水分添加量對高水分擠壓大豆粕植物蛋白肉品質及結構特性的影響
2023-11-20 07:45:02岳程程佟麗鳳邢維臣段玉敏王致力肖志剛
食品工業科技 2023年22期
關鍵詞:結構
岳程程,王 哲,佟麗鳳,邢維臣,段玉敏,王致力,肖志剛,王 鵬,
(1.沈陽師范大學,遼寧沈陽 110034;2.東方集團哈爾濱福肴食品有限公司,黑龍江哈爾濱 150001)
大豆是重要的油料作物和植物蛋白來源之一,在農業中占有重要的地位。低溫脫脂大豆粕(簡稱大豆粕)是大豆榨油后的剩余副產物,其含有35%~55%的蛋白質以及豐富的功能因子,如大豆異黃酮、大豆膳食纖維等[1],常作為飼料行業的加工原料。雖然大豆粕含有較豐富的植物蛋白質資源,但由于其經濟價值低,造成了優質植物蛋白資源的浪費,因此,有必要對大豆粕開展高值化開發利用。近年來,作為動物肉替代品的高水分擠壓植物蛋白肉受到了越來越多的關注[2-3],由于大豆分離蛋白中含有近90%的球蛋白和5%的白蛋白,使其成為加工植物蛋白肉的理想原料[4]。大豆分離蛋白主要是從大豆粕中分離提取的,其使用成本要遠高于大豆粕,同時,以大豆分離蛋白作為植物蛋白肉的加工原料也會直接推高產品的生產成本。此外,大豆粕中的部分不溶性膳食纖維在擠壓過程中會轉變為可溶性膳食纖維,這會影響擠壓產品的流變性能及品質[5],因此,探究利用大豆粕替代大豆分離蛋白作為高水分擠壓植物蛋白肉加工原料的可能性,對生產企業控制生產成本及提高大豆粕經濟價值都具有重要意義。
高水分擠壓技術是新興的植物蛋白重組技術[6],擠壓原料的水分含量通常達到60%以上,采用該技術制成的產品質地緊密、口感細膩,具有類似于動物肌肉的纖維組織[7],而且無需復水即可直接食用。由于水分具有增塑、導熱、降黏、熟化和利于成型的作用,因此,在擠壓過程中的水分添加量對植物蛋白肉的順利擠出和最終產品特性的形成有著十分重要的作用。產品原料被水浸潤軟化后,其熔融溫度低于熱分解溫度,易于后續的擠壓加工和營養成分的保護[8]。此外,水分還可以作為反應溶劑降低物料的黏度和在機筒內的停留時間,進而降低蛋白質亞基之間的聚合交聯,增加二硫鍵與氫鍵、疏水作用之間的協同作用,并影響產品的色澤,提高產品的纖維化程度[9]。Lin 等[10]以大豆分離蛋白和小麥粉為原料,通過研究發現物料含水率是影響擠壓過程最重要的因素。張玉潔等[11]以大豆分離蛋白(SPI)和金線魚魚糜為原料,證實了水熱參數組合作用顯著(P<0.05)影響SPI-魚糜混合物的微觀結構以及質構特性,并且物料水分的影響更顯著。
為了探究大豆粕能否作為高水分擠壓植物蛋白肉的加工原料,本文以低溫脫脂大豆粕為原料,固定進料速度、螺桿轉速、機筒溫度等條件,考察水分添加量對擠壓植物蛋白肉理化性質和微觀結構的影響。本文的研究結果將為大豆粕作為原料在高水分擠壓植物蛋白肉中的應用提供依據和參考。
1 材料與方法
1.1 材料與儀器
低溫脫脂大豆粕(水分含量10.62%;蛋白含量38.5%(干基);纖維含量4.33%(干基);灰分含量4.28%(干基))黑龍江冬雪生物科技有限公司;磷酸鹽博邁德生物化學試劑有限公司;尿素 天津市瑞金特化學品有限公司;巰基乙醇 麥克林試劑有限公司;十二烷基硫酸鈉 Coolaber Science &Technology。
UVTE-36 雙螺桿擠壓機 長沙創享食品科技有限公司;CT3 4500 型質構儀 美國Brookfield 公司;CR-400 型色彩色差計 日本柯尼卡美能達有限公司;TA Q20 差式掃描量熱儀 美國TA 公司;貝克曼冷凍離心機 美國貝克曼庫爾特公司;SU3500 掃描電子顯微鏡 日本Hitachi Ltd 公司。
1.2 實驗方法
1.2.1 大豆粕植物肉擠壓實驗 根據擠壓機的構造特點和預實驗結果,將水分添加量分別設置為10.5、11、11.5、12 和12.5 kg/h(通過預實驗測得上述水分添加量范圍內制備的植物肉含水量皆大于60%,所對應的含水量分別為61%、62%、63%、64%和65%),其他擠壓參數不變,將粉碎且過100 目篩的低溫脫脂大豆粕以8 kg/h 的速度從喂料區加入到雙螺桿擠壓機機筒中,設定螺桿轉速為220 r/min,機筒共開啟六段加熱區,溫度分別設定為(40±1)、(60±1)、(90±1)、(120±1)、(150±1)和(160±1)℃,冷卻裝置溫度設定在(55±1)℃,進行擠壓試驗。將擠出的植物蛋白肉冷卻至室溫后,對一部分試樣進行質構和顏色測試,另一部分試樣經真空冷凍干燥后粉碎,并過100 目篩后進行后續分析。
1.2.2 豆粕植物肉色澤測定 采用色差儀測定植物蛋白肉的L*、a*、b*的數值。對每個樣品3 個不同位置的色澤進行測定,每個實驗測定至少進行3 次,去掉異常值,取平均值。
1.2.3 豆粕植物肉質構特性測定 參考肖志剛等[12]的方法并做適當修改。將植物蛋白肉樣品裁剪為長寬均為25 mm 高為10 mm 的長方體,設置質構儀操作條件:TPA 模式,探頭選用TA-5,測試前速度1.0 mm/s,測試速度1.0 mm/s,測試后速度1.0 mm/s,下壓程度50%,測定樣品硬度、彈性、咀嚼性的數值,每個樣品進行3 次平行測定,取平均值。
1.2.4 豆粕植物肉持水性、持油性測定
1.2.4.1 豆粕植物肉持水性 參考Mazaheri 等[13]的方法并加以改進。稱取1.5 g 植物蛋白肉樣品放入到50 mL 的離心管中,然后加入30 mL 的去離子水,室溫下渦旋10 min 后于4000 r/min 離心20 min,然后倒掉上清液,植物蛋白肉的持水性根據下列公式進行計算:
式中:m 為樣品的質量,g;m1為樣品的質量+離心管質量,g;m2為離心管除去上清液后的質量,g。
1.2.4.2 豆粕植物肉持油性 參考Mazaheri 等[13]的方法并加以改進。稱取1.5 g 植物蛋白肉樣品放入到50 mL 的離心管中,然加入30 mL 的葵花籽油,室溫下渦旋10 min 后于4000 r/min 離心20 min,然后倒掉上清液,植物蛋白肉的持油性根據下列公式進行計算:
式中:m 為樣品的質量,g;m1為樣品的質量+離心管質量,g;m2為離心管除去上清液后的質量,g。
1.2.5 豆粕植物肉熱特性分析 參考張金闖[14]的方法并加以改進。稱取5 mg 樣品置于耐高壓鋁盤中,密封。使用TA Q20 差式掃描量熱儀,掃描溫度區間25~130 ℃、升溫速率5 ℃/min,N2保護,N2流速為50 mL/min。空鋁盤做參比。記錄熱轉變起始溫度(To)、熱轉變峰值溫度(TP)、熱轉變焓值(ΔH)和峰寬(ΔT),每個樣品重復測定3 次,結果取平均值。
1.2.6 豆粕植物肉SDS-PAGE 凝膠電泳測定 利用SDS-PAGE 凝膠電泳測定植物蛋白肉的相對分子量,具體操作參考張金闖[14]的方法并加以改進。將不同水分添加量擠出樣品經過冷凍干燥后,粉碎,過100 目篩網。將過篩樣品(2 mg)溶于0.5 mL 樣品緩沖液(0.08 mol/L Tris-HCl 緩沖液,pH6.8),1%(w/v)SDS,2%(v/v)2-ME,5%(v/v)甘油和0.025%(w/v)溴酚藍并充分混合。然后將樣品在沸水浴中加熱10 min,12000 r/min 離心10 min。進樣時取上清5 μL進樣,Marker 分子量范圍為10000 至250000 Da。在Mini-PROTEAN 系統上進行電泳,所配制電泳膠為5%的濃縮膠和12%的分離膠。電泳選取恒壓模式,首先電壓選用80 V,溴酚藍條帶跑至濃縮膠底端時換用110 V 電壓,等溴酚藍條帶跑至底端停止。采用考馬斯亮藍R250 振蕩染色60 min。染色完成后,用脫色液進行振蕩脫色10 h,每隔1 h 更換一次脫色液。使用Gel DocTMXR+凝膠成像系統拍攝蛋白質電泳光譜的圖像。
1.2.7 豆粕植物肉掃描電鏡觀察 將冷凍干燥后的植物蛋白肉樣品進行橫縱方向切片并固定在樣品臺上。采用Hitachi IB-5 離子鍍膜儀進行噴金處理后,置于SU8010 掃描電子顯微鏡下放大300 倍觀察并拍照。
1.2.8 豆粕植物肉化學交聯作用測定 參考肖志剛等[15]的方法并加以改進。取粉末狀樣品0.25 g(精確至0.1 mg)于25 mL 燒杯中,分別加入10 mL 八種溶劑:a.0.035 mol·L-1pH7.6 磷酸鹽緩沖溶液(P);b.磷酸鹽緩沖液中尿素含量為8 mol·L-1尿素(P+U);c.磷酸鹽緩沖溶液中2-巰基乙醇(2-ME)0.1 mol·L-1(P+M);d.磷酸緩沖液中十二烷基硫酸鈉(SDS)1.5 g·100 mL-1(P+S);e.磷酸鹽緩沖溶液中尿素和2-ME 分別為8 和0.1 mol·L-1(P+U+M);f.磷酸鹽緩沖液中SDS 和尿素分別為1.5 g·100 mL-1和8 mol·L-1(P+U+S);g.磷酸鹽緩沖液中SDS 和2-ME 分別為1.5 g·100 mL-1和0.1 mol·L-1(P+S+M);h.磷酸鹽緩沖溶液中尿素為8 mol·L-1,SDS 為1.5 g·100 mL-1,2-ME 為0.1 mol·L-1(P+U+S+M),常溫下攪拌浸提1 h 后放入冰箱4 ℃保存,離心(12000×g,10 ℃,15 min),取上清液。沉淀中再分別加入10 mL 上述八種溶劑,重復浸提兩次,離心。取1 mL 上清液于1.5 mL 離心管中(36×3 個),混勻(搖床160 r/min,1 h)。加20 μL 到96 孔酶標板的樣品孔中,采用Lowery法測得(測定波長650 nm)上清液中可溶性蛋白含量,原料和擠出樣品中總的蛋白質含量采用凱氏定氮法測定,蛋白質溶解度計算方法為上清液中可溶性蛋白與樣品中總蛋白的比值。每次測量重復三次。蛋白質溶解度,總蛋白質量濃度根據下列公式進行計算:
根據不同溶液中的溶解度計算各種化學鍵:天然狀態蛋白:a;氫鍵作用:b-a;二硫鍵:c-a;疏水作用:d-a;氫鍵和二硫鍵交叉作用:e+a-b-c;氫鍵和疏水作用交叉作用:f+a-b-d;疏水和二硫鍵交叉作用:g+a-c-d;氫鍵和二硫鍵和疏水作用交叉作用:h+b+c+d-a-e-f-g。
1.2.9 豆粕植物肉蛋白質二級結構分析 參考Guo等[16]的方法并加以改進,將1 mg 樣品和100 mg 溴化鉀稱重制成透明片劑,掃描范圍設置為1600~1700 cm-1,分辨率為4 cm-1和信號掃描64 次。使用Peakfit 版本4.12 軟件處理光譜。
1.3 數據處理
數據結果分析采用SPSS 20.0 統計軟件和Origin 2018 軟件進行分析及圖形繪制。通過單因素方差分析(ANOVA)確定顯著性,然后進行Duncan 檢驗,通過Pearson 相關系數法對實驗數據進行相關性分析,所得試驗結果均采用三次重復試驗的平均值和標準差表示。
2 結果與分析
2.1 水分添加量對植物蛋白肉外觀色澤的影響
水分添加量對植物蛋白肉外觀色澤的影響如表1 所示,隨著水分添加量從10.5 kg/h 增加到11.5 kg/h時,植物蛋白肉亮度值(L*)逐漸增加,而紅綠值(a*)逐漸下降,這表明適當增加水分含量有利于提高植物蛋白肉產品的外觀色澤。水分添加量的增加促進了組織化蛋白形成更多的定向纖維和多孔結構,并且隨著纖維化程度的提高,在植物蛋白肉的表面形成了一層光滑的薄膜結構[8];同時水分含量增加也會導致物料粘度降低,這減少了推動物料通過機筒所需的力以及物料與螺桿和機筒之間的摩擦,縮短了物料在擠壓機中的停留時間,降低了美拉德反應引起的褐變程度[17],從而提高了植物蛋白肉的光澤度。此外,隨著水分含量的增加,水分子與受熱斷開的蛋白質鏈上的親水結合位點的結合更加緊密,塑化作用增強,這可能與顏色L*的增加有關。陳鋒亮等[18]和張汆[19]也發現了物料水分的增加會促使大豆蛋白素肉或花生蛋白素肉變得更亮白。然而,當水分添加量從11.5 kg/h進一步升高到12.5 kg/h 時,植物蛋白肉的亮度值呈下降趨勢,這可能是因為當水分添加量過多時,物料的黏度也隨之升高,物料在機筒內的滯留時間變長,因此加劇了物料之間發生美拉德反應的程度[20],從而使植物肉的外觀色澤下降。實驗結果表明,適量的物料水分含量有助于提高植物蛋白肉產品的外觀色澤。
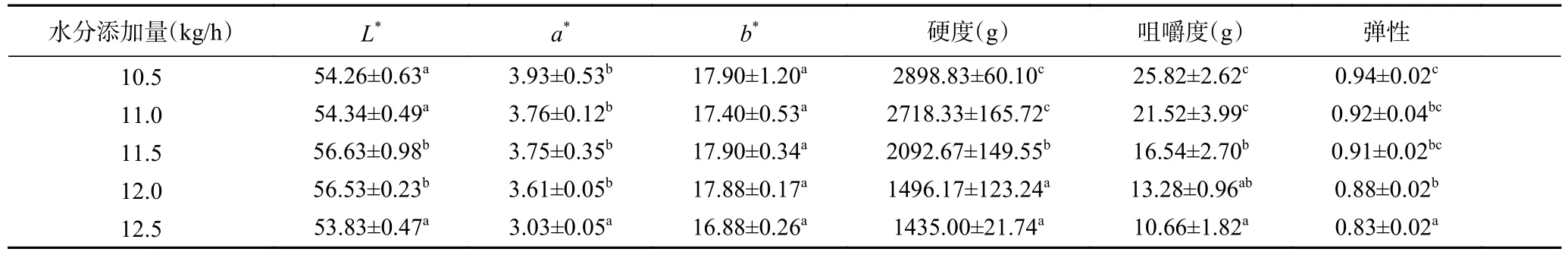
表1 水分添加量對植物蛋白肉外觀色澤、質構特性的影響Table 1 Effect of water addition on appearance and color and texture properties of plant protein meat
2.2 水分添加量對植物蛋白肉質構特性的影響
表1 列出了水分添加量對植物蛋白肉質構特性的影響結果。從表1 中可以看出,不同的水分添加量對植物蛋白肉的硬度、咀嚼度和彈性具有顯著影響(P<0.05)。隨著水分添加量的逐漸增加,物料在機筒內結合了過多的水分,導致植物蛋白肉的質地變得愈加柔軟,其硬度、咀嚼度和彈性均顯著降低(P<0.05),尤其以硬度變化最為明顯,植物蛋白肉的硬度由2898.83 g 顯著降至1435.00 g(P<0.05),而咀嚼度和彈性則分別由25.82 g 和0.94 降至10.66 g和0.83,由此可知,通過調節水分添加量能夠調控植物蛋白肉產品的口感。
2.3 水分添加量對植物蛋白肉持水性、持油性的影響
水分添加量對植物蛋白肉持水性、持油性的影響如圖1 所示,隨著水分添加量的增加,植物蛋白肉的持水性、持油性均呈現出先升高后降低的趨勢。當水分添加量低于11.5 kg/h 時,植物蛋白肉持水性和持油性的升高主要是因為水分在擠壓過程中起到了增塑的作用[21],一定量的水分可以加快聚合物分子的運動,促進蛋白質-蛋白質及蛋白質-水的相互作用,從而促使更多的疏水基團暴露,這加速了蛋白質的變性,并引起蛋白質的凝聚度下降,蛋白質的溶脹特性和形成網絡結構的能力進一步得到增強,產品組織化程度下降,從而導致植物蛋白肉的持水和持油能力均得到提高[22]。此外,豆粕中的不溶性膳食纖維在擠壓的作用下會轉變為可溶性膳食纖維,這也有助于提高植物蛋白肉的持水能力[23]。水分添加量由12.0 kg/h增加至12.5 kg/h 時,植物蛋白肉的持水性和持油性均呈下降趨勢,這可能是由于過量的水分阻礙了蛋白質與蛋白質之間的相互作用,導致形成了較弱的凝膠網絡[24],從而造成植物蛋白肉持水性和持油性的下降。可以看出,適當提高水分添加量有利于增強植物蛋白肉的持水和持油能力。

圖1 水分添加量對植物蛋白肉持水性、持油性的影響Fig.1 Effects of water addition on water absorption capacity,oil absorption capacity of plant protein meat
2.4 水分添加量對植物蛋白肉熱特性的影響
水分添加量對植物蛋白肉熱特性的影響如表2所示,由于豆粕中大豆蛋白11S 亞基的熱轉變焓值明顯高于7S 亞基,其熱轉變峰更明顯,因此,在本研究中,植物蛋白肉的熱特性變化僅以11S 亞基為代表進行分析[18]。當水分添加量由10.5 kg/h 增加到11.0 kg/h 時,植物蛋白肉發生熱轉變的起始溫度(To)和峰值溫度(Tp)均呈現出上升的趨勢,而峰寬(ΔT)和熱轉變焓值(ΔH)卻出現明顯下降,這可能與水分含量的突然增大有關。物料水分含量的突然增大會使蛋白分子結構變得靈活[25],分子結構更易于被打開[26],從而降低了植物蛋白肉的熱穩定性。當水分添加量由11.0 kg/h 繼續升高到12.5 kg/h 時,植物蛋白肉發生熱轉變的起始溫度(To)和峰值溫度(Tp)隨之逐漸下降,但峰寬(ΔT)和熱轉變焓值(ΔH)卻逐漸升高,To和Tp分別從78.84 和101.21 ℃降低至67.23和96.31 ℃,ΔT 和ΔH 分別從34.15 ℃和26.75 J/g升高至58.71 ℃和57.88 J/g,這表明水分含量進一步的升高加快了植物蛋白肉中大豆蛋白的熱轉變速率。而植物蛋白肉中大豆蛋白的熱焓值△H 顯著增大,說明其分子結構更加穩固,打開其分子結構所需要的能量明顯增加。

表2 水分添加量對植物蛋白肉熱特性的影響Table 2 Effect of water addition on thermal properties of plant protein meat
2.5 水分添加量對植物蛋白肉相對分子量的影響
豆粕中的大豆蛋白主要組分包括7S 亞基(分子量分別為78.41、73.00、54.05 kDa)和11S 亞基(分子量分別為42.18、37.62、34.03、31.58、22.05、19.02 kDa)[18]。從圖2 的SDS-PAGE 電泳結果中可以看出,隨著擠壓過程中水分添加量的升高,植物蛋白肉樣品中的7S/11S 球蛋白比例也發生了顯著的變化。當水分添加量為10.5 kg/h 時,植物蛋白肉樣品電泳條帶中亞基的顏色與條帶頂端顏色均很淺。隨著水分添加量的進一步增加,7S 中的α′亞基(78.41 kDa)、α亞基(73.00 kDa)、β亞基(54.05 kDa)以及11S 中的A 組分(42.18 kDa)和B 組分(22.05 kDa)的亞基條帶逐漸變寬,并且亞基條帶顏色以及擠出樣品條帶頂端的顏色逐漸變深,這可能是因為隨著水分添加量的增加,豆粕中的大豆蛋白經擠壓蒸煮后發生聚合、交聯等變性反應的程度增強,形成了分子量較大的聚合物[27]。然而,當水分添加量由12 kg/h 增加到12.5 kg/h 時,植物蛋白肉樣品條帶頂端的顏色略微變淺,這可能是因為形成的較大分子量的聚合物可以透過濃縮膠,但卻不能透過分離膠的網絡孔隙,因此在條帶的頂端發生了聚集[28]。可以看出,適當提高水分添加量有利于提高蛋白質聚合交聯變性的程度。

圖2 水分添加量對植物蛋白肉相對分子量的影響Fig.2 Effect of water addition on relative molecular weight of plant protein meat
2.6 水分添加量對植物蛋白肉微觀形貌的影響
掃描電子顯微鏡觀察植物蛋白肉的形態如圖3所示。在擠壓過程中,由于豆粕中含有的纖維成分與蛋白質流動性能不同,在高溫、高剪切條件下兩者發生相分離,從而在冷卻裝置中逐漸形成具有拉絲效果的纖維結構[29]。當水分添加量為10.5 kg/h 時,植物蛋白肉拉絲效果并不明顯,所形成的纖維結構內部孔洞較大,出現了明顯分層的現象,這主要是因為在相對較低的水分含量下,植物蛋白肉在冷卻模具中受到較高摩擦力和剪切力所導致[30]。隨著水分添加量增加至11.5 kg/h 時,植物蛋白肉表面形成了更加致密的纖維細絲結構,組織蛋白層狀結構清晰且整齊,具有精細的定向結構,這主要是由于合適的水分含量促進疏水基團暴露[31],加速了蛋白質的變性[32],使豆粕中的蛋白質分子由最初的無序多孔結構變為結構致密且具有一定規則的晶型排列結構,這有利于三維網絡結構的形成,從而使植物蛋白肉形成更多具有類似于肉類纖維的條狀纖維結構。然而隨著水分添加量繼續升高至12.5 kg/h 時,植物蛋白肉形成了更多層狀而非纖維狀的結構且疏松多孔,這主要是因為隨著水分添加量的升高,機筒的高溫導致更多的水蒸汽產生,蛋白質形成了更多的膜狀氣孔結構[9],這加劇了蛋白質內部疏水基團的暴露和蛋白質交聯變性程度的增大,造成產品氣孔結構更加明顯。此外,過量的水分促使豆粕中更多的糖類物質溶于水中,加劇了蛋白質的溶脹程度[28],使得蛋白質-蛋白質相互作用減弱,產生了不相容的聚合物網絡[33],從而導致植物蛋白肉形成了更多分離的層狀結構。
2.7 水分添加量對植物蛋白肉化學交聯作用力的影響
水分添加量對植物蛋白肉化學交聯作用力的影響如表3 所示,植物蛋白肉的結構是由疏水作用、氫鍵、二硫鍵以及彼此之間的交互作用共同維持的。從表3 可以看出,隨著水分添加量的增加,植物蛋白肉中的氫鍵數量和疏水作用均隨之升高,這表明提高水分含量能夠促進蛋白質與水混合,從而形成更多的氫鍵[16]。當水分添加量由10.5 kg/h 增加到11 kg/h時,植物蛋白肉中二硫鍵顯著地增加(P<0.05),而當水分添加量進一步增加時二硫鍵數量則無顯著變化。Sheard 等[34]認為當物料水分含量較高時,疏水作用是維持蛋白聚集的主要作用力;而隨著水分含量的降低,二硫鍵逐漸形成并成為穩定結構的主要作用力。此外,當水分添加量由10.5 kg/h 增加到12.5 kg/h時,氫鍵、疏水相互作用、二硫鍵三者交叉作用呈現先升高后下降的趨勢,且在水分添加量為11.5 kg/h時,氫鍵、疏水相互作用、二硫鍵三者交叉作用達到最強,由此可知,適當地提高水分添加量有利于蛋白質變性,對氫鍵、疏水相互作用、二硫鍵三者交互作用具有促進作用。

表3 水分添加量對植物蛋白肉化學交聯變化的影響Table 3 Effect of water addition on the chemical cross-linking force of plant protein meat
綜合上述結果可知,植物蛋白肉的結構由共價鍵與非共價鍵的交互作用共同維持。在所考察的水分添加量范圍內,共價鍵與非共價鍵的交互作用對維持植物蛋白肉結構的貢獻最大,非共價鍵作用和共價鍵作用次之,并且非共價鍵作用大于共價鍵作用,這與張金闖[14]的研究結果一致。
2.8 水分添加量對植物蛋白肉二級結構的影響
水分添加量對植物蛋白肉紅外光譜圖的影響如圖4a 所示。在3700~3100 cm-1內,5 種水分添加量的植物蛋白肉均出現較寬的特征條帶,這主要由O-H、N-H 伸縮振動和氫鍵伸縮振動引起的,在3000~2700 cm-1處出現的尖銳吸收峰,這歸因于C-H 收縮振動[35]。植物蛋白肉在2500~2300 cm-1處出現的吸收峰主要與O=C=O 和R-N=C=O 伸縮振動有關[36],此處吸收峰強度隨著水分含量的增加先增強后減弱,并且在水分添加量為11.0 kg/h 時達到最強,這表明植物蛋白肉的穩定性隨著水分添加量的升高呈先升高后降低的趨勢,這與前面熱特性分析結果一致。而在1500~1700 cm-1處的吸收峰與美拉德反應產物如羰基化合物、希夫堿和吡嗪類相關的C=O、C=N 和C-N 伸縮振動有關[37],吸收峰強度均隨著水分含量增加而減弱,表明擠壓過程中水分含量對美拉德反應的發生程度具有一定影響。

圖4 水分添加量對植物蛋白肉二級結構的影響Fig.4 Effect of water addition on the secondary structure of plant protein meat
由圖4a 可知,隨著水分添加量的變化并未有新峰出現,但結合傅里葉自卷積、二階導數和高斯曲線擬合(圖4b~圖4f),通過定量分析頻譜的二階導數發現蛋白質二級結構隨著水分添加量的增加發生顯著變化。根據文獻[16]報道可知,α螺旋由分子內氫鍵維持,位于1646~1662 cm-1;β轉角為松散有序結構,位于1662~1681 cm-1;無規則卷曲結構由C=O和H2O 之間形成的氫鍵維持,位于1637~1645 cm-1;β1 為反向平行式折疊的β層狀結構,由分子間氫鍵維持,位于1608~1622 和1682~1700 cm-1。β2 為平行式折疊的β層狀結構,由分子內的氫鍵維持,位于1622~1639 cm-1。
水分添加量對植物蛋白肉二級結構的影響如表4 所示。由表4 與圖4b~圖4f 可知,隨著水分添加量的增加,植物蛋白肉的二級結構分別出現在不同位置,并且具有不同峰面積。β折疊為植物蛋白肉二級結構的主要作用力,其中α螺旋與β2 所占含量高于β1,表明分子內氫鍵作用力更強,這與Guo 等[16]研究的結果一致。隨著水分添加量由10.5 kg/h 增加到11.5 kg/h 時,β1、β2 與α螺旋含量均呈降低趨勢,這表明植物蛋白肉中維持β1、β2 與α螺旋結構的氫鍵作用力減弱,這可能與水分含量的突然增大有關。結合DSC 與化學交聯結果可知,物料水分添加量的突然增加會使蛋白分子結構變得靈活,其分子結構更易于被打開[26],從而使植物蛋白肉的熱穩定性降低。此外,當水分添加量由10.5 kg/h 增加到11.5 kg/h時,β轉角和無規則卷曲分別呈現出先升高后降低與先降低后升高趨勢,這表明在適當的擠壓水分條件下,分子二級結構中松散有序結構逐漸增加,這有利于β折疊、α螺旋、無規則卷曲結構向β轉角的轉化。當繼續增大水分含量時,加劇了蛋白質內部疏水基團的暴露和蛋白質交聯變性程度的增大,蛋白質形成了更多的膜狀氣孔結構,這與掃描電鏡觀察到的結果一致。

表4 水分添加量對植物蛋白肉二級結構的影響Table 4 Effect of water addition on the secondary structure of plant protein meat
2.9 植物蛋白素肉的相關性分析
對不同水分添加量植物蛋白肉的質構特性、色差、二級結構、變性熱焓、化學交聯鍵進行相關性分析,結果如圖5 和表5 所示。從圖5 可知,12.5 和12.0 kg/h 水分添加量與10.5、11.0、11.5 kg/h 水分添加量方向不同,并且11.5 kg/h 水分添加量與其他4 組所在象限均不同,表明水分含量對植物蛋白肉作用效果顯著。同時從載荷可以看出,硬度與咀嚼度這兩個特性彼此接近,表明它們之間存在正相關關系。此外,二硫鍵所處正交方向,表明其獨立變化,對其他性能沒有影響。由表5 可知硬度、咀嚼度、彈性指數與氫鍵、疏水相互作用、二硫鍵和疏水相互作用均呈負相關,而咀嚼度、彈性指數與α螺旋含量呈正相關。P<0.01 時,L*與β轉角含量呈正相關,L*與氫鍵+二硫鍵呈負相關且皮爾遜系數最大。P<0.01 時,持水性與β1 呈負相關,P<0.05 時,持水性與β轉角和氫鍵+二硫鍵+疏水相互作用呈正相關。持油性在P<0.01 時與疏水相互作用呈正相關且皮爾遜系數最大為0.743。

表5 植物蛋白肉的相關性分析Table 5 Correlation analysis of soybean meal simulated meat

圖5 植物蛋白肉的相關性分析載荷圖、分值圖及雙標圖Fig.5 Loading plot and scora plot and biplot correlation analysis of soybean meal simulated meat
綜上所述,植物蛋白肉質構特性主要是化學交聯力中的氫鍵、二硫鍵、氫鍵+疏水相互作用及二級結構中α螺旋共同作用的結果。二級結構中β1、β2 和β轉角的變化對植物蛋白肉的亮度影響顯著。此外植物蛋白肉持水性是β1、β轉角、無規則卷曲、氫鍵+二硫鍵+疏水相互作用的結果,而持油性變化主要受疏水相互作用影響。
3 結論
本文以低溫脫脂大豆粕為原料,探究了水分添加量對高水分擠壓植物蛋白肉理化性質和微觀結構的調控作用。研究結果表明,水分添加量作為擠壓過程中的重要參數對植物蛋白肉的品質特性影響效果顯著,隨著擠壓過程中水分添加量的增加,植物蛋白肉的硬度、彈性、咀嚼性均顯著下降,而亮度值、持水性和持油性均呈先升高后降低趨勢。水分添加量在11.5 kg/h 時,植物蛋白肉具有較好的品質特性與纖維結構。適當提高水分添加量有利于α-螺旋、β-折疊、無規則卷曲結構向β-轉角的轉化,同時促進共價鍵與非共價鍵的交互作用,對植物蛋白肉松散有序結構的增加,形成具有一定規則的晶型排列結構具有促進作用。高水分擠壓植物肉具有更接近于動物肉的纖維結構和質構等特性,對其開發在減少全球碳排放、滿足民眾日益增加的肉類需求、減少動物性食品的消費以及確保中國和世界食物供給安全等方面具有重要意義。
猜你喜歡
小獼猴智力畫刊(2023年4期)2023-04-23 08:49:58
哲學評論(2021年2期)2021-08-22 01:53:34
中華詩詞(2019年7期)2019-11-25 01:43:04
模具制造(2019年3期)2019-06-06 02:10:54
中學生數理化·高一版(2018年1期)2018-02-10 05:20:03
影視與戲劇評論(2016年0期)2016-11-23 05:26:01
七彩語文·寫字與書法(2016年7期)2016-07-28 21:40:22
七彩語文·寫字與書法(2016年6期)2016-07-15 19:36:34
人間(2015年21期)2015-03-11 15:23:21
現代企業(2015年9期)2015-02-28 18:56:50
