36 h,根據大田試驗倒春寒日最低溫度范圍,以正常溫度(25 ℃)為對照,將溫度設置3、5、7、9 ℃共4 個溫度處理,分別培養3 d,25 ℃復溫后繼續培養,記錄發芽時間、發芽數、芽長等數據。每個試驗組每個油莎豆種質材料50粒種子,重復3次。
將油莎豆種子45 ℃溫水浸泡吸脹36 h 后播種于盛珍珠巖的營養缽中,澆灌1/2 經典霍格蘭(Hoagland)營養液培養萌發后,當幼苗第3 片葉完全展開后,進行低溫處理。將幼苗置人工氣候箱中,分別于3 、5 、7 、9 ℃ 4個溫度處理2 h,以正常溫度(25 ℃)為對照。每個試驗組的每個種質材料8株,3次重復。
1.3 測定目標及方法
1.3.1 大田試驗出苗時間和出苗率統計 2018—2020 年播種后10 d,在18 個油莎豆種質材料小區分別隨機挑選3條壟每天調查出苗情況,記錄出苗時間(emergence date, ED)、出苗數(emergence plant,EP),直至6月3日。統計播種穴數和出苗穴數,根據以下公式計算出苗率(emergence rate, ER)。
1.3.2 室內發芽試驗 以正常溫度為對照,統計18 個油莎豆種質材料在不同處理后種子發芽數、發芽時間、芽長等數據。計算發芽率(germination rate, GR)、發芽指數(germination index,GI)和種子活力指數(seed vigor index,VI)等參數[17-18]。
式中,M指發芽種子數,N指供試樣品的種子數;Gt指不同發芽天數的發芽率,Dt指對應的發芽天數;S指長出部分鮮重。
1.3.3 生化特性指標測定 采用張憲政[19]的丙酮乙醇混合液法測定計算葉綠素(a+b)含量[chlorophyll content,Chl(a+b)]。相對膜透性(relative membrane permeability, RMP)測定參考李錦樹等[20]的方法。丙二醛(malondialdehyde, MDA)含量測定參照Guo 等[21]的方法并進行改進。可溶性糖(soluble sugar, SS)含量和游離脯氨酸(proline, Pro)含量測定參照常博文等[7]的方法。
1.3.4 抗氧化酶活性測定 超氧化物歧化酶(superoxide dismutase, SOD)活性的測定參照Giannopolitis 等[22]的方法。過氧化物酶(peroxidase,POD)活性的測定參照Klapheck等[23]的方法。
1.4 統計分析
使用SPSS 22.0 軟件進行統計分析,用Duncan’s檢驗(P=0.05)進行顯著性分析。根據隸屬函數法,綜合各指標數據計算18 份種質材料隸屬函數值進行抗寒性評價[24-25]。
2 結果與分析
2.1 油莎豆大田出苗時間及出苗率統計
2018—2020 年4 月28 日至6 月3 日期間試驗場地連續3年均發生倒春寒現象。2018—2020年5 月11 日開始調查出苗情況,由圖1 可知,HLJ105、RY126、BY156、HLJ106、RY102 和BY141出苗相對較早,每年均在5 月11 日開始出苗,于5月20 日出苗結束,出苗率均達到85.0%以上;QY186、GD187、XY109、ZYS01、HB122 和HN162出苗相對較晚,5 月14 日開始出苗,5 月22 日出苗結束,出苗率均低于65.0%。
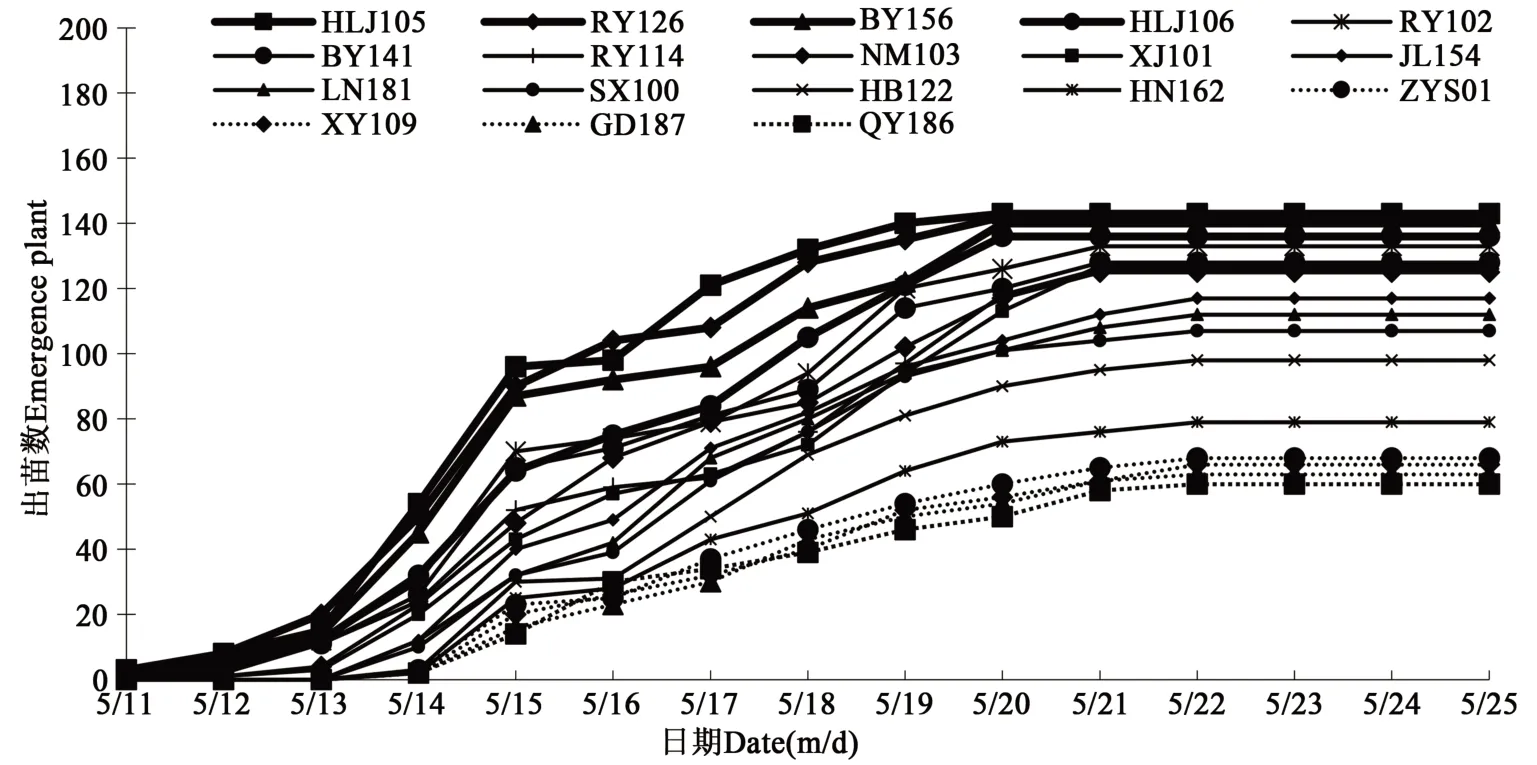
圖1 2018—2020年18份種質材料出苗情況Fig. 1 Emergence of 18 germplasm materials in 2018-2020
2018—2020 年均在6 月3 日統計出苗率,由表1 可知,3 年出苗率較高的種質材料為HLJ105、RY126、BY156、HLJ106、RY102 和BY141,平均出苗率為95.21%、94.63%、93.32%、90.50%、88.96%和85.93%;而出苗率較低的種質材料為QY186、GD187、XY109、ZYS01、HN162 和HB122,平均出苗率為40.13%、41.88%、43.84%、45.34%、52.98%和65.56%。從3年氣溫變化、出苗情況、出苗率綜合比較,HLJ105、RY126、BY156 和HLJ106 耐低溫性較強,QY186、GD187、XY109 和ZYS01 耐低溫性較弱。
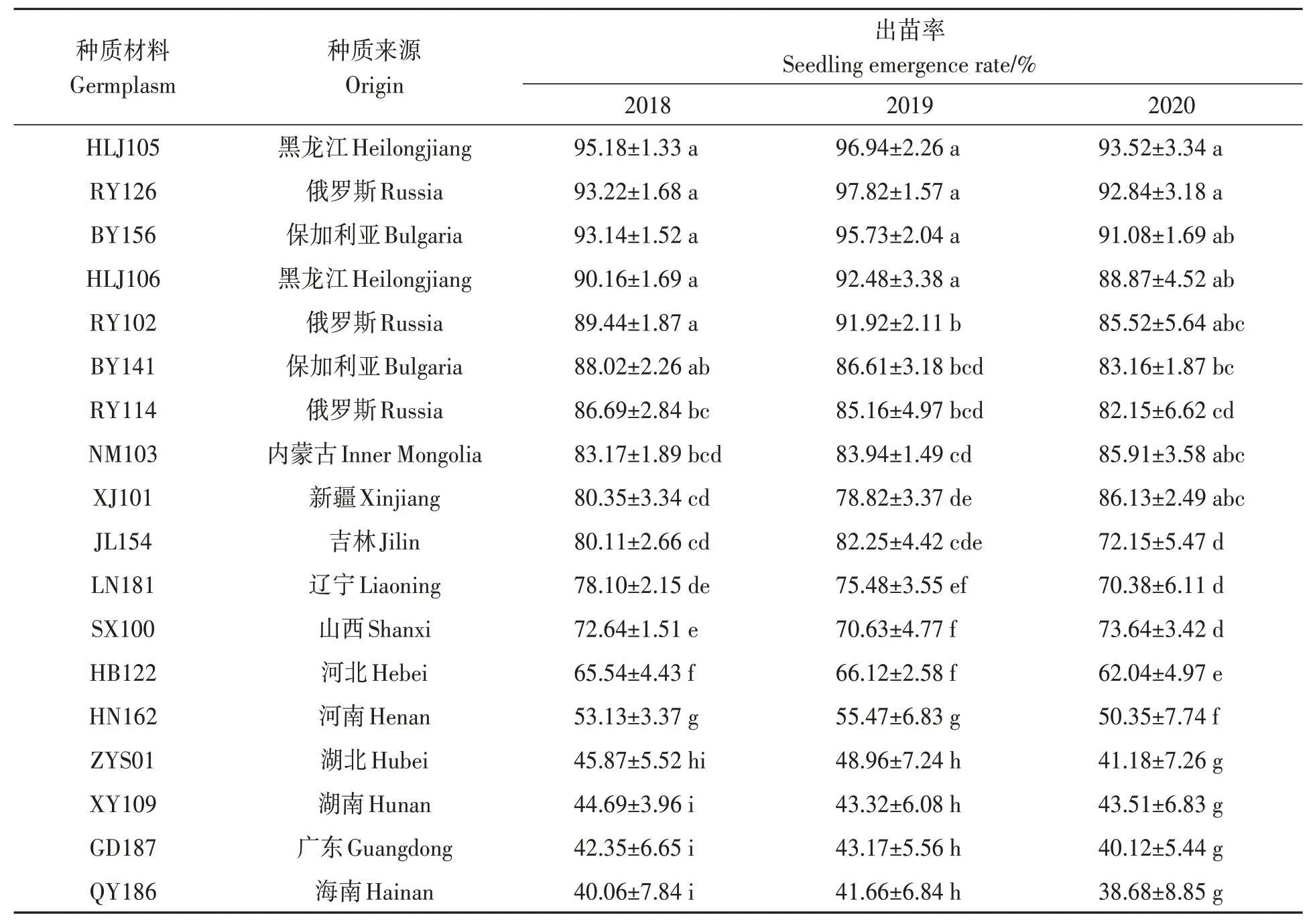
表1 油莎豆種質材料出苗率Table 1 Seedling emergence rates of all Cyperus esculentus germplasms
2.2 室內低溫脅迫對油莎豆種子發芽指數和活力指數的影響
由表2 可知,隨著處理溫度的降低,不同油莎豆種質材料發芽指數均逐漸下降。3 ℃處理時,HLJ105、RY126、BY156 和HLJ106 種子發芽指數相對較高,與其他14 份材料差異均達顯著水平(P<0.05);QY186、GD187、XY109和ZYS01種子發芽指數相對較低,除與HN162差異不顯著外,與其余13 份材料差異均達顯著水平(P<0.05)。5 ℃處理時,HLJ105、RY126、BY156 和HLJ106 種子發芽指數顯著高于其他14 份材料;QY186、GD187、XY109 和ZYS01 種子發芽指數相對較低,除與HN162、SX100 和JL154 差異不顯著外,顯著低于其余11 份材料。7 ℃處理時,HLJ105、RY126、BY156、HLJ106、RY102、BY141、NM103、JL154 和HN162共9份材料之間差異不顯著,但均顯著高于除LN181外的其他8份材料。9 ℃處理時,與對照組發芽指數基本一致,可達到對照組的97.6%。
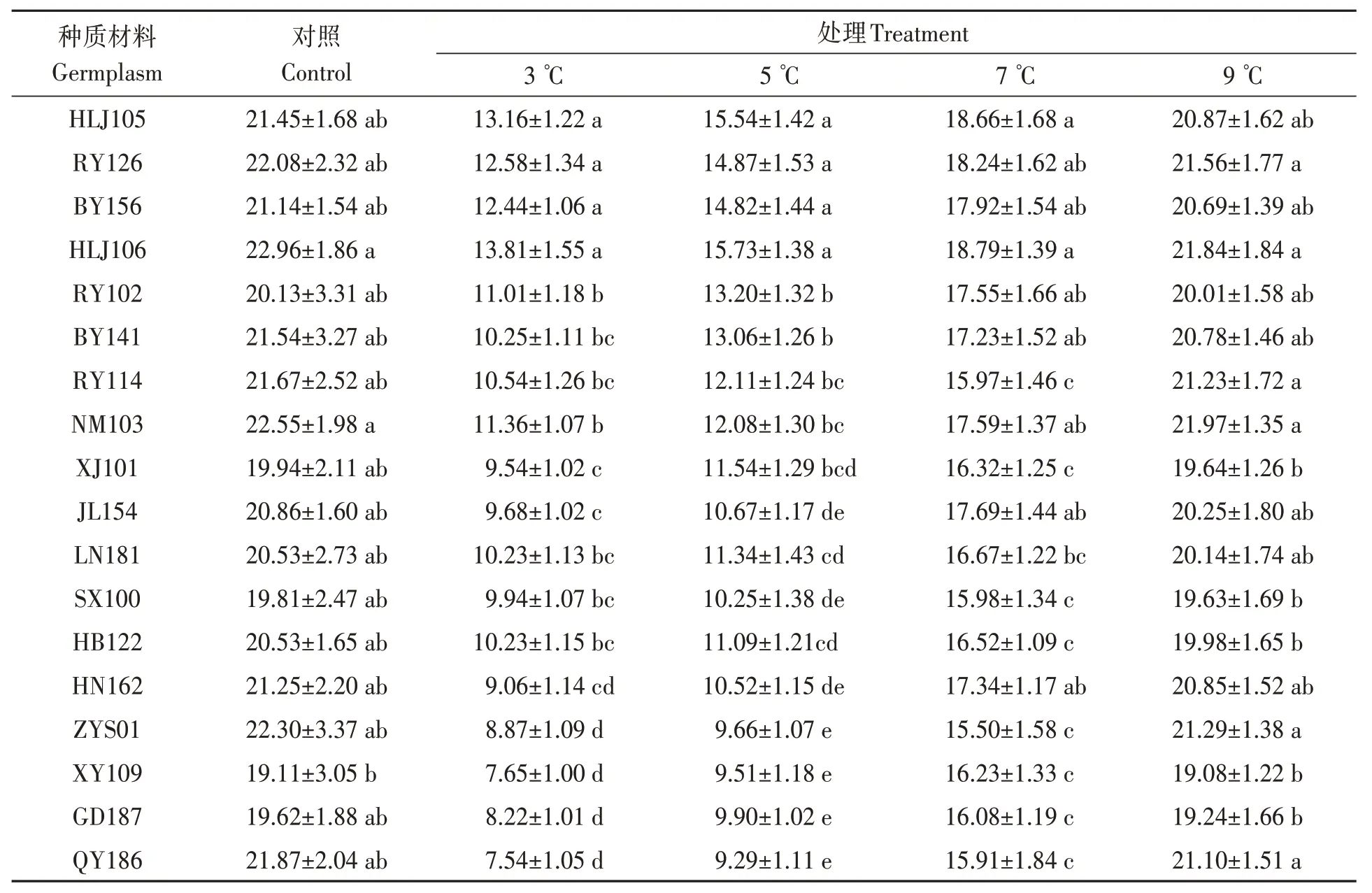
表2 低溫脅迫對油莎豆種子發芽指數的影響Table 2 Effects of low temperature on germination index of Cyperus esculentus germplasms
由表3可知,隨著處理溫度的降低,除NM103外其他油莎豆種子活力指數逐漸下降。3 ℃處理時,HLJ105、RY126、BY156 和HLJ106 種子活力指數相對較高,與其他14 份材料差異均達顯著水平(P<0.05);QY186、GD187、XY109 和ZYS01 種子活力指數顯著低于其他14 份材料(P<0.05)。5 ℃處理時,HLJ105、RY126、BY156、HLJ106、RY102和BY141 種子活力指數相對較高,與其他12 份材料差異均達顯著水平;QY186、GD187、XY109 和ZYS01 種子活力指數相對較低,與其他14 份材料差異均達顯著水平。7 ℃處理時,HLJ105、RY126、BY156、HLJ106、BY141、ZYS01、RY102 和LN181 這8 份材料之間差異不顯著,均與GD187之間差異顯著。9 ℃處理時,處理組較對照組相比活力指數下降幅度不大,可達到對照組的96.8%。
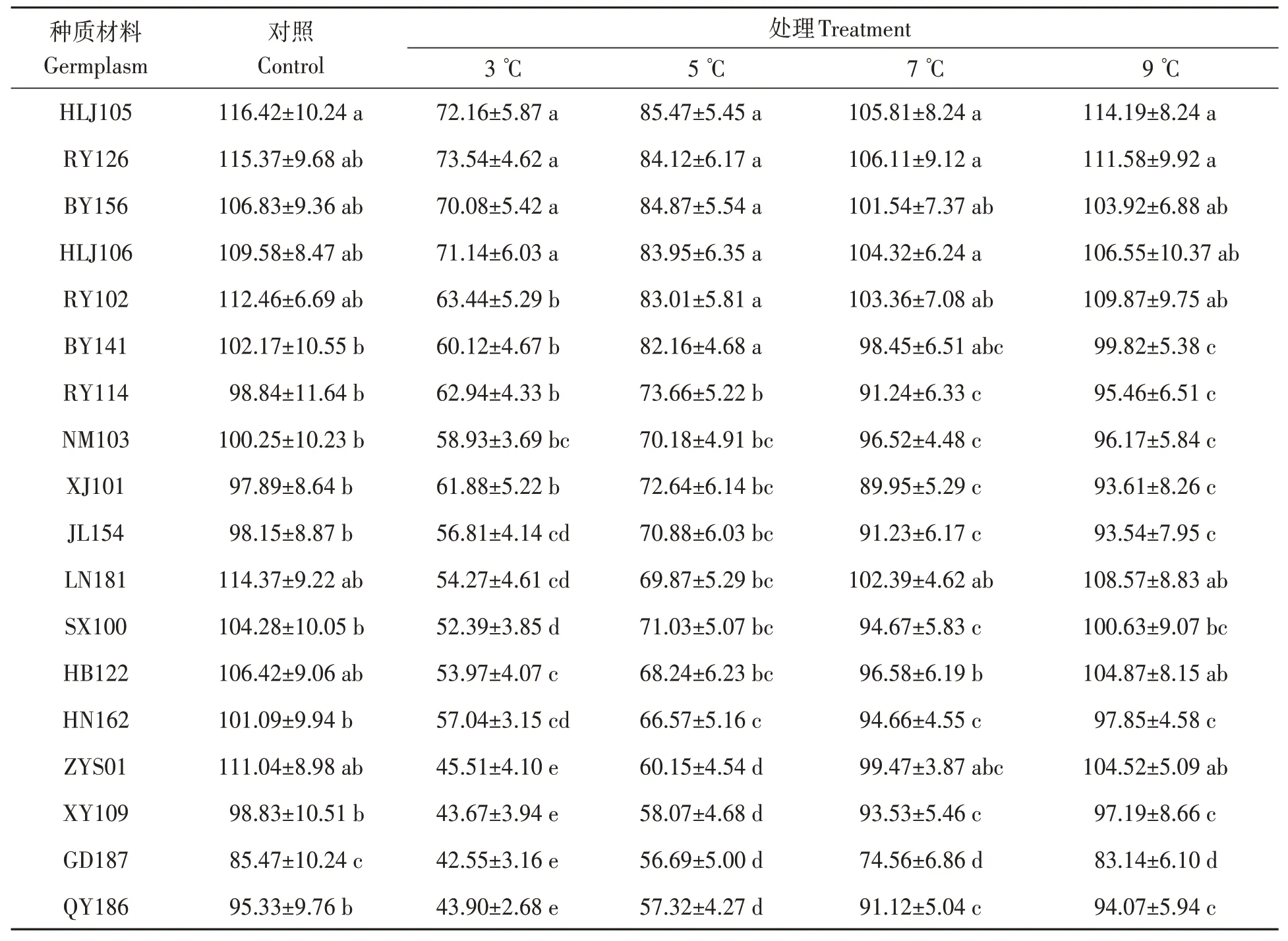
表3 低溫脅迫對油莎豆種子活力指數的影響Table 3 Effects of low temperature on seed vigor of Cyperus esculentus germplasms
2.3 低溫脅迫對油莎豆幼苗葉綠素含量的影響
隨著溫度的降低,油莎豆幼苗葉片葉綠素(a+b)含量逐漸降低(圖2)。3 ℃處理時,HLJ105、RY126、BY156 和HLJ106 葉綠素(a+b)含量較高;QY186、GD187、XY109 和ZYS01 葉綠素(a+b)含量較低。5 ℃處理時,HLJ105、RY126、BY156 和HLJ106葉綠素(a+b)含量相對較高。7 和9 ℃處理時所有材料之間葉綠素(a+b)含量差別不明顯。
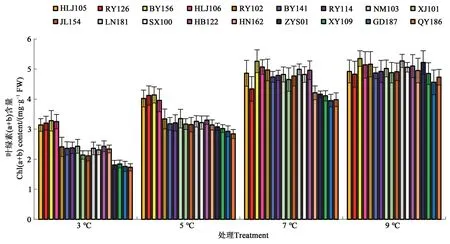
圖2 低溫脅迫對油莎豆幼苗葉片葉綠素含量的影響Fig. 2 Effects of low-temperature on the Chl(a+b) contents of Cyperus esculentus germplasms leaves
2.4 低溫脅迫對油莎豆幼苗葉片質膜損傷程度的影響
隨著處理溫度的降低,油莎豆葉片相對膜透性逐漸增大(表4)。3 和5 ℃處理時,HLJ105、RY126、BY156 和HLJ106 與對照相比相對膜透性均增加,但顯著低于其他14 份種質材料。3 和5 ℃處理,QY186、GD187、XY109 和ZYS01 與對照相比相對膜透性均增加,但顯著高于其他14 份材料。7 ℃處理下,QY186、GD187、XY109、HN162、JL154 和ZYS01 相對膜透性較大。9 ℃處理時,QY186 和GD187 相對膜透性與XJ101、BY141、RY102、BY156 和HLJ105 差異顯著,與其他材料之間差異不明顯。
3和5 ℃處理,丙二醛含量變化與相對膜透性基本相似,HLJ105、RY126、BY156 和HLJ106 丙二醛含量較對照均增加,但丙二醛含量相對較少,顯著低于其他14 份種質材料;QY186、GD187、XY109 和ZYS01 丙二醛含量較對照均增加,但丙二醛含量相對較大,顯著高于其他14 份材料。7 ℃處理時,HLJ105、RY126、BY156、RY114、SX100 和HLJ106 丙二醛含量相對較低,與除HN162 外的其他11 份材料差異顯著。9 ℃處理時,HLJ105、RY126、BY156、RY102、RY114、LN181、HLJ106 的丙二醛含量與QY186、GD187、XY109、ZYS01 之間差異顯著,與其他材料之間差異不顯著。
2.5 低溫脅迫對油莎豆幼苗葉片可溶性物質含量的影響
除RY114 外,油莎豆葉片游離脯氨酸含量隨著溫度的降低急劇升高(表5)。當處理溫度為3 ℃時,HLJ105、RY126、BY156 和HLJ106 游離脯氨酸含量顯著高于其他14 份材料;QY186、GD187、XY109和ZYS01顯著低于其他14份材料。處理溫度為5 ℃時,HLJ105、RY126、BY156、RY102、HLJ106 和BY141 游離脯氨酸含量顯著高于其他12 份材料。處理溫度為7 ℃時,HLJ105、RY126、BY156、RY102、BY141、RY114和HLJ106游離脯氨酸含量顯著高于其他11 份材料;處理溫度為9 ℃時,HLJ105、BY156 與HB122、QY186、GD187、XY109之間游離脯氨酸含量差異顯著。
除HN162 外,油莎豆葉片可溶性糖含量隨著溫度的降低而增加(表5)。處理溫度為3 ℃時,HLJ105、RY126、BY156 和HLJ106 可溶性糖含量顯著高于其他14 份材料;QY186、GD187、XY109和ZYS01 顯著低于其他14 份材料。在處理溫度5 ℃時,HLJ105、RY126、BY156、HLJ106、RY102、BY141 和RY114 可溶性糖含量顯著高于其他11份材料。在處理溫度7 ℃時,QY186、GD187、XY109、HN162、XJ101 和ZYS01 可溶性糖含量顯著低于其他12份材料。
2.6 低溫脅迫對油莎豆幼苗葉片抗氧化酶活性的影響
由表6 可知,SOD 活性隨著溫度的降低而升高(HN162 除外)。當溫度降至3 ℃時,HLJ105、RY126、BY156 和HLJ106 葉片中SOD 活性顯著高于其他14 份材料;QY186、GD187、XY109 和ZYS01 顯著低于其他14 份材料。當處理溫度為5 ℃時,HLJ105、RY126、BY156、HLJ106 和RY102材料葉片中SOD 活性顯著高于其他13 份材料;QY186、GD187、XY109 和ZYS01 顯著低于其他14份材料。7 和9 ℃時,HLJ105、RY126、BY156、HLJ106、RY102、BY141 和RY114 材料SOD 活性顯著高于其他11 份材料,其余材料之間差異不顯著。
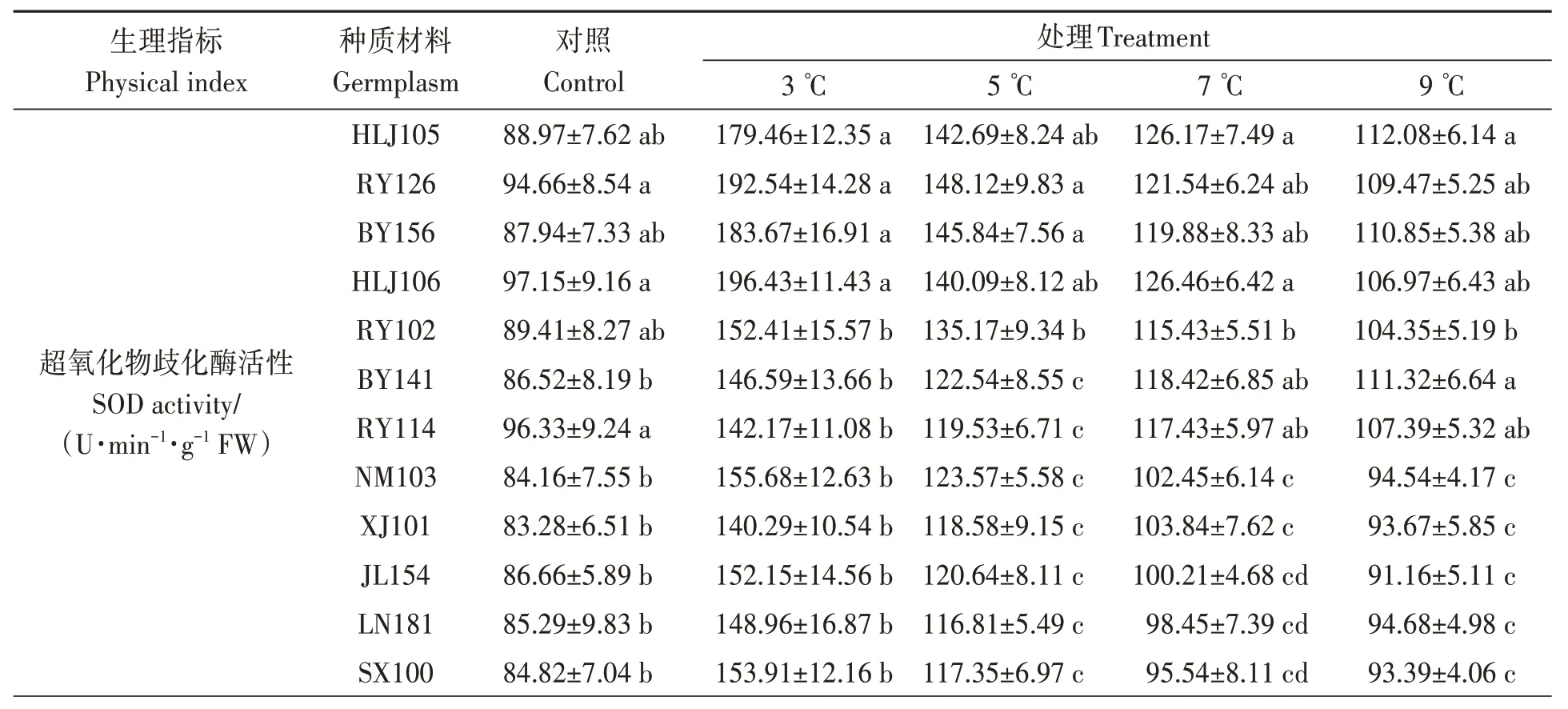
表6 低溫脅迫對油莎豆幼苗葉片抗氧化酶活性的影響Table 6 Effects of low-temperature on anti-oxidase activity of Cyperus esculentus germplasms leaves
POD 活性變化趨勢與SOD 大致相同。當溫度降至3 ℃時,HLJ105、RY126、BY156 和HLJ106葉片中POD 活性顯著高于其他14 份材料;QY186、GD187、XY109 和ZYS01 顯著低于其他14份材料。當溫度為5 ℃時,HLJ105、RY126、BY156、HLJ106、RY102、BY141和RY114材料葉片中POD 活性顯著高于其他11 份材料。當溫度為7和9 ℃時,18份材料之間POD活性差異不顯著。
2.7 18份油莎豆種質材料抗寒性綜合評價
18份油莎豆種質材料抗寒性綜合評價情況見表7。參照劉慧民等[24]、張文娥等[25]的方法,將各材料綜合抗寒能力平均隸屬度分為5 級。抗寒性綜合評價值(cold-resistance integrated assessment value, IAV)≥0.70為強抗寒性,定為Ⅰ級;抗寒性綜合評價值在0. 60~0. 69為抗寒性,定為Ⅱ級;抗寒性綜合評價值在0.40~0. 59為中抗寒性,定為Ⅲ級;抗寒性綜合評價值在0. 30~0. 39為弱抗寒性,定為Ⅳ級;綜合評價值< 0.30 為不抗寒性,定為Ⅴ級。經大田、室內種子發芽和幼苗抗寒性試驗綜合評價HLJ105、RY126、BY156和HLJ106抗寒性綜合評價值分別為0.708 6、0.712 4、0.715 3 和0.705 4,為強抗寒性種質材料;RY102和BY141抗寒性綜合評價值分別為0.616 0 和0.601 5,為抗寒性材料;RY114、NM103、XJ101、JL154、LN181 和SX100 抗寒性綜合評價值在0.40~0. 59,為中抗寒性材料;HB122 和HN162 抗寒性綜合評價值在0. 30~0. 39之間,為弱抗寒性材料;QY186、GD187、XY109 和ZYS01抗寒性綜合評價值均小于0.30,為不抗寒性材料。
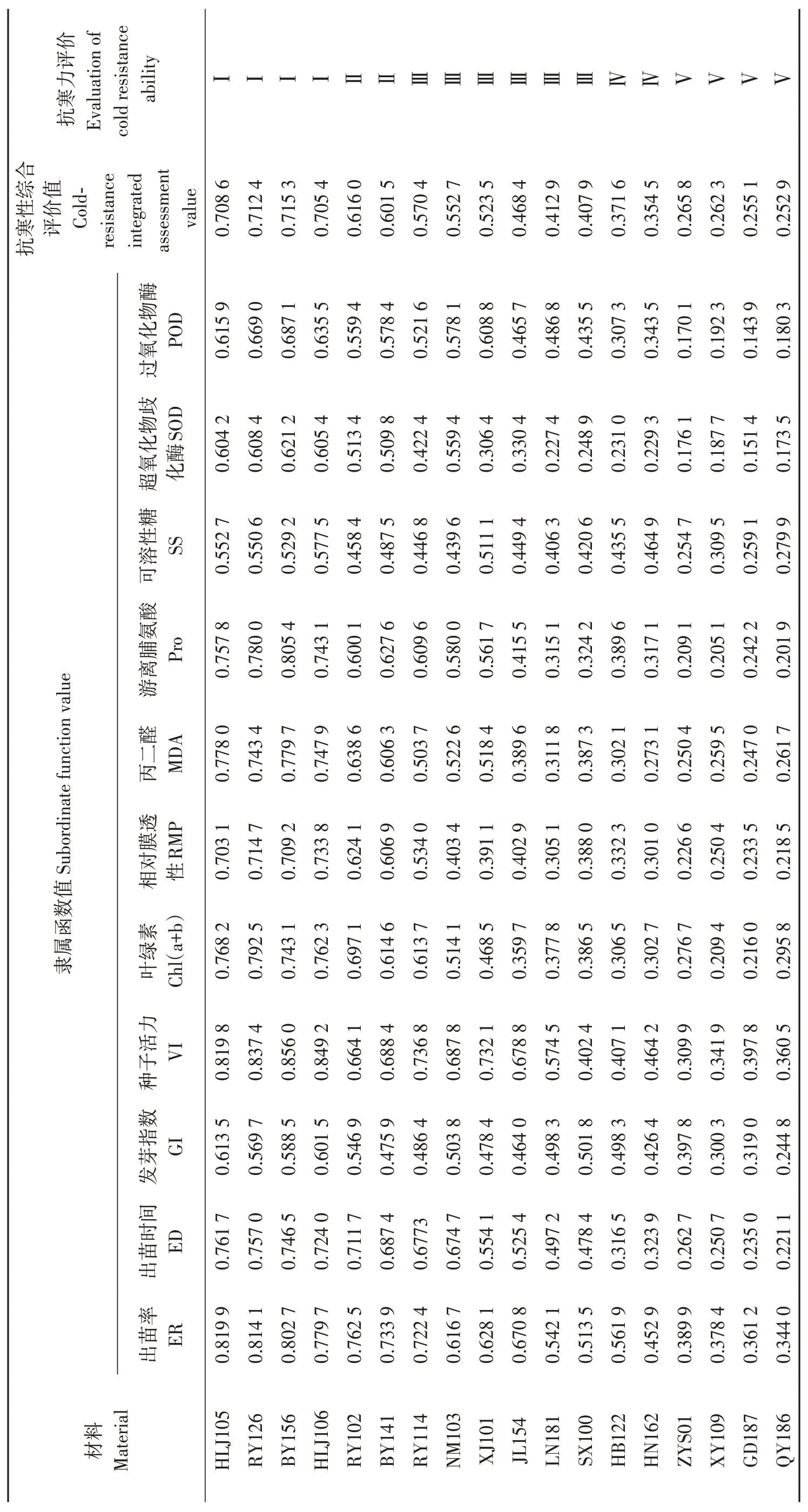
表7 油莎豆各指標隸屬函數值、權重及抗寒性綜合評價值Table 7 Subordinate function values,weight function and cold-resistance integrated evaluation values of all indexes in Cyperus esculentus germplasms
2.8 各測定指標抗寒性相關分析
低溫脅迫下出苗率(ER)、發芽指數(GI)和超氧化物歧化酶(SOD)活性與抗寒性綜合平均值(IAV)相關系數分別為0.978 2、0.953 6和0.970 1,呈顯著正相關(P<0.05),出苗時間(ED)和丙二醛(MDA)含量與抗寒性綜合平均值(IAV)相關系數分別為-0.912 8 和-0.923 3,呈顯著負相關(P<0.05)(表8)。種子活力(VI)、葉綠素[Chl(a+b)]、游離脯氨酸(Pro)、可溶性糖(SS)含量和過氧化物酶(POD)活性與抗寒性綜合評價值(IAV)呈正相關,但相關性均不顯著;相對膜透性(RMP)與抗寒性綜合評價值(IAV)呈負相關,相關性不顯著。出苗率(ER)與發芽指數(GI)、種子活力(VI)呈正相關,相關性達到極顯著水平(P<0.01)。
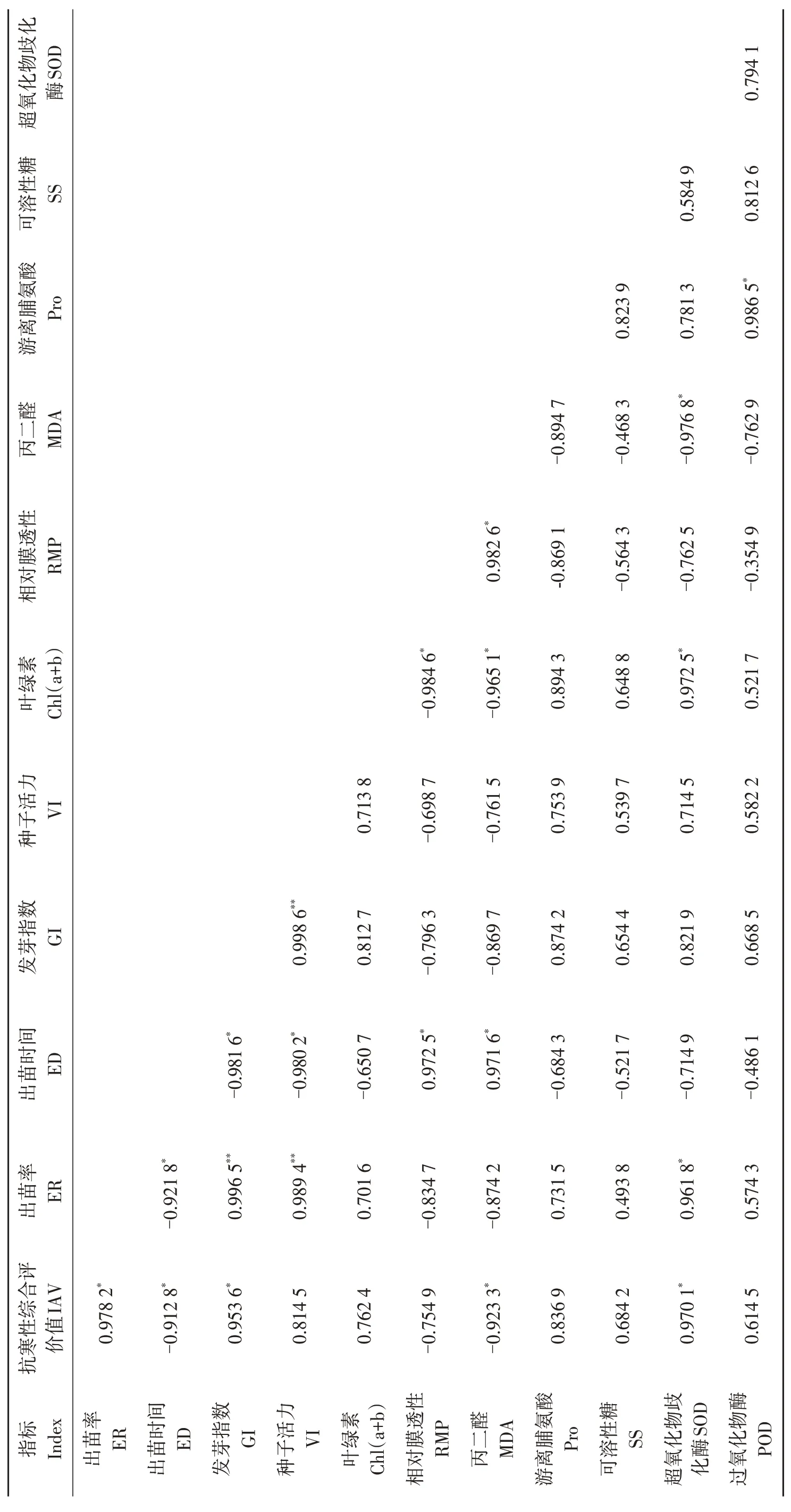
表8 供試油莎豆材料各測定指標及苗期抗寒相關性分析Table 8 Correlation analysis of subordinate function values of cold resistance index for tested varieties in Cyperus esculentus germplasms
3 討論
低溫不僅影響植物生長發育,而且限制其地理分布,當受到低溫脅迫時植物可通過調節自身生理代謝和基因表達來增強低溫耐受性[26]。近年來,倒春寒現象在我國發生頻率逐漸上升,尤其東北地區更為嚴重,輕者補苗,重者毀種,給農業生產帶來不可估量的損失[27]。種子在吸脹萌動和初始發育階段對低溫最為敏感[28]。本團隊前期通過低溫脅迫篩選出耐低溫和不耐低溫的花生品種,明確了耐低溫品種發芽率、發芽指數、種子活力和芽長均高于不耐低溫品種[7,29]。本試驗以油莎豆為研究對象,大田試驗中連續3 年經歷倒春寒,HLJ105、RY126、BY156 和HLJ106 出苗時間較其他材料早3 d,出苗率均大于90.0%,并且與其他材料相比差異顯著,說明其耐低溫能力較強,低溫對其種子萌發出苗影響較小;而QY186、GD187、XY109 和ZYS01 出苗時間較其他材料晚,出苗率均小于50.0%,并且與其他材料相比差異顯著,說明其耐低溫能力較弱,倒春寒對其種子萌發出苗影響較大。室內試驗發芽率、種子活力等指數與大田結果一致。
細胞膜對維持植物細胞內環境穩定性起著重要作用。低溫脅迫時,丙二醛含量激增,導致質膜透性增大,胞內電解質外滲,引起相對電導率升高[30]。本研究顯示,隨著溫度的降低,18份種質材料的質膜透性和丙二醛含量均顯著升高,說明溫度越低油莎豆葉片細胞膜受冷害程度越嚴重;當溫度降至3和5 ℃時,材料HLJ105、RY126、BY156和HLJ106 的相對膜透性和丙二醛含量上升幅度低于其他材料,而QY186、GD187、XY109 和ZYS01 上升幅度高于其他材料,說明耐低溫性強的油莎豆材料細胞膜受低溫傷害較小,抗寒性優于其他材料。質膜透性和丙二醛含量呈顯著正相關,說明油莎豆葉片質膜損傷大部分是由丙二醛含量積累所導致,與之前花生抗寒性評價研究結果不一致[29],有待進一步深入研究。
