塔什庫爾干羊FecB 基因非探針法高分辨率熔解曲線分型方法的研究
2023-10-26 02:56:16張云生鄭新寶阿依努爾亞森
中國畜禽種業 2023年9期
關鍵詞:檢測
李 浩,郭 濤,陳 童,2,張 璐,張云生,2,鄭新寶,2,阿依努爾·亞森,2*
[1.新疆畜牧科學院,新疆烏魯木齊 830011;2.農業農村部畜禽資源(羊)評價利用重點實驗室,新疆烏魯木齊 830011;3.新疆維吾爾自治區動物衛生監督所,新疆烏魯木齊 830026]
多胎性狀是綿羊繁殖率的重要指標,多胎基因作為控制綿羊產羔數量的主效基因,發揮了重要作用。影響綿羊多胎性狀的候選基因主要有骨形態發生蛋白受體-1B(BMPR-1B)、形態發生蛋白15(BMP15)、生長分化因子9(GDF9)、親吻素(KISS-1)、骨形態發生蛋白4(BMP4)、視黃酸受體γ(RARG)、視黃醇結合蛋白4(RBP4)等。其中,BMPR-1B 基因的FecB突變型對綿羊的排卵數具有累加效應,即每增加一個拷貝將額外多排卵1.5~3.0 枚,已成為綿羊高繁殖力標記基因之一[1]。
綿羊多胎性狀由多基因調控。BMPR-1B 基因的突變體FecB基因是一個常染色體基因,可通過共顯性效應和部分顯性效應提高排卵率,在提高綿羊排卵數和產羔數等方面發揮了重要作用。FecB基因作為綿羊多胎性狀的主效基因,具有豐富的遺傳多樣性[2],不同品種的綿羊廣泛地存在FecB基因,可以作為選育肉用綿羊多胎類型的有效候選基因。我國的湖羊、小尾寒羊、多浪羊、策勒黑羊和巴音布魯克羊等優良綿羊品種均攜帶有FecB基因,其中湖羊攜帶的B 等位基因頻率高達0.83[3],產羔率能達到250%,其攜帶的多胎FecB 基因是決定湖羊個體多胎性狀的主要因素。
由于繁殖性狀的遺傳力普遍較低,因此,如果能確定與繁殖有關的主效基因,即可通過分子標記輔助選擇將其引入育種中,從而快速將優良基因型導入育種群體[4]。通過雜交等方式將FecB基因導入繁殖力較低的羊群,以期提高綿羊群體的繁殖性能,從而提高經濟效益,成為綿羊雜交改良的重要途徑之一。羅生金[5]用含有BB 型的湖羊公羊與當地的哈薩克母羊進行雜交,將FecB基因導入哈薩克羊中并進行檢測,初步培育出哈薩克羊多胎類型,建立了新的育種群。于振興等[6]從湖羊上提取FecB突變利用轉基因技術成功導入新疆細毛羊體內,為培育高繁殖率的新疆細毛羊奠定基礎,這些研究都為眾多地方綿羊品種的多胎類型(品系)選育工作提供了思路。
塔什庫爾干羊主要生活于帕米爾高原3000~5000m 的東坡,是區別于其他脂臀羊的高山放牧品種,具有適應性強、抗逆性強、適宜放牧、產肉性能好等特點。但由于受高寒干旱生態環境的限制,飼養管理粗放,特別是在秋冬季節飼草短缺,嚴重制約著塔什庫爾干羊繁殖性能的發揮,產羔率僅有105%左右。因此,建立塔什庫爾干羊多胎類型(品系),提高塔什庫爾干羊群體的繁殖性能,可以大幅提高養殖戶的經濟效益,增加農牧民收入。
綿羊的多胎類型(品系)選育離不開準確地對多胎性狀進行基因檢測。綿羊基因多態性檢測可分為限制性片段長度多態性(Restriction fragment length polymorphism,RFLP)、單鏈構象多態性(Single-strand conformation polymorphism,SSCP)及高分辨率熔解曲線(High resolution melting analysis,HRM)等幾種方法。但前兩種基因檢測技術普遍存在成本高、操作技術繁瑣,不能滿足綿羊生產中對FecB基因快速、低成本、高通量檢測的需求。HRM 可以通過目的基因片段PCR 擴增之后進行產物的熔解曲線分析來區分目的片段的微小序列差異,在基因突變、單核苷酸多態性(Single nucleotide polymorphisms,SNP)、甲基化和人類白細胞抗原(Human leukocyte antigen,HLA)配型等多個方面具有較大優勢[7]。因此,本試驗利用HRM 對塔什庫爾干羊基因組中FecB基因突變位點進行分析,建立快速、高靈敏度和特異性強的基因分型方法,為塔什庫爾干羊多胎新品系培育奠定基礎。
1 材料與方法
1.1 試驗動物
查閱塔什庫爾干縣各鄉鎮塔什庫爾干羊產羔記錄,篩選出塔什庫爾干羊產雙羔(多羔)母羊67 只,雙羔(多羔)羔羊136 只。用ACD 抗凝采血管頸靜脈采血3mL,-20℃凍存。
1.2 主要試劑及儀器
HRM 平臺熒光定量PCR 儀(型號為Light Cycler 480),購自羅氏公司;血液基因組提取試劑盒(上海生工);TaqDNA 聚合酶、10×PCR Buffer、Mgcl2(25mmol/L)、dNTP(10mmol/L)、Marker、6×DNA Loading Dye(上海生工);引物由上海生工公司提供。
1.3 DNA 的提取
采用常規酚-氯仿抽提法提取血樣中基因組DNA。利 用NanoDrop2000(Thermo Scientific,USA)測定提取DNA 樣品的完整性和濃度,將所有樣品的濃度調整為50ng/μL 左右。
1.4 PCR 擴增及產物檢測
PCR 擴增程序:95°C預變性3min;95℃30s,55℃60s,循環數為35 次。HRM 分析程序:95℃變性5min,40°C 冷卻2min;75~95℃收集熒光信號,每秒鐘收集25 次信號。利用LightCycler 480 自帶軟件分析數據。
1.5 不同長度引物篩選
根據FecB 基因的序列(GenBank 登錄號:AF357007.1),應用Oligo 軟件由上海生工公司分別合成2 對引物,將引物稀釋成10mmoL/L 的濃度-20℃保存。引物信息詳情見表1。
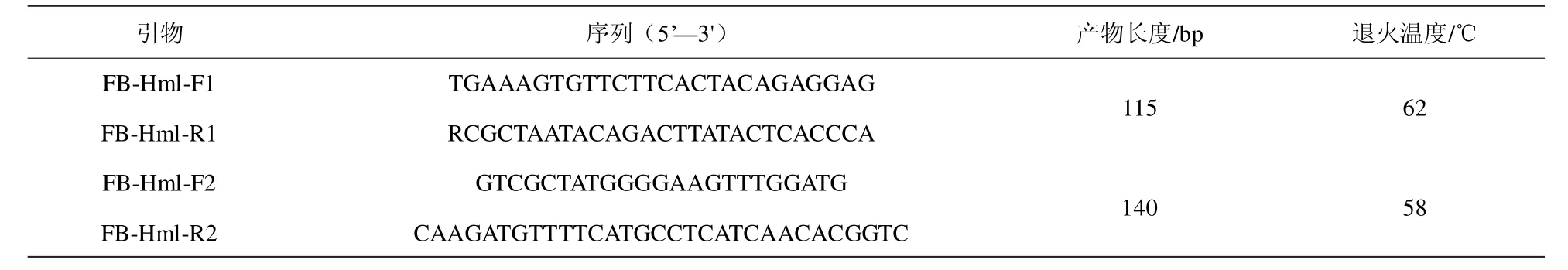
表1 擴增引物序列、產物長度及退火溫度
1.6 不同引物濃度的影響
對比了不同引物濃度下的HRM 分析,以確定最適合基因分型的引物濃度。引物稀釋后的工作濃度為10mmol/L,詳見表2。

表2 擴增引物濃度
1.7 不同鎂離子濃度的影響
鎂離子是影響PCR 的關鍵因素。通過對比不同濃度鎂離子,可以確定最適合的FecB 基因分型PCR 體系中Mg2+的濃度(表3)。

表3 鎂離子濃度
1.8 不同模板濃度的影響
在反應過程中不同的模板濃度與熒光染料的結合量不同,會導致信號強度差異變大,造成分型不準確。通過對不同模板濃度的影響進行HRM 分析,可以分析出最適合塔什庫爾干羊FecB 基因突變分型的模板濃度。本試驗采用的模板濃度分別為:500pg、5ng、50ng、500ng。
1.9 HRM 分析
每個優化的HRM 分析體系在反應過程中包括PCR 擴增階段和HRM 分析兩個階段,其中第一階段為40 個循環,第二階段為1 個循環。
2 結果
2.1 PCR 擴增片段的HRM 優化結果
根據前期試驗,本試驗設計了115bp、140bp 2 對不同片段大小的擴增引物,PCR 擴增之后進行HRM 分析。結果表明,115bp 擴增引物聚合性以及溫度的一致性較好,適合SNP 位點的分型。140bp 的效果較差,不適合塔什庫爾干羊FecB 基因的SNP 位點分型。
2.2 不同引物濃度對HRM 分析的影響
通過HRM 反應后分析,結果表明,在所設的梯度中添加量太大或太小時,擴增效率較差,熒光信號提前減弱,熔解溫度偏移。本試驗中引物添加量為0.6μL 時擴增效率較好,可以確定為較優添加量。
2.3 不同鎂離子濃度對HRM 分析的影響
通過HRM 反應后分析,結果表明,鎂離子濃度在低于1.6mmol/L 或高于4mmol/L 時,擴增反應達不到平臺期,擴增曲線散亂,熔解曲線中熒光信號下降。鎂離子濃度在2.4~3.2mmol/L時,擴增曲線和熔解曲線聚合性較好,熔解溫度差異分辨率清楚。
2.4 不同模板濃度對HRM 分析的影響
通過HRM 反應后分析,結果表明,模板濃度在設計范圍內時,擴增曲線均達到了平臺期。但是在500pg 以下或500ng 以上時,反應不穩定,效率較低。當反應模板濃度在50ng 時,擴增曲線、熔解曲線和熔解溫度差異曲線較符合塔什庫爾干羊FecB 基因分型的要求。
2.5 優化后的HRM 體系對塔什庫爾干羊FecB 基因SNP 位點分型結果
利用優化后的HRM 體系對塔什庫爾干羊多胎候選基因FecB進行分型分析,通過對高分辨率溶解數據進行歸一化和溫度平移處理得到的溶解曲線圖(圖1),結合溶解溫度差異顯示圖(圖2)可以清楚地區分出塔什庫爾干羊FecB基因不同堿基變異的差異。檢測結果表明,該基因多態性表現為一個單堿基位點突變,可以利用HRM體系實現FecB基因檢測的高通量、高準確率等實際需求。

圖1 熔解曲線圖
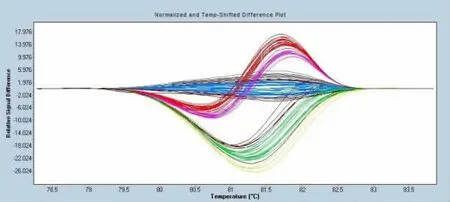
圖2 熔解溫度差異圖
2.6 塔什庫爾干羊FecB 基因檢測結果
由表4 可知,母羊和羔羊不攜帶純合型(BB),攜帶雜合型(B+)母羊占31.3%,羔羊占25.7%;攜帶野生型的母羊占68.7%,羔羊占74.3%。

表4 塔什庫爾干羊FecB 基因檢測結果
3 討論
綿羊的多胎基因FceB 被定位在第6 號染色體上,具有提高排卵和產羔的生物學作用[8],是目前已知的綿羊高繁殖力主效基因。一些綿羊品種如湖羊、小尾寒羊等攜帶的多胎FecB 基因是其產羔率能達到200%、250%的主要原因。在湖羊、小尾寒羊的雜交后代群體中,均能檢測到攜帶多胎FecB 基因的個體,其中,純合型(BB)母羊個體產羔率可達到250%,雜合型(B+)母羊個體產羔率可達到200%。因此,通過多胎基因檢測鑒定,可以區分純合型(BB)、雜合型(B+)以及野生型的母羊個體[9]。
有研究發現含有一個FecB基因B 等位基因的母羊(B+)排卵數增多1.50~1.65 個,產羔數增加0.9~1.2 個;含有兩個FecB 基因B 等位基因的母羊(BB)排卵數增多2.7~3.0 個,產羔數增加1.1~1.7 個;含有FecB 基因的母羊平均每胎產羔超過1 只,而野生型母羊產羔均為單胎[10],這說明FecB 基因確實能夠控制綿羊的多羔性狀。對杜泊羊與湖羊雜交后代羊羔的FecB 基因進行檢測分析后,發現隨著雜交代數增加,B 等位基因頻率逐漸降低,產羔率也隨之降低,且與基因型相關。
塔什庫爾干羊是經過長期封閉選育形成的適應高海拔、高寒條件的高原綿羊品種,但其繁殖率低,生產性能不高。由于塔什庫爾干羊存在FecB基因,但雙羔率較低,可以應用綿羊多胎基因檢測技術和精準選配技術,選擇純合型(BB)湖羊或小尾寒羊同塔什庫爾干羊雜交,在母羔出生后1月齡以內,就可采血鑒定多胎FecB基因,提前2 年完成對生產母羊個體的多胎性選留。也可以利用塔什庫爾干羊攜帶有FecB基因雜合型(B+)的特點,實現公、母羊多胎基因雜合型(B+)的精準選配,從而大量、穩定地生產攜帶有多胎FecB 基因的雜交后代,組建塔什庫爾干羊多胎類型核心群,提高群體產羔率30%~50%[10],可以有效提高塔什庫爾干羊養殖的生產效益。
飼養管理粗放、營養狀況不佳是塔什庫爾干羊雙胎率低的重要原因。因此,塔什庫爾干母羊在配種前要保證足夠的營養供給,維持良好的體況,以此來提高母羊受胎率。在塔什庫爾干羊母羊妊娠期和產羔后需要加強營養,以滿足母羊妊娠及哺乳雙羔的營養需要,提高羔羊成活率。
綿羊FecB基因分型多采用SSCP 和PCRRLFP 分析,HRM 是近些年興起的用于擴增子的特異性研究的新技術。利用HRM 和PCR-RLFP對綿羊多胎主效基因FecB基因進行檢測,具有準確、高效、自動化程度高等特點,可以在短時間內實現大規模群體多胎基因檢測[11]。引物的不同長度、不同濃度以及不同鎂離子濃度、不同模板濃度都會對HRM 分型產生影響。本研究中,最佳的HRM 分型條件為PCR 擴增片段在115bp、引物添加量為0.6μL、鎂離子濃度的最佳添加量在2.4~3.2mmol/L 和反應模板濃度50ng,這與蔣立斌等[11]研究相符,能夠準確地檢測FecB基因突變的不同類型,滿足高靈敏度、高通量、高準確性的要求。
4 結論
利用HRM 和PCR-RLFP 技術可以在塔什庫爾干羊群體中檢測到FecB基因,雖未發現純合型基因,但也可以利用FecB基因檢測技術對塔什庫爾干羊進行早期選種,縮短世代間隔,提高選擇強度,培育塔什庫爾干羊多胎類型。還可以選擇多胎性好的種公羊個體,通過多胎性狀雜交導入,提高塔什庫爾干羊群體中多胎FecB基因的比例,使群體的多羔率增加。此外,要加強母羊在配種與妊娠期間的營養和飼養管理,克服高寒高海拔生態環境的影響,提高受胎率和羔羊成活率,有效增加塔什庫爾干羊的繁殖性能。
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48
