分子標記輔助選擇小麥赤霉病抗病基因聚合體
2023-11-15 10:42:32陳天青李紅艷隋建樞羅永露吳文強何慶才
種子 2023年8期
陳天青, 李紅艷, 王 偉, 隋建樞, 羅永露, 吳文強, 程 斌, 何慶才
(1.貴州省農業科學院旱糧研究所, 貴陽 550006; 2.貴州醫科大學, 貴陽 550025)
條銹病(Stripe rust,SR)和赤霉病(Fusarium head blight,FHB)分別是由條形柄銹菌(Pucciniastriiformisf.sp.tritici)和禾谷鐮刀菌(Fusariumgraminearum)引起的小麥病害[1-2]。條銹病會破壞葉片的表皮組織,使植株葉面積指數減少,光合作用降低,導致小麥生長發育受阻、產量降低[3]。赤霉病主要為害穗部,影響籽粒儲藏物質的合成,籽粒被侵染后產生的真菌毒素會嚴重危害人畜安全[4]。貴州省因氣候潮濕多雨,一直是小麥多種病害(條銹病、白粉病、葉銹病)重發區,近年來小麥赤霉病由我國長江中下游冬麥區、華南冬麥區西擴,已上升為貴州省小麥的主要病害[5]。
抗病基因的發掘和抗病育種是防治小麥病害的有效方法。國內外眾多學者長期從事于抗病基因發掘工作。目前,國際上已正式命名了83個抗條銹病基因(Yr1~Yr83)[6],7個赤霉病抗病基因(Fhb1~Fhb7)[7-12]。
由于許多條銹病抗病基因具有小種專化性,隨著新的條銹菌強毒性小種條中34的出現,很多已知的條銹病抗病基因的抗性正在喪失[13]。貴協2號是貴州大學張慶勤教授通過野生二粒小麥與光稃野燕麥遠緣雜交、普通小麥回交等方法選育而成的小麥新品系,經田間成株期抗性鑒定對條銹病免疫至高抗水平,目前抗病基因雖不明確,但仍作為貴州主要的小麥條銹病抗源使用。
小麥赤霉病抗性是多基因控制的數量性狀,容易受環境的影響,目前已命名的主效基因中,Fhb1、Fhb2、Fhb4、Fhb5為報道較多、且較為穩定的主效基因,而Fhb1又是其中效應最大、抗性最穩定、并在國內外應用最廣泛的抗病基因[14]。目前,貴州省對赤霉病的研究僅限于材料的抗性鑒定和篩選,鮮有赤霉病抗性改良和聚合育種的報道。NMAS22是南京農業大學馬正強教授團隊以山東背景材料PH691為受體,赤霉病抗病品種望水白為供體,經雜交、不斷與PH691回交,分子標記輔助選育而成的新品系,對赤霉病表現為高抗,擺脫了望水白的不利農藝性狀,且攜帶有Fhb1、Fhb2、Fhb4、Fhb5等4個赤霉病抗病基因。
本研究將條銹病新抗源貴協2號和赤霉病新抗源NMAS22雜交,并構建了F2代臨時分離群體,采用田間鑒定與分子標記輔助選擇相結合的方法,篩選條銹病與赤霉病兼抗的植株,為新種質創制提供基礎材料。
1 材料與方法
1.1 試驗材料
小麥材料為貴協2號、NMAS22及以其構建的F2代臨時群體;禾谷鐮刀菌菌株(PH-1、F0609、F0980)由貴州省農業科學院植物保護研究所陳文博士惠贈。
1.2 試驗設計
將貴協2號和NMAS22雜交獲得的F2代的籽粒播種,試驗采用單行區,行長2 m,行距0.3 m,點播,每5 cm播種1粒,每行播種40粒,播種20行。在小區的四周種植條銹病誘發材料SY95-71。
1.3 小麥條銹病田間鑒定
采用自然誘發鑒定,在自然發病的情況下對F2代群體進行成株期調查,待SY95-71充分發病后,觀察記錄群體植株對條銹病的反應型。抗條銹性評價分級參考0~4級判定標準[15]。
1.4 小麥赤霉病田間鑒定
在小麥揚花初期,采用單花滴注法接種小麥赤霉病致病菌。將禾谷鐮刀菌菌株分生孢子液(105個/mL)滴入中選單株麥穗中部小穗的單側小花中。小麥黃熟前(接種后約21 d)調查接種穗的病小穗率和病害發生等級,計算平均嚴重度[16]。根據劃分標準劃分為免疫(I)、高抗(HR)、中抗(MR)、中感(MS)、高感(HS)[17]。
平均嚴重度(S)=∑(各病級穗數×相應病級數)/總調查總穗數。
1.5 分子標記檢測
取具有條銹病抗性植株的葉片裝入離心管,經液氮冷凍后用植物組織研磨儀研磨成粉末,CTAB法[18]提取DNA。采用與Fhb1、Fhb2、Fhb4、Fhb5連鎖的標記(表1)對中選植株進行赤霉病抗病基因分子檢測。PCR擴增采用10 μL體系,即2×TaqMaster Mix 5 μL,5 μmol/L正反向引物各0.2 μL,DNA模板(35 ng/μL) 2 μL,加ddH2O 2.6 μL補足10 μL。PCR擴增程序:94 ℃預變性5 min,94 ℃變性1 min,退火1 min(退火溫度根據引物設置),72 ℃延伸2 min,共35個循環,72 ℃終延伸10 min,4 ℃保存。

表1 抗赤霉病基因檢測標記信息
采用1.5%瓊脂糖凝膠或8%非變性聚丙烯酰胺凝膠電泳分析擴增產物。把F2代群體的帶型與親本NMAS22、貴協2號的帶型相比較,與NMAS22帶型相同記錄為R,與貴協2號帶型相同記錄為S,雜合型記錄為H,未有效擴增記為N。對于單標記檢測的基因(Fhb2和Fhb4),帶型為R、H的單株,認為其攜帶有抗病基因;對于雙標記檢測的抗病基因(Fhb1和Fhb5),兩個標記的帶型為R/R、H/H或R/H的單株,認為其攜帶有抗病基因,其中R/R為基因純合型,H/H和R/H為基因雜合型。
數據分析方法:一般統計分析采用Excel2013軟件進行,赤霉病抗病基因型與田間抗性的相關分析采用SPSS version20軟件進行。
2 結果與分析
2.1 小麥群體赤霉病與條銹病田間抗性表現
采用自然誘發法對732個NMAS22×貴協2號F2代單株進行條銹病抗性鑒定,結果顯示,在該群體中有349株表現為抗病(0~2級),有383株表現為感病(3~4級)。
采用單花滴注法完成了對F2代群體中685個單株的赤霉病抗性鑒定,結果表明,332個植株表現為高抗赤霉病,占群體總數的48.4%,135個植株表現為中抗赤霉病,占群體總數的19.6%,46個植株表現為中感赤霉病,占群體總數的6.7%,172個植株表現為高感赤霉病,占群體總數的25.1%。
2.2 赤霉病抗病基因的分子標記檢測及遺傳分析
用與抗赤霉病基因Fhb1、Fhb2、Fhb4、Fhb5相關的6對分子標記,對挑選出的具有條銹病抗性的植株樣本進行檢測,結果如圖1和表2所示。經對雙標記檢測基因的合并分析,在349份被檢測植株中,檢測到攜帶Fhb1、Fhb2、Fhb4、Fhb5的單株分別為201,245,248,194株,占被檢測植株的57.6%,70.2%,71.1%,55.6%,其中純合基因型植株分別為48,104,92,57株,占被檢測植株的13.8%,29.8%,26.4%,16.3%。
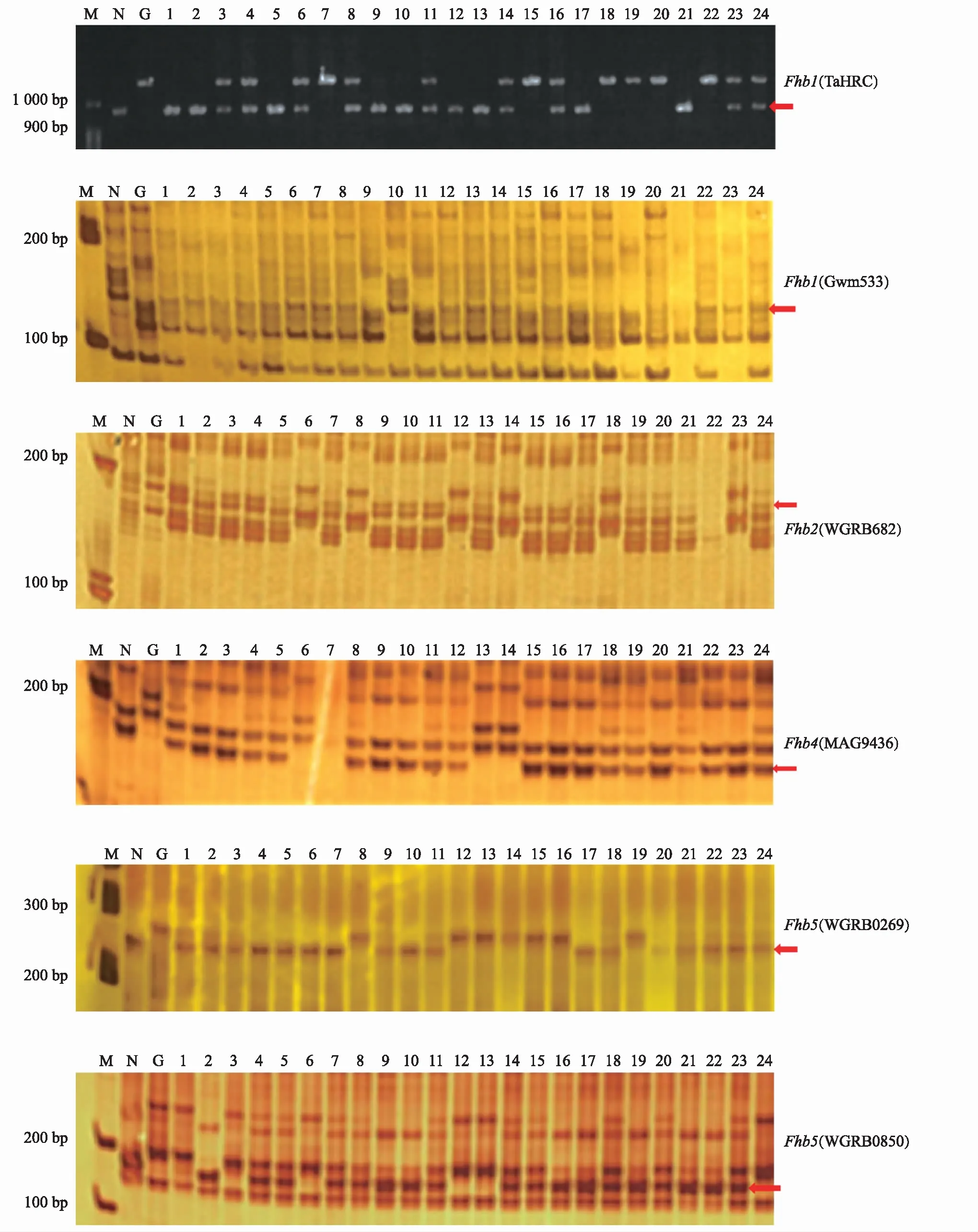
注:紅色箭頭指示抗病基因條帶;M為Marker;N為NMAS22;G為貴協2號;1~24號為F2代群體的部分單株。
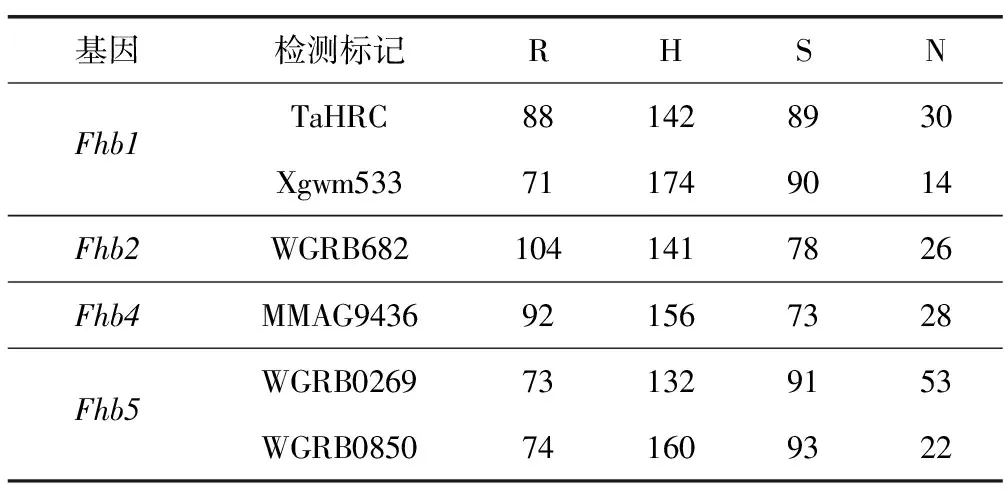
表2 抗赤霉病分子標記檢測結果
對以上標記不同基因型統計結果分別進行分離比的適合度測驗,結果(表3)顯示,Fhb1基因位點的TaHRC標記和Xgwm533標記、Fhb2基因位點的WGRB682標記、Fhb4基因位點的MAG9436標記、Fhb5基因位點的WGRB02659標記和WGRB0850標記的卡方值分別為3.85,2.66,5.57,2.50,5.65,2.36,均小于p=0.05水平的卡方值,F2代群體各單株基因型符合預期的1∶2∶1的分離比,表明其結果遵循孟德爾遺傳定律,即在F2代群體中對條銹病抗性單株的篩選不會引發Fhb1、Fhb2、Fhb4、Fhb5基因位點偏分離。
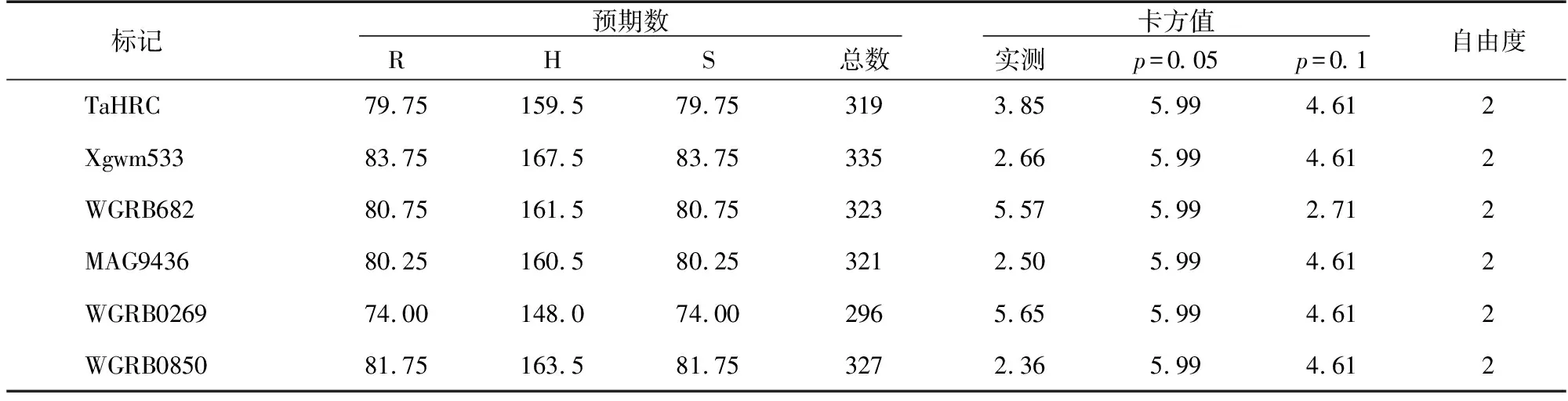
表3 卡方測驗分析結果
2.3 赤霉病抗病基因組合類型分析
由表4可見,三基因組合類型的單株最多,占被檢測單株的40.1%,其次為四基因組合和雙基因組合,分別占被檢測單株的24.4%和22.6%,而只攜帶1個抗病基因或不攜帶抗病基因的單株較少,僅占被檢測單株的10.6%和2.3%。
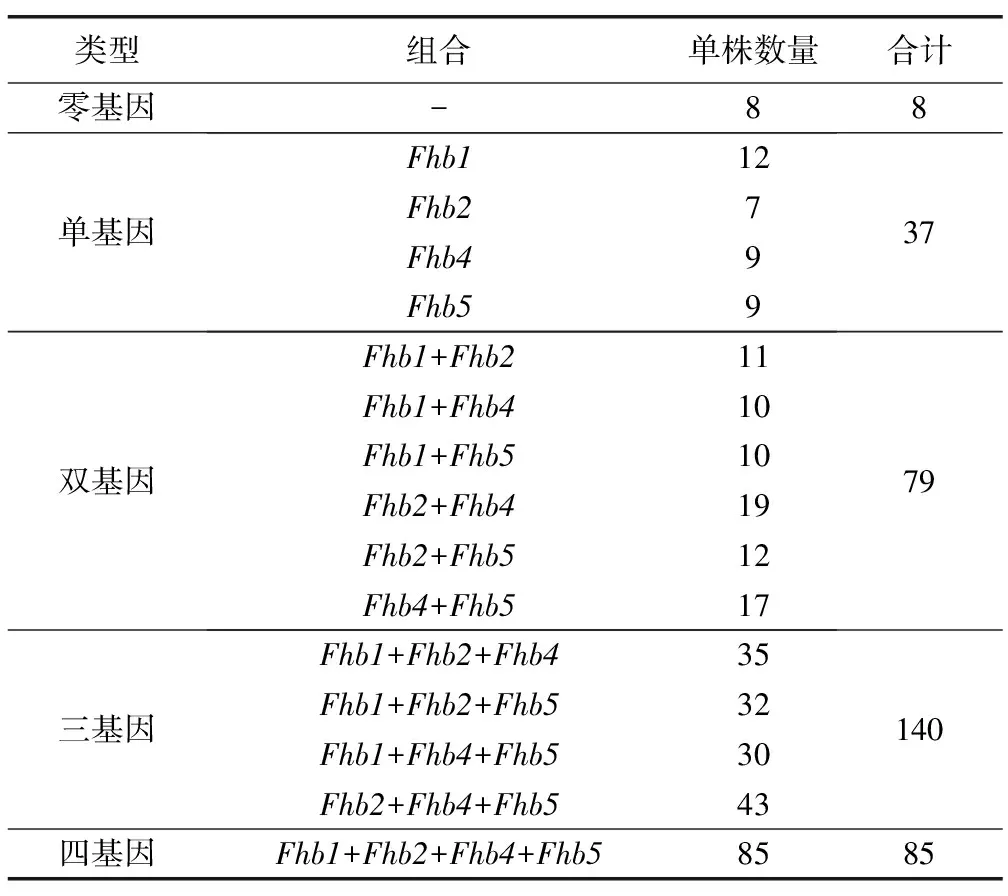
表4 赤霉病抗病基因組合分析
2.4 抗性相關性分析
將田間性狀鑒定結果與分子標記檢測結果做相關性分析,結果(表5)顯示,Fhb1基因位點的TaHRC標記和Xgwm533標記與病小穗率相關性為-0.261與-0.180,呈顯著負相關,Fhb2、Fhb4、Fhb5基因位點標記與病小穗率相關性均不顯著。
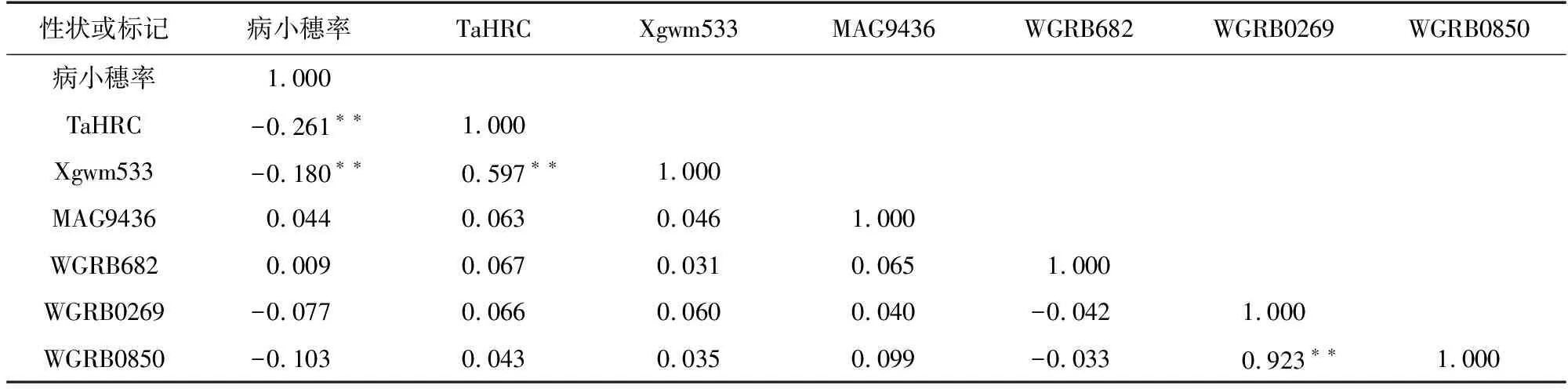
表5 相關性分析結果
2.5 四基因組合中純合基因數量分析
由表6可見,85份四基因組合型單株中,有25份攜帶1個純合的抗赤霉病基因,37份攜帶2個純合的抗赤霉病基因,20份攜帶3個純合的抗赤霉病基因,僅3份攜帶4個純合的抗赤霉病基因。

表6 四基因組合類型中純合基因數量
4 討 論
小麥赤霉病和小麥條銹病的危害逐年加重,除氣候和環境變化影響外,抗病品種缺乏也是重要原因之一。
小麥赤霉病抗性是由多基因控制的數量遺傳性狀,其抗性鑒定結果易受到環境影響,田間表型鑒定效果差,不利于育種后代的選擇[19-20]。要篩選抗赤霉病的育種材料,需要建立適宜的抗性鑒定技術和創造小麥發病有利條件,如氣候、溫度、濕度等。分子標記輔助選擇技術可以從基因型方面進行直接選擇,具有不受外界環境影響的優點,便于對表型鑒定困難、遺傳力低、隱性等性狀進行選擇,可使品種選育進程效率增加,目前已在多種作物的性狀改良上得到成功應用[21-23]。
目前,很多與赤霉病抗性有關的QTL被定位,但是只有少部分QTL位點能發揮較強的效果,如Fhb1,絕大多數的QTL位點效應比較小,容易受環境和遺傳背景的影響[20],阻礙了分子標記輔助選擇技術在赤霉病抗病育種上廣泛應用。目前,正式命名的7個抗病基因(Fhb1~Fhb7)中,Fhb1、Fhb2、Fhb4、Fhb5來自于普通小麥,分別被定位在普通小麥的3BS、6BS、4B、5A 染色體上[7-11]。江蘇溧陽小麥地方品種望水白高抗赤霉病,且同時攜帶了Fhb1、Fhb2、Fhb4、Fhb5,南京農業大學馬正強教授團隊已經開發了這4個基因位點的相關標記,并轉育了攜帶4個基因的育種材料NMAS22。
本研究利用條銹病新抗源貴協2號與NMAS22雜交,獲得F2分離群體,并對群體進行條銹病自然鑒定和赤霉病接種鑒定,獲得了349份具有條銹病抗性的植株、467份具有赤霉病抗性的植株。利用Fhb1、Fhb2、Fhb4、Fhb5等4個基因位點標記對349份條銹病抗病株進行檢測,最終篩選出85株攜帶4個抗赤霉病基因(基因型純合或雜合)的單株。
抗侵染、抗擴展均是小麥對赤霉病抗性的表現形式,本研究中的相關性分析結果顯示,Fhb1基因位點上的兩對標記均與病小穗率呈極顯著相關,而Fhb2、Fhb4、Fhb5這3個基因位點與病小穗率的相關性較低。Fhb1是公認的效應值高、抗性比較穩定、受遺傳背景影響較小的抗病基因,在本研究中得到了進一步的證明,可廣泛應用于分子標記輔助育種,而Fhb2對赤霉病抗擴展效應值低于Fhb1,在本研究中可能受到遺傳背景和環境的影響,表現為與赤霉病抗性相關性不大,故Fhb2不宜作為單一利用的基因應用于分子標記輔助育種。Fhb4和Fhb5在本研究中表現為與赤霉病抗性不相關。Fhb4和Fhb5是抗侵染類型的基因[24],本課題采用單花滴注接種法進行赤霉病抗性鑒定,單花滴注接種鑒定反映的是植株被侵染后的抗擴展能力,卻無法反映植株的抗侵染能力[19]。Fhb4和Fhb5在降低實際生產中小麥品種赤霉病發病率上具有重要意義,今后需同樣重視該類型基因的應用。本研究已篩選出在Fhb1、Fhb2、Fhb4、Fhb5等4個基因位點均攜帶赤霉病抗性基因的植株,為今后選育聚合4個抗病基因品種奠定了基礎,但這些植株中有些基因位點基因型為雜合型,后續研究需進一步對中選單株后代進行赤霉病抗病基因的追蹤與分析。
