古老月季葉綠體基因組密碼子分析
2023-10-24 10:36:32程培蕾嚴陶韜高靜瑤張彤鈺
江蘇農業科學 2023年17期
程培蕾 嚴陶韜 高靜瑤 張彤鈺
摘要:我國古老月季月月粉具有連續開花、重瓣和標志性茶香等優良性狀,是重要的育種材料。為明確月月粉葉綠體基因組密碼子的特點,從NCBI網站獲得了完整的葉綠體基因組信息,并對所有基因的密碼子使用偏性進行計算分析。結果表明,月月粉葉綠體包含53個基因,1~3位密碼子GC含量平均值分別為47.08%、39.54%、29.14%,絕大多數密碼子前2位的GC含量較高。編碼區有效密碼子數平均值為46.86,表明其偏性較弱。相關性分析顯示密碼子第3位與前2 位存在明顯差異,可能對密碼子使用偏性具有關鍵作用。29個RSCU>1的密碼子中以A、U 結尾的有28個。中性繪圖分析表明,GC12與GC3間相關系數為0.088,相關性不顯著,ENC-plot分析、PR2-plot分析結果表明自然選擇在月月粉葉綠體基因組密碼子使用模式中起決定性作用,并篩選到16個最優密碼子。結果可為月季遺傳轉化過程中提高外源基因的表達提供有用信息。
關鍵詞:月季;葉綠體基因組;密碼子偏好性;最優密碼子
中圖分類號:S685.120.1 文獻標志碼:A 文章編號:1002-1302(2023)17-0034-07
三聯體密碼子通過簡并性方式編碼氨基酸[1],由多個同義密碼子編碼相同氨基酸的現象極其普遍,僅由單個密碼子編碼的氨基酸數量極少,已知的只有蛋氨酸(Met)和色氨酸(Trp)[2]。不同植物基因組中任意氨基酸同義密碼子的選擇是非隨機的,這被稱為同義密碼子使用偏好性[3]。密碼子使用偏好性不僅因物種、家族或生物界內的群體而異,也因生物體內基因的不同而異。自然選擇和突變在生物體密碼子使用偏好性方面起著決定性作用[4-5]。密碼子使用偏好性在多種生命過程中發揮作用,例如蛋白質翻譯和折疊等過程,也反映了物種或基因的起源、突變模式和進化[6-7]。對密碼子使用偏好性的研究可以揭示生物之間的系統發育關系,水平基因轉移,基因的分子進化,確定驅動其進化的影響因素,并且對外源基因密碼子的優化可以在一定程度上提高轉基因育種中的外源基因表達水平[8]。
多數植物葉綠體基因組高度保守,廣泛應用于植物間的進化規律、遺傳關系、葉綠體基因表達和葉綠體轉化研究[9]。近年來,高通量測序技術豐富了葉綠體基因組數據庫,與植物基因組結構和系統發育分析相關的研究也更加廣泛和深入。越來越多的園藝作物開展了葉綠體基因組密碼子使用偏好性分析,例如山茶、睡蓮、含笑、連翹、云南油杉、木薯等[10-15]。
月季(Rosa chinensis)是栽培品種極其豐富的木本花卉,不僅具有很高的觀賞價值,更可作為食品添加劑、熏蒸劑、化妝品和傳統醫療產品使用[16-17]。我國古老月季月月粉是重要的育種材料,具有多種優良性狀,包括連續開花、重瓣和標志性茶香[18]。隨著其全基因組序列的公布[19-20],月月粉成為研究木本植物花發育、花顏色、氣味釋放和春化反應分子機制的理想模式物種。
本研究從月季葉綠體編碼序列的角度,系統分析了古老月季月月粉葉綠體基因組的密碼子組成、密碼子偏好性情況及影響因素,并篩選出最優密碼子。本研究揭示了月月粉密碼子使用偏好性,以期為進一步研究月季的遺傳進化和轉基因提供參考方向。
1 材料與方法
1.1 材料
2022年8月于蘇州農業職業技術學院進行相關數據分析。月季月月粉完整的葉綠體基因組從NCBI(https://www.ncbi.nlm.nih.gov)網站獲得,GenBank登錄編號為MH332770。葉綠體基因組大小為156 591 bp,根據各基因的注釋結果獲得不重復的85條CDS(coding DNA sequence)。對獲得的原始編碼序列進行篩選分析,保留了序列長度 300 bp 以上,起始為ATG、結尾為TAG、TGA、TAA的53條非重復CDS用于下一步分析。
1.2 方法
1.2.1 密碼子組成分析 參照梁湘蘭等的方法統計了53條月月粉葉綠體基因密碼子數量和不同位點的GC含量[21]。有效密碼子數 (effective number of codon,ENC)測定參考李魁印等的方法,ENC數值用以表征同義密碼子使用偏度,數值區間為20~61,ENC值=20表示偏度最高,隨數值增加偏度變弱[22]。RSCU(relative synonymous codon usage)指相對同義密碼子使用度,當數值高于1時表示實際使用頻率高于其他密碼子,確定為高頻密碼子,其計算方法參照文獻[23]。
1.2.2 密碼子中性繪圖分析 月月粉葉綠體基因組密碼子中性繪圖的橫、縱坐標分別為GC3值和GC1、GC2平均值,散點表示相應的基因。橫縱坐標間的相關關系代表了密碼子偏好性的主要影響因素,當兩者呈現顯著相關時,不同位點的堿基使用差別不明顯,突變成為決定性因素;而當兩者呈現不顯著相關時,堿基組成差異性與回歸系數呈負相關關系,自然選擇成為決定性影響因素,繪圖方法參照文獻[24]。
1.2.3 ENC-plot分析 ENC-plot分析參考楊祥燕等的方法[25]。ENC-plot繪圖可以表征突變和自然選擇在密碼子偏好性中的作用,以散點到標準曲線的距離為基本劃分依據,當二者距離近時代表偏好性主要以突變影響為主,而當兩者距離相對較遠時,偏好性則主要取決于自然選擇對其的影響。以ENC比值頻數的分布情況來量化突變和自然選擇的影響,其比值具體計算公式為ENC差值(ENC期望值-ENC實際值)與ENC 期望值之比,其中,ENC期望值基本計算公式:2+GC3+29/[GC23+(1-GC3)2][25]。
1.2.4 PR2-plot分析 統計月月粉53個葉綠體基因各密碼子第 3 位點的A、T、C、G分布情況,PR2偏倚分析(PR2-bias plot analysis)圖中,A3/(A3+T3)與G3/(G3+C3)的取值分別被設置為縱、橫坐標,縱橫坐標的中心交界點數值為0.5,該點A、T相同,C、G也相同,即密碼子無偏性使用,其他散點距離中心點的矢量距離代表偏好程度[25-26]。
1.2.5 最優密碼子篩選 將ENC值由低到高排序,最低和最高的10%分別作為月月粉葉綠體基因高、低偏性庫,RSCU>1表示高頻密碼子,高、低偏性庫間差值ΔRSCU(高表達RSCU-低表達RSCU)在0.08及以上時表示高表達優越密碼子,最優密碼子的篩選標準為高頻率密碼子和高表達優越密碼子的交集[27]。
2 結果與分析
2.1 月月粉密碼子偏好性相關分析
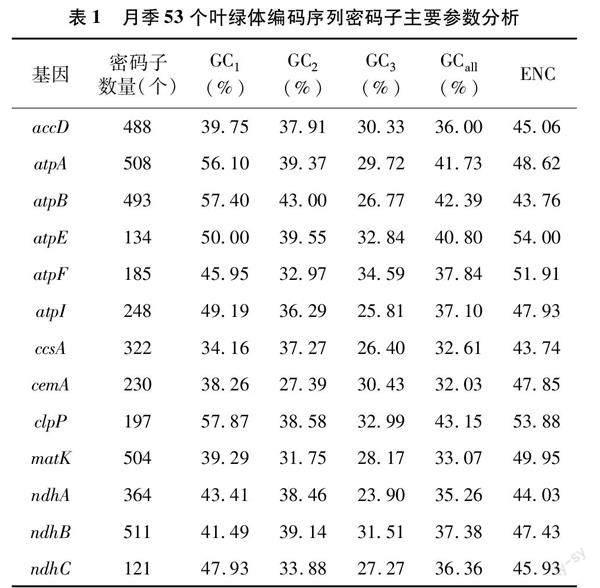
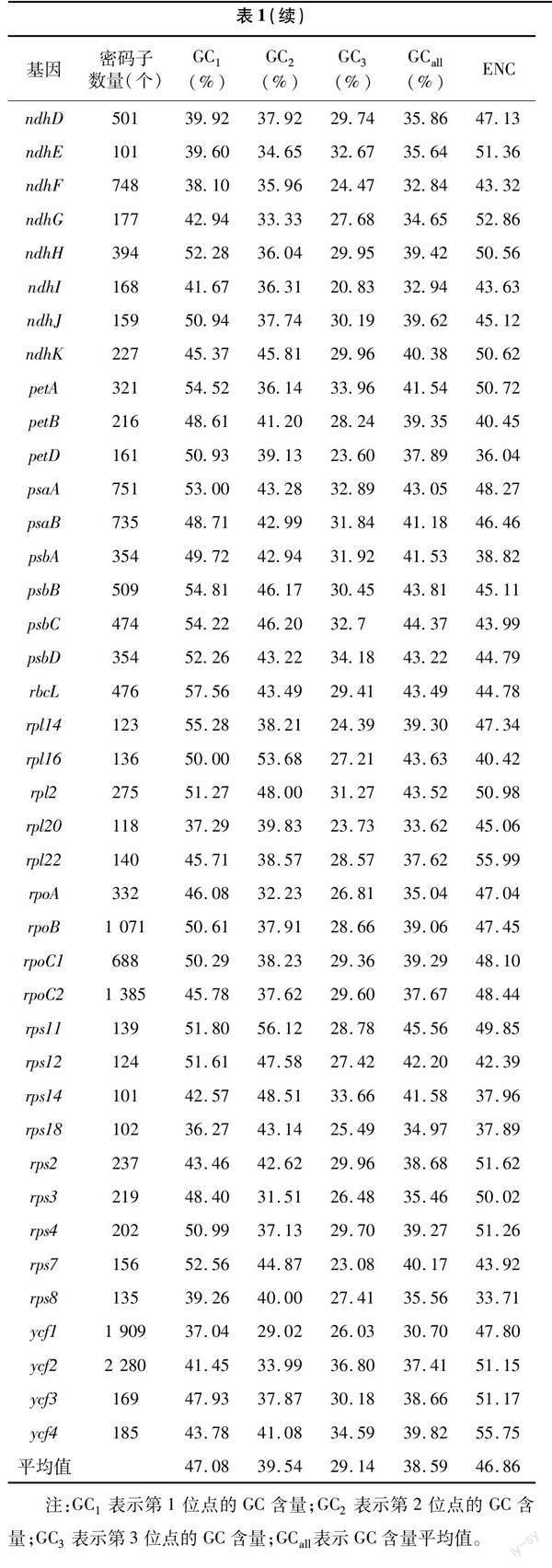
月季葉綠體基因組序列下載自NCBI 數據庫(GenBank登錄號:MH332770),53個葉綠體基因的密碼子偏好性相關參數分析結果見表1。其密碼子的GC含量平均值為38.59%,其中,密碼子由第1位點至第3位點的GC含量平均值分別為47.08%、39.54%、29.14%,1位GC含量最高的基因clpP為57.87%,2位GC含量最高的基因rps11為56.12%,第3位點GC含量最高的基因ycf2為36.80%,可見,密碼子GC含量在不同位點差別較大,且絕大多數基因密碼子前2位的GC含量較高,第3位的GC含量較低,表明葉綠體基因更加偏好使用A 或U結尾。此外,月季53個葉綠體基因ENC平均值為46.86,最大值和最小值分別為55.99和33.71,月月粉53個編碼序列密碼子的主要參數分析結果表明密碼子偏性較弱。
對月季葉綠體基因密碼子參數的相關性分析結果(圖1)表明,GC1與GC2極顯著正相關,GC1與GC3以及GC2與GC3均不顯著相關,GCall與3個位點的GC值(GC1、GC2、GC3)均為極顯著正相關,表明第1位與第2位堿基呈現極高的相似度,但這兩者與第3 位均存在較大差異;ENC與GC1無顯著相關關系,與GC2和GC3相關性較強,分別為顯著和極顯著相關,猜測密碼子使用的偏好性主要受到GC3含量的影響;密碼子數與其他參數均不顯著相關,說明月季葉綠體中密碼子堿基組成不影響密碼子數。
月月粉53個葉綠體基因RSCU分析結果見圖2,這些基因由 21 304個非終止密碼子組成。Ile(AUA)的RSCU值為0.925 2,Leu(CUA)的RSCU值為0.748 8,其他以A或U結尾的密碼子RSCU值均大于1.0。共篩選到29個RSCU值大于1.0的密碼子,其中以U結尾的有16個、以A 結尾的有12個、以G結尾的有1個。表明月月粉葉綠體基因密碼子更加偏向于在末位選擇使用A或U。
2.2 密碼子堿基組成的差異性
中性繪圖分析結果(圖3)表明,月季葉綠體基因組GC分布較窄,GC3 的變化范圍為20.83%~36.80%,GC12 的取值范圍為32.83%~53.96%。GC12與GC3的相關系數為r=0.08 8(r2=0.007 8),相關性不顯著,且基因均位于對角線的上方,猜測自然選擇作為影響密碼子偏好性的主要原因發揮作用。以基因的功能為依據對其進行分類,53條序列可以進一步分為15類,其中保守的假定葉綠體ORF(conserved hypothetical chloroplast,ORF)和被膜蛋白(envelope membrane protein)靠近對角線, 其他基因均距離對角線較遠。
2.3 ENC-plot繪圖分析
對葉綠體基因的密碼子進行ENC-plot分析(圖4),標準曲線下方聚集了大多數葉綠體基因,且距離較遠,標準曲線兩側僅均勻分布了部分基因,表示月季葉綠體基因組密碼子偏好性受突變的影響較小,受自然選擇的影響較大。對月季葉綠體基因的ENC頻數分布進行了統計(表2),有17個基因分布在-0.05~0.05區間,基因頻率占比為32%,有25個基因分布在0.05~0.15區間,基因頻率占比為47%,表明實際的ENC數值與預期值之間差距較大,因此主要是自然選擇影響了月月粉葉綠體基因組密碼子的偏好性。
2.4 月季葉綠體基因組PR2-plot繪圖分析
對月季53個葉綠體基因進行PR2-plot 繪圖分析(圖5),結果表明密碼子使用情況為A3/A3+T3(平均值為0.460)低于G3/G3+C3(平均值為0.543)。這些基因在平面4個區域內呈不均勻分布,其中右下角區域基因數量最多,而左上角區域基因數量最少。此外,4種堿基在使用頻率方面也存在差異,G的使用頻率高于C,T的使用頻率高于A;且基因小核糖體亞基蛋白和光系統Ⅱ亞單位偏離中心點(A=T且C=G)的矢量距離最遠。嘧啶與嘌呤間的使用頻率差異也進一步證明, 自然選擇決定了月月粉葉綠體基因組密碼子使用模式,而突變在這一過程中發揮的作用較弱。
2.5 最優密碼子確定
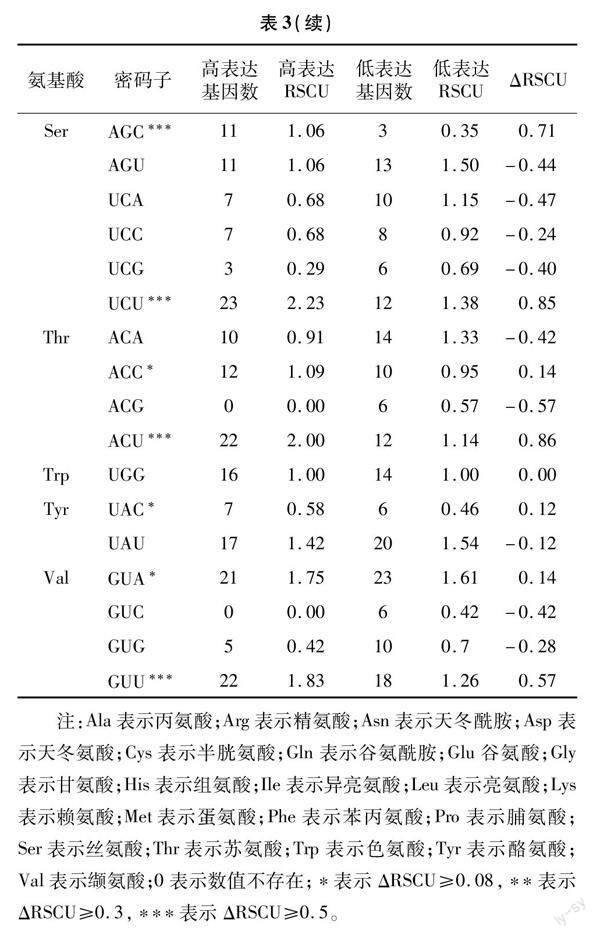
月季53個葉綠體基因的高、低偏性庫,以及2個庫的RSCU值統計結果見表3。高表達優越密碼子的篩選標準為高、低偏性庫之間差值在0.08以上, 共篩選到21個。進一步分析了這21個高表達優越密碼子的末位堿基情況,絕大多數(14個)以A/U結尾,以C結尾的次之(6個),以G結尾的最少(僅1個)。將RSCU>1的密碼子作為高頻密碼子,共篩選到29個,其中28個以A/U 結尾,僅1個以G 結尾。高頻密碼子與高表達優越密碼子的交集作為月月粉最優密碼子,本研究中共獲得了16個最優密碼子,分別為GCA、GCU、CGU、UGU、CAA、GAA、GGU、AUU、UUA、UUG、AGC、UCU、ACC、ACU、GUA、GUU,這些最優密碼子第3 位堿基有5個為A,8個為U,2個為C,1個為G。
3 討論與結論
古老月季月月粉葉綠體基因組的密碼子使用偏好性對于探討其進化過程至關重要,有助于該物種的分子育種應用。盡管先前已經有關于大花香水月季葉綠體基因組密碼子使用特點的相關研究,兩者均為二倍體多年生植物,但其基本性狀差異較大,最典型的差異為月月粉可以周年連續開花,而大花香水月季為早春一次開花型[28-29]。此外隨著全基因組信息的公布,月月粉成為研究木本植物花發育、花顏色、氣味釋放和春化反應分子機制的理想模式物種[19-20]。本研究旨在系統分析古老月季月月粉葉綠體基因組的有效密碼子數、相對同義密碼子使用、密碼子偏好性及其影響因素,這將為優化月月粉分子育種提供一定參考。
同義密碼子使用偏好性是生物類群中常見的現象,一般指不同物種之間或同一物種基因組內密碼子出現頻率的差異[30]。密碼子第3位堿基的同義突變不能改變氨基酸的類型,但仍被認為是決定氨基酸類型的一個重要特征,且其GC含量決定了密碼子偏好性[31-32]。本研究中,月月粉葉綠體基因組GC含量平均值為38.59%,密碼子第3位的GC含量為29.14%,密碼子末位堿基使用A、U的頻率高于G、C,在多個物種中存在相似的現象,包括菠蘿[25]、苦馬豆[21]、文心蘭[33]等。對密碼子使用模式的兩大主要影響因素(堿基突變和自然選擇)進行確認,當突變為絕對原因時,則應按比例使用4個堿基(A、C、G 和 T)[34]。然而,月月粉葉綠體基因組中的大多數編碼序列都是A、U結尾,這表明自然選擇是對其密碼子使用模式的更大影響因素。
中性理論認為堿基突變和自然選擇以中性或接近中性的形式影響了密碼子第3個堿基的使用[4]。如果自然選擇作為主要的影響因素,則第3個位點的GC含量往往分布在一個小范圍內,且與前2位沒有顯著相關性[35]。在本研究的中性分析結果顯示,月月粉葉綠體基因組的密碼子GC3 的變化范圍較小(20.83%~36.80%),GC12 與GC3 的相關系數為r=0.088(r2=0.007 8),相關性不顯著,表明其密碼子使用偏好性主要受自然選擇的影響。此外,月月粉葉綠體ENC頻數比值結果顯示,其主要的分布區間為0.05~0.15,實際數值與預期數值之間存在較大的差別[25];PR2-plot分析結果中的嘌呤A/G與嘧啶T/C間的差異都表明自然選擇在月月粉密碼子使用偏好性中的決定性作用,而突變發揮的作用較弱[24-28]。自然選擇、突變等因素的作用,促進了密碼子使用偏性的發生,而最優密碼子形成的重要條件就是強烈的正向選擇和突變壓力[36]。本研究篩選到16個最優密碼子,這些最優密碼子將為月月粉遺傳轉化過程中提高基因的表達效率提供有用信息。
參考文獻:
[1]Grantham R,Gautier C,Gouy M,et al. Codon catalog usage and the genome hypothesis[J]. Nucleic Acids Research,1980,8(1):49-62.
[2]Guan D L,Ma L B,Khan M S,et al. Analysis of codon usage patterns in Hirudinaria manillensis reveals a preference for GC-ending codons caused by dominant selection constraints[J]. BMC Genomics,2018,19(1):542.
[3]Wright F. The ‘effective number of codons used in a gene[J]. Gene,1990,87(1):23-29.
[4]Sharp P M,Stenico M,Peden J F,et al. Codon usage mutational bias,translational selection,or both?[J]. Biochemical Society Transactions,1993,21(4):835-841.
[5]Lesnik T,Solomovici J,Deana A,et al. Ribosome traffic in E.coli and regulation of gene expression[J]. Journal of Theoretical Biology,2000,202(2):175-185.
[6]Supek F. The code of silence:widespread associations between synonymous Codon biases and gene function[J]. Journal of Molecular Evolution,2016,82(1):65-73.
[7]Athey J,Alexaki A,Osipova E,et al. A new and updated resource for codon usage tables[J]. BMC Bioinformatics,2017,18(1):391.
[8]Parvathy S T,Udayasuriyan V,Bhadana V. Codon usage bias[J]. Molecular Biology Reports,2022,49(1):539-565.
[9]Wicke S,Schneeweiss G M,DePamphilis C W,et al. The evolution of the plastid chromosome in land plants:gene content,gene order,gene function[J]. Plant Molecular Biology,2011,76(3):273-297.
[10]Wang Z J,Cai Q W,Wang Y,et al. Comparative analysis of codon bias in the chloroplast genomes of Theaceae species[J]. Frontiers in Genetics,2022,13:824610.
[11]毛立彥,黃秋偉,龍凌云,等. 7種睡蓮屬植物葉綠體基因組密碼子偏好性分析[J]. 西北林學院學報,2022,37(2):98-107.
[12]周 濤,楊 林,舒軍霞,等. 3種含笑屬植物葉綠體基因組密碼子偏好性分析[J]. 西部林業科學,2022,51(3):91-100.
[13]宋 蕓,賈孟君,曹亞萍,等. 連翹葉綠體基因組特征分析[J]. 園藝學報,2022,49(1):187-199.
[14]李江飛,王 瑜,顏廷雨,等. 云南油杉葉綠體基因組密碼子偏好性分析[J]. 中南林業科技大學學報,2022,42(4):30-39.
[15]趙 英,付海天,金 剛,等. 木薯葉綠體基因組密碼子偏好性分析[J]. 江西農業學報,2022,34(4):46-52.
[16]Fougère-Danezan M,Joly S,Bruneau A,et al. Phylogeny and biogeography of wild roses with specific attention to polyploids[J]. Annals of Botany,2015,115(2):275-291.
[17]Friedman H,Rot I,Agami O,et al. Edible flowers:new crops with potential health benefits[J]. Acta Horticulturae,2007(755):283-290.
[18]Bendahmane M,Dubois A,Raymond O,et al. Genetics and genomics of flower initiation and development in roses[J]. Journal of Experimental Botany,2013,64(4):847-857.
[19]Saint-Oyant L H,Ruttink T,Hamama L,et al. A high-quality genome sequence of Rosa chinensis to elucidate ornamental traits[J]. Nature Plants,2018,4(7):473-484.
[20]Raymond O,Gouzy J,Just J,et al. The Rosa genome provides new insights into the domestication of modern roses[J]. Nature Genetics,2018,50(6):772-777.
[21]梁湘蘭,郭 松. 苦馬豆葉綠體基因組密碼子偏好性分析[J]. 西北林學院學報,2022,37(2):121-126.
[22]李魁印,張海玲,張 鴻,等. 蕁麻葉綠體基因組密碼子偏好性分析[J]. 種子,2022,41(2):51-58.
[23]何亞玲,彭業軍,李 錦,等. 天山雪蓮葉綠體基因組密碼子偏好性分析[J]. 石河子大學學報(自然科學版),2022,40(1):84-92.
[24]原曉龍,李云琴,張勁峰,等. 降香黃檀葉綠體基因組密碼子偏好性分析[J]. 廣西植物,2021,41(4):622-630.
[25]楊祥燕,蔡元保,譚秦亮,等. 菠蘿葉綠體基因組密碼子偏好性分析[J]. 熱帶作物學報,2022,43(3):439-446.
[26]原曉龍,劉 音,康洪梅,等. 蒜頭果葉綠體基因組密碼子偏好性分析[J]. 西南林業大學學報(自然科學),2021,41(3):15-22.
[27]羅 茜,方 正,余 婕,等. 圓錐南芥葉綠體基因組特征及密碼子偏好性分析[J]. 分子植物育種,2022,20(7):2261-2270.
[28]牛 元,徐 瓊,王崳德,等. 大花香水月季葉綠體基因組密碼子使用偏性分析[J]. 西北林學院學報,2018,33(3):123-130.
[29]Guo X L,Yu C,Luo L,et al. Comparative transcriptome analysis of the floral transition in Rosa chinensis ‘Old Blush and R.odorata var. gigantea[J]. Scientific Reports,2017,7(1):6068.[HJ2mm]
[30]Wei L,He J,Jia X,et al. Analysis of codon usage bias of mitochondrial genome in Bombyx mori and its relation to evolution[J]. BMC Evolutionary Biology,2014,14(1):262.
[31]Ingvarsson P K. Gene expression and protein length influence codon [JP+3]usage and rates of sequence evolution in Populus tremula[J].Molecular Biology and Evolution,2007,24(3):836-844.
[32]Ye Y,Ni Z,Bai T,et al. The analysis of chloroplast genome codon usage bais in Pinus massoniana[J]. Genomics and Applied Biology,2018,37(10):4464-4471.
[33]李冬梅,呂復兵,朱根發,等. 文心蘭葉綠體基因組密碼子使用的相關分析[J]. 廣東農業科學,2012,39(10):61-65.
[34]Liu H B,Lu Y Z,Lan B L,et al. Codon usage by chloroplast gene is bias in Hemiptelea davidii[J]. Journal of Genetics,2020,99(1):1-11.
[35]Kawabe A,Miyashita N T. Patterns of codon usage bias in three dicot and four monocot plant species[J]. Genes & Genetic Systems,2003,78(5):343-352.
[36]Hershberg R,Petrov D A. Selection on codon bias[J]. Annual Review of Genetics,2008,42:287-299.
收稿日期:2022-12-29
基金項目:江蘇省高等學校基礎科學(自然科學)研究面上項目(編號:21KJB210006);江蘇省種業振興“揭榜掛帥”項目[編號:JBGS(2021)091];江蘇省林業科技創新與推廣項目[編號:LYKJ(2021)02];蘇州農業職業技術學院博士提升計劃科研啟動基金項目(編號:BS2103)。
作者簡介:程培蕾(1990—),女,山東蒙陰人,博士,講師,從事觀賞植物遺傳育種與分子生物學研究。E-mail:plcheng@szai.edu.cn。
