亞洲沙塵中鐵、銅對黃海近海表層優勢浮游細菌豐度影響的模擬研究?
2023-10-18 04:04:06丁雅楠祁建華
中國海洋大學學報(自然科學版) 2023年11期
丁雅楠, 那 紅, 祁建華??
(1. 中國海洋大學海洋環境與生態教育部重點實驗室, 山東 青島 266100;2. 青島海洋科學與技術試點國家實驗室 海洋生態與環境科學功能實驗室, 山東 青島 266237)
海洋浮游細菌是海洋生物群落的重要組成部分,它們可以適應高溫、高壓、低壓、低氧等復雜多變的海洋環境,能夠參與碳、氮、磷、硫等生源要素的生物地球化學循環,在海洋食物鏈中起著重要作用,也是海洋微生物食物環的重要環節[1-3]。細菌的豐度和群落結構決定了它們的生態功能,而環境條件的變化,如沙塵沉降等,能夠影響細菌的生物活動、群落結構和生態功能[4-6]。
近海營養鹽及微量金屬的濃度均高于遠洋,與貧營養開闊海洋相比,近海富營養化水域的浮游細菌生長效率更高[7-8],并且近海生物在富營養環境中對必需金屬等元素的需求可能非常高[9],也有些海洋細菌對高濃度重金屬具有耐受性[10-11]。沙塵沉降為近海輸入大量的營養鹽及微量金屬元素,同樣會影響近海浮游細菌的生長和生理作用。其中,鐵對海洋的初級生產和次級生產具有限制作用,同時鐵也是許多代謝過程(有氧呼吸、光合作用、硝化作用和固氮等)必不可少的輔助因子[12]。銅是海洋生物呼吸蛋白質和氧化酶的組成部分[13],對海洋浮游細菌至關重要;但海洋中銅濃度較高時,可能會對浮游植物具有毒性作用[14],進而影響海洋浮游細菌的生長。目前有研究發現,在高營養低葉綠素(HNLC)海區(如佛羅里達群島),撒哈拉沙塵中的鐵可以緩解該海區鐵的限制,從而刺激弧菌生長[15];而在北大西洋百慕大群島附近進行的沙塵添加實驗表明,無機鐵并不是刺激海洋弧菌生長的主要因素,沙塵中的磷以及其他微量金屬元素(Zn、Cu、Mn等)可能是刺激弧菌生長的主要因素[16]。也有在HNLC地區(如亞北極太平洋地區)和印度洋極地地區的研究發現,單獨添加鐵并不能促進該地區細菌的生產和生長能力[17-18]。目前,微量金屬對海洋浮游細菌生長的研究主要集中于鐵對海洋浮游細菌生長的影響,銅對海洋浮游細菌作用的研究較少。關于海洋浮游細菌對沙塵沉降響應的研究多集中于貧營養海域,而富營養近海受沙塵傳輸沉降的影響更為頻繁,因此研究沙塵對近海海洋細菌的影響具有重要意義。
隨著近海生態系統富營養化程度逐漸加重,沿海水域中浮游細菌的產量、生物量和生長速率也會增加[19-20]。黃海處于亞洲沙塵源區和華北城市群的下風帶,受亞洲沙塵影響的概率較大[21]。目前有關沙塵沉降對黃海浮游細菌影響的研究多集中于遠洋[6,22],因此,本研究在去除攝食壓力的條件下,通過室內培養實驗來研究亞洲沙塵中鐵、銅對黃海近海表層優勢浮游細菌豐度的影響,為深入揭示沙塵沉降對海洋浮游細菌的影響及其作用機制提供科學參考。
1 材料與方法
1.1 培養實驗設置
本研究分別于2021年6和8月,在實驗室內進行了鐵、銅和沙塵添加培養實驗,各組物質添加濃度見表1,每組設置3個平行樣。所有用于海水取樣和培養實驗的器皿均在10%的鹽酸中浸泡24 h后用超純水清洗干凈,去除金屬污染物。海水采樣點為周圍無顯著人為污染源的黃海近岸海域(36°05′N,120°27′E),采集表層海水后,記錄采樣時間和海水溫度。將采集的海水帶回實驗室,依次用酸洗過并高溫高壓滅菌的63 μm尼龍篩絹、5 μm混合纖維濾膜和2 μm聚碳酸酯膜過濾,濾去浮游動物和浮游植物,進行高溫高壓滅菌后,分裝于1 L無菌聚碳酸酯瓶中。將優勢細菌菌懸液(1∶100)接種至600 mL的培養體系中,分別添加不同濃度(見表1)的鐵、銅和沙塵溶液混合均勻,將培養瓶置于恒溫振蕩培養箱中25 ℃、150 r·min-1避光培養168 h。

表1 各室內培養組添加物質濃度Table 1 Treatment applied to indoor culture experimental bottles
本研究中所用沙塵氣溶膠樣品是在2021年1月12日青島發生沙塵天時采集,采用KC-1000型大流量采樣器(青島嶗山電子有限公司)以1.05 m3·min-1的流量連續采樣12 h,將大氣顆粒物收集在Whatman 41#纖維濾膜上。采樣結束后對樣品進行稱重,于-20 ℃冷凍保存。在進行培養實驗之前,將一定量的沙塵氣溶膠樣品置于15 mL超純水(18.2 MΩ·cm)中,在0 ℃下超聲萃取30 min,制備沙塵萃取液,濃度為0.67和2 mg·L-1。
1.2 樣品測定
1.2.1 優勢浮游細菌豐度 室內培養周期為168 h,分別于培養的第0、3、6、12、24和36小時取樣,之后每24小時取樣一次。每次取100 μL樣品,將其涂布于平板上,置于恒溫培養箱中25 ℃避光培養72 h后,進行平板計數,計算其豐度。
1.2.2 各營養物質濃度分析 分別于培養的第0、1、3、5、7天取樣,取30 mL水樣于酸洗過的潔凈HDPE瓶中,水樣經0.45 μm濾膜過濾后,濾液于-20 ℃冷凍保存,用于營養鹽和金屬分析。同時,分別取20 mL水樣置于高溫灼燒過的棕色玻璃瓶中,經0.7 μm的GF/F濾膜過濾后,加入1滴飽和HgCl2溶液,將濾液于-20 ℃冷凍保存,用于溶解態有機碳(Dissolved Organic Carbon,DOC)分析。
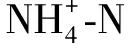
1.3 數據處理與分析方法
1.3.1 細菌豐度比(Tn/T0) 通過比較添加營養物質后各組第n時取樣的細菌豐度(CFU·mL-1)與實驗開始時的比值,將黃海近海表層優勢浮游細菌的生長進行歸一化處理[15]。
1.3.2 細菌豐度的相對變化 細菌豐度相對變化的計算方法為(A-C)/C,其中A和C分別為第n時取樣的各營養物質添加組和對照組中黃海近海表層優勢浮游細菌豐度的平均值[23]。
1.3.3 黃海表層近海優勢浮游細菌對各營養鹽的吸收(釋放)速率 單個菌落對營養鹽的吸收速率Isp[24-25](μmol·CFU-1·h-1),計算公式如下所示:
Isp=(Cn-C0)/B(tn-t0)。
(1)
式中:C0、Cn分別為培養實驗開始和取樣時的營養鹽濃度(μmol·L-1);t0、tn分別為培養實驗開始和取樣時間;B為培養期間細菌的平均含量(CFU·mL-1),細菌在此期間以指數形式增長,則B用下式進行計算:
B=(Bn-B0)/(lnBn-lnB0)。
(2)
式中B0、Bn分別為培養開始和取樣時的細菌含量(CFU·mL-1)。
2 結果與討論
2.1 初始海水營養物質濃度

2.2 FeCl3添加對黃海表層近海優勢浮游細菌豐度的影響
于2020年9月采集黃海近岸表層水,在實驗室進行培養、篩選、純化,最終得到黃海表層近海優勢浮游細菌。經鑒定該細菌為Rhodobacteraceaebacterium(α-變形菌綱)。生理上高度多樣化的Rhodobacteraceae是沿海海洋細菌的主要群體之一,可以迅速對環境變化做出反應[23,32]。Hou等[22]在黃海進行沙塵添加實驗時發現,在培養后期,Rhodobacteraceae會取代一些γ-變形菌,在海洋浮游細菌群落結構中占據優勢。因此,本研究以Rhodobacteraceaebacterium作為優勢菌代表,研究沙塵沉降對海洋表層浮游細菌的影響。
不同濃度鐵的添加對黃海表層近海優勢浮游細菌豐度均有促進作用,但影響效應有所不同(見圖1)。與對照組相比,低濃度的鐵能促進該優勢浮游細菌豐度增長,但無顯著差異(P>0.05),這與Obernosterer等[33]在克爾蓋倫群島進行的單一鐵添加(添加濃度為1 nM)的結果相似。低鐵組在培養初期,優勢浮游細菌處于對新環境的適應期,細菌豐度逐漸增加,至第12小時,低鐵組細菌豐度是對照組的3.39倍(見圖2)。在培養24 h之后,進入指數生長期,并且在第48小時細菌豐度達到最大值,低鐵添加組是對照組的1.68倍。培養中期細菌豐度開始降低,在培養的第120小時,低鐵組優勢浮游細菌豐度降至最低值,但依然高于對照組,是對照組的1.38倍。在培養后期,低鐵組優勢浮游細菌豐度再次緩慢增加,細菌豐度略高于對照組,相對增加了9%~15%。高濃度的鐵促進優勢浮游細菌豐度,但變化趨勢與對照組基本一致,沒有顯著差異(P>0.05)。高鐵組在培養初期,優勢浮游細菌增長相對較快,在培養的第12小時,高鐵添加組細菌豐度已是對照組的6.58倍,顯著促進細菌生長(P<0.05)。在培養的第72小時,優勢浮游細菌豐度達到最大值,是對照組的1.33倍。與對照組相同,高鐵組細菌豐度達到峰值后逐漸降低,在培養的第120小時,細菌豐度降至最低值,比對照組下降41%。在培養后期,細菌緩慢增長,而細菌豐度逐漸增加,高鐵組細菌豐度比對照組增加了2%~15%。由此可見,不同濃度鐵的添加均能延后黃海表層近海優勢浮游細菌豐度峰值的出現,但峰值豐度均高于對照組,并且可以在培養前期短暫地刺激黃海表層近海優勢浮游細菌的生長,呈現明顯的促進作用,這與Westrich等[15]在佛羅里達群島的一系列沙塵添加對弧菌生長的實驗結果相似。同時,Westrich等[15]還發現,在沙塵添加的24 h內,與對照組相比,各添加組弧菌豐度均顯著增加,且其豐度增加量主要與沙塵濃度有關;在最小沙塵添加量(溶解鐵含量為0.02 nmol·L-1),弧菌平均生長速度為對照組的6倍。本研究中,在添加0.5 μmol/L無機鐵后,黃海表層近海優勢浮游細菌豐度在第12小時才增加到對照組的6.58倍,其生長速度遠低于Westrich等研究中的溶藻弧菌和霍亂弧菌,這與不同細菌種類的自身特性、生長所需的營養物質、對外源營養物質的響應以及海水背景情況等因素有關[23,32]。
黃海近海磷酸鹽濃度較低,優勢細菌生長可能受到磷酸鹽的限制,鐵的添加使培養體系中的N/P值明顯降低(見圖3),促進細菌消耗DIN,釋放出部分磷酸鹽,緩解了培養體系的磷限制,從而在培養前期刺激優勢細菌豐度增加。前期細菌快速生長,消耗了體系中的營養物質,后期體系中的N/P值逐漸增大,可能是由于營養缺乏導致細菌生長減慢,豐度下降,優勢細菌礦化出無機氮。在新西蘭近海的室內培養實驗結果表明,異養細菌豐度對單獨添加鐵的響應很小,但在近海淺水區混合添加Co、Mn、Cu、Fe、Ni,可顯著促進異養浮游細菌豐度增加,這表明新西蘭近海細菌生長受多種金屬的共同限制[34]。Hou等[22]在黃海進行的船基圍隔實驗結果表明,單獨添加0.02 μmol/L的鐵可顯著促進海洋浮游細菌生物量的增加,表明黃海開闊海域浮游細菌生長受鐵限制。由于不同海洋細菌細胞營養成分以及對營養物質的吸收率不同,開闊海域和沿海水域會出現不同的營養限制[35-37]。因此,即使在黃海近海鐵濃度較高情況下,外源輸入的鐵仍然可以在短期內促進Rhodobacteraceaebacterium生長,同時減輕培養體系中的磷限制,添加鐵的濃度越高,這種影響越明顯。
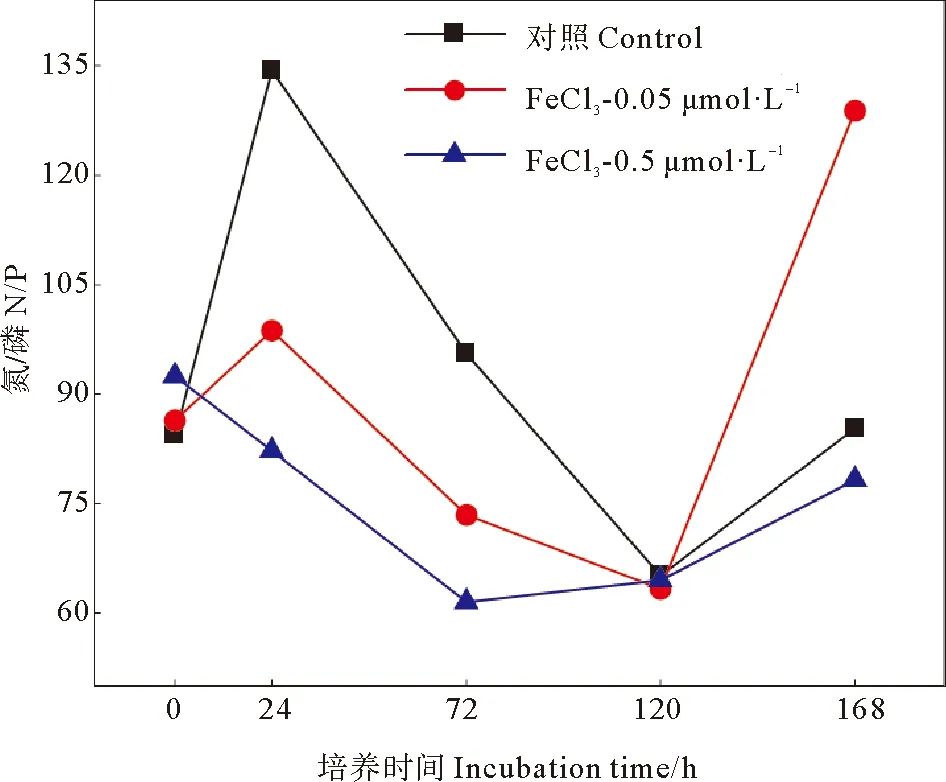
圖3 添加FeCl3后各組N/P變化

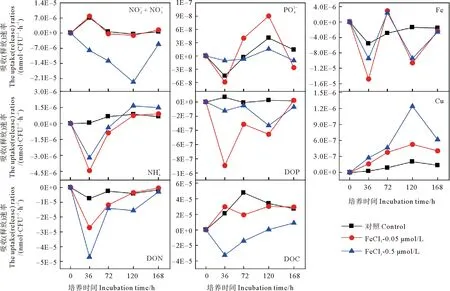
圖4 添加FeCl3后各組中黃海表層近海優勢浮游細菌對各營養鹽的吸收(釋放)速率
2.3 CuCl2添加對黃海表層近海優勢浮游細菌豐度的影響
不同濃度銅的添加對黃海表層近海優勢浮游細菌豐度影響結果如圖5所示。結果顯示,添加銅對細菌豐度有一定抑制,但與對照組并無顯著差異(P>0.05)。在培養初期,低銅添加組細菌豐度逐漸增加,但略低于對照組。在培養的第24小時,低銅組細菌豐度比對照組降低了29%(見圖6);在培養的第36小時低銅添加組細菌豐度達到峰值,但仍低于對照組。達到峰值后,低銅組細菌豐度開始降低,至第72小時降至最低值,比對照組降低了18%。之后低銅組細菌豐度開始緩慢增加,在第120小時達到第二個峰值,但此期間豐度均低于對照組,比對照組降低了10%~26%。這表明,低濃度銅的加入,對該優勢浮游細菌的生長呈現一定的抑制作用。Baltar等[34]在新西蘭沿海單獨添加7 nmol·L-1銅的研究同樣發現,海洋異養細菌豐度無顯著變化。高銅添加組的細菌豐度變化趨勢與對照組基本一致,整體表現為先升高后降低再升高的復雜變化。在培養初期,高濃度銅的加入對該優勢浮游細菌的生長表現出一定的抑制作用,高銅組細菌豐度比對照組降低了4%~8%。隨著培養時間的增加細菌豐度逐步增加,培養的第24小時,高銅組細菌豐度為對照組的1.46倍。與對照組不同,高銅組細菌豐度在培養的第48小時達到第一個峰值,但該峰值仍低于對照組,為對照組的94%。之后高銅組細菌豐度逐漸降低,在培養的第72小時降至最低值,比對照組低53%。培養后期,高銅組細菌豐度也在第120小時達到第二個峰值,細菌豐度仍低于對照組。總的來說,高濃度銅的添加對黃海表層近海優勢浮游細菌生長的抑制作用高于低濃度銅的添加,影響更為復雜,這可能與細菌對銅生物利用性的響應以及其細胞銅配額有關[41]。由圖7可知,在細菌指數生長時,銅的添加使培養體系中的N/P值降低,減輕了體系中的磷限制,而優勢細菌豐度與對照組相近,甚至低于對照。由此可見,在體系中銅濃度相對較高的情況下,盡管銅的添加可以緩解磷限制,但銅的添加仍會抑制優勢細菌生長,這可能與銅的毒性作用有關。

((a)培養168 h內各組細菌豐度比;(b)培養12 h內各組細菌豐度比。(a)Bacterial abundance ratio of each group within 168 h of culture;(b)Bacterial abundance ratio of each group within 12 h of culture.)
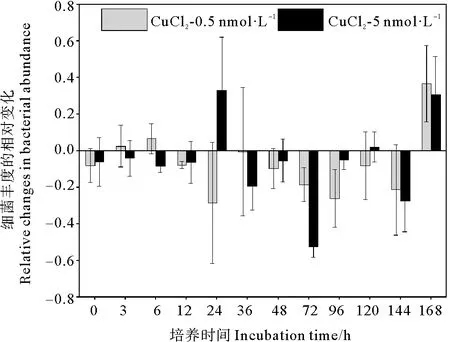
圖6 添加CuCl2后各組細菌豐度的相對變化
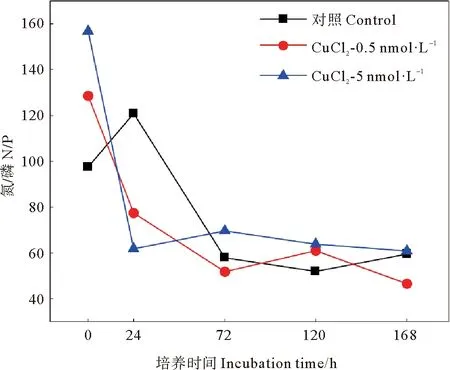
圖7 添加CuCl2后各組N/P變化
2.4 沙塵添加對黃海表層近海優勢浮游細菌豐度的影響
沙塵添加培養實驗中黃海表層近海優勢浮游細菌豐度的變化如圖9所示。低濃度沙塵組與對照組優勢細菌豐度變化趨勢相似,對浮游細菌豐度具有一定促進作用,在第144小時存在顯著差異(P<0.05)。在培養初期,低沙塵組細菌豐度逐漸增加,但比對照組低5%~13%(見圖10)表現出輕微的抑制作用。而在培養的第12、72小時,低沙塵添加組細菌豐度高于對照組,相對增加了11%和16%。該組細菌豐度在培養的第144小時達到最大值,比對照組高60%,此時,低濃度沙塵顯著刺激細菌生長(P<0.05)。高沙塵添加組優勢浮游細菌豐度變化趨勢與低沙塵添加組相似,無顯著差異(P>0.05)。培養初期,高沙塵組細菌豐度逐漸增加,但比對照組低11%~19%。在培養中期,高濃度沙塵的添加主要促進了優勢浮游細菌生長,細菌豐度約為對照組的1.01~1.2倍。培養后期,高濃度沙塵對優勢細菌的生長具有較強的促進作用,細菌豐度為對照組的1.38~1.48倍。可見,沙塵添加對黃海表層近海浮游細菌的影響主要是初期抑制、后期促進。培養前期,沙塵的添加使得培養體系中N/P值增大(見圖11),這可能是由于沙塵攜帶的鐵、銅對優勢細菌利用氮、磷的影響較小或者沙塵釋放的無機氮濃度較高,磷酸鹽濃度較低,從而加重了體系中的磷限制,對細菌生長有輕微的抑制作用。而后期,N/P值進一步增大,但細菌豐度卻顯著增加,這表明磷不再是影響優勢細菌生長的主要限制因子,可能是沙塵顆粒中其他復雜的化學組分或者多種營養物質共同限制。
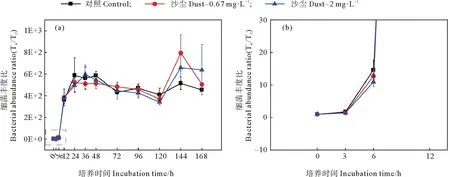
((a)培養168 h內各組細菌豐度比;(b)培養12 h內各組細菌豐度比。(a)Bacterial abundance ratio of each group within 168 h of culture;(b)Bacterial abundance ratio of each group within 12 h of culture.)

圖10 添加沙塵后各組細菌豐度的相對變化

圖11 添加沙塵后各組N/P變化
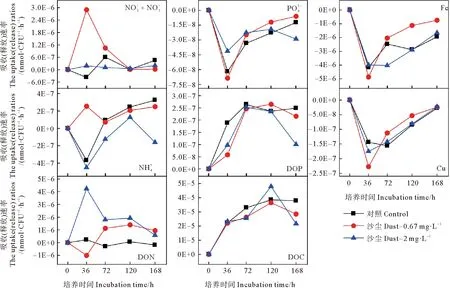
圖12 添加沙塵后各組中黃海表層近海優勢浮游細菌對各營養鹽的吸收(釋放)速率
本研究中,黃海表層近海優勢浮游細菌(Rhodo-bacteraceaebacterium)對無機鐵、銅以及沙塵的添加具有一定響應,與對照組相比,低鐵組(第12小時)以及低沙塵組(第144小時)均可顯著促進細菌生長。這表明,沙塵濃度與海洋浮游細菌生長緊密相關,在一定的閾值內,沙塵沉降對海洋細菌的豐度、活性和生產等會產生積極的影響[47]。在黃海進行的船基圍隔培養實驗顯示,單獨添加鐵及沙塵顯著促進海洋浮游細菌生長[7,34],而在北大西洋進行的沙塵添加實驗表明,沙塵攜帶的無機鐵并不是刺激弧菌生長的主要因子[48],因此,不同海區營養限制不同,沙塵沉降對海洋細菌的影響也不同。
沙塵沉降帶來的外源營養物質可以直接影響海洋細菌生長,而Rhodobacteraceae類群與微型浮游植物密切相關[49],因此沙塵還可能通過刺激浮游植物生長[15],間接影響本研究中優勢細菌的豐度。黃海近岸由于受人類活動影響,各營養物質濃度高于遠洋[26],這也使得外源營養物質的輸入對該優勢浮游細菌生長產生的影響不如大洋顯著,同時,培養前期沙塵攜帶的營養物質加劇了體系中的磷限制,因此優勢細菌對沙塵的響應較小。Rahav等[50]在地中海低營養、低葉綠素海域四年現場研究中發現,在超寡營養海域,異養細菌豐度和細菌生產力增加更快,且生態系統營養越寡,沙塵添加對細菌豐度以及生產力的影響越明顯。但本研究結果表明,即便在近岸高金屬、高營養鹽水體,沙塵輸入仍對細菌豐度有一定影響,尤其在培養后期有顯著促進作用,同時影響海洋細菌對營養鹽、DOC和微量金屬的利用。此外,沙塵沉降對近海浮游細菌豐度以及利用各營養物質的影響,仍然可能改變近海浮游細菌群落結構和代謝平衡,進而對海洋碳和營養物質循環產生重要影響[51-54]。
3 結論
(1)外源鐵可以在培養前期內促進黃海表層近海優勢浮游細菌(Rhodobacteraceaebacterium)豐度的增加,促進優勢細菌對氮、磷、DOC和Fe的吸收利用,同時緩解體系中的磷限制。
(3)在近海富營養狀態下,沙塵輸入在前期對黃海表層近海優勢浮游細菌生長有輕微的抑制作用,培養后期則促進優勢細菌豐度增加。低濃度沙塵主要促進該優勢浮游細菌對DON、Fe和Cu的吸收以及對DIN的釋放,而高濃度沙塵則主要促進優勢浮游細菌對DON的釋放。沙塵沉降會加劇黃海近海的磷限制,對近海富營養海域的浮游細菌生長有明顯影響。
猜你喜歡
我愛學·笑話與口才(2025年3期)2025-02-24 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
趣味(語文)(2020年3期)2020-07-27 01:42:46
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
作文與考試·初中版(2017年12期)2017-04-19 20:26:27
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
中國衛生(2015年2期)2015-11-12 13:13:54
