基于線粒體Cyt b基因的洪澤湖翹嘴鲌群體遺傳多樣性分析
2023-11-21 08:14:30李大命楊子萍劉燕山谷先坤唐晟凱楊家新
水產科技情報 2023年6期
李大命 楊子萍, 劉燕山 谷先坤 唐晟凱 楊家新
(1 江蘇省淡水水產研究所,江蘇省內陸水域漁業資源重點實驗室,江蘇南京 210017;2 南京師范大學海洋科學與工程學院,江蘇南京 210023)
洪澤湖(118°10′~118°52′E,33°06′~33°40′N)位于江蘇省西部淮河下游,水域面積約1 597 km2,是我國五大淡水湖之一。洪澤湖具有相對穩定的水位、寬廣的水域和豐富的水生生物資源,為漁業發展提供了良好條件,是我國主要淡水漁業基地之一[1]。近幾十年來,由于過度捕撈、水體環境質量下降、水利工程建設等多重因素影響,洪澤湖魚類物種數量減少,群落結構趨于單一化,魚類小型化趨勢嚴重,大型肉食性魚類如翹嘴鲌等資源衰退嚴重[1-2]。
翹嘴鲌(Culteralburnus)隸屬于鯉形目(Cypriniformes)、鯉科(Cyprinidae)、鲌亞科(Culterinae)、鲌屬(Culter),俗稱白魚、翹嘴白絲、大白魚等,廣泛分布于我國各個水系[3]。翹嘴鲌為兇猛肉食性魚類,是水域生態系統中的頂級消費者,能有效控制小型魚類種群數量,減輕小型魚類對浮游生物的壓力,在穩定生態系統結構和功能中發揮重要作用[4-5]。近年來,漁業主管部門在洪澤湖開展了翹嘴鲌人工放流,對于恢復翹嘴鲌資源發揮了重要作用,但盲目的增殖放流可能會對魚類資源遺傳結構產生不利影響[6]。因此,迫切需要了解洪澤湖的翹嘴鲌種質資源狀況和遺傳多樣性水平,用于指導洪澤湖翹嘴鲌增殖放流、種質資源保護和管理工作。
目前,分子標記技術被廣泛用于研究物種群體的遺傳多樣性和遺傳結構。由于線粒體DNA (mitochondrial DNA, mtDNA)具有分子小、結構簡單、進化快、嚴格的母系遺傳等特點,因此成為群體遺傳學和分子系統學的重要分子標記[6]。Cytb基因(線粒體細胞色素b基因)是mtDNA的13個蛋白編碼基因之一,其結構和功能較為清晰,進化速率適中,包含著從種內到種間的較強進化信號,故被廣泛用于物種多樣性和系統進化等方面的研究[7-9]。王利華等[10]應用COⅠ和Cytb基因分析了6種鲌屬魚類(達氏鲌、海南鲌、尖頭鲌、蒙古鲌、擬尖頭鲌和翹嘴鲌)的遺傳關系,認為線粒體COⅠ和Cytb基因可以作為DNA條形碼鑒定鲌屬魚類。李大命等[11]采用Cytb基因序列研究了滆湖3種鲌屬魚類(翹嘴鲌、蒙古鲌、達氏鲌)的遺傳多樣性,發現3種鲌魚的遺傳結構趨于單一化。張桂寧等[12]通過Cytb基因序列分析發現,長江下游4個翹嘴鲌群體的遺傳多樣性較高。2009年,伊西慶[13]采用線粒體ND2基因序列對采集的16尾洪澤湖翹嘴鲌進行了遺傳分析,檢測到較高的單倍型多樣性和較低的核苷酸多樣性,但該研究所分析的樣本量較少,其結果無法準確反映洪澤湖翹嘴鲌的遺傳狀況。近十多年,尚未有關于洪澤湖翹嘴鲌遺傳多樣性的研究報道。本研究擬采用線粒體Cytb基因序列,對6個洪澤湖翹嘴鲌群體的遺傳多樣性和遺傳結構進行分析,以期了解當前洪澤湖翹嘴鲌群體的遺傳多樣性狀況,并為翹嘴鲌增殖放流、種質資源保護及利用提供科學的理論依據。
1 材料和方法
1.1 樣品采集和處理
2021年在洪澤湖開展漁業資源監測,采用三重刺網在6個采樣點采集翹嘴鲌210尾,其中洪澤群體(HZ)35尾,高渡群體(GD)34尾,顧勒河群體(GLH)34尾,新開河群體(XKH)33尾、臨淮群體(LH)36尾,馬浪崗群體(MLG)38尾。各群體樣品采樣點位置見表1。測量每尾魚的體長,稱取體質量,并剪取尾鰭組織放入無水乙醇中保存。
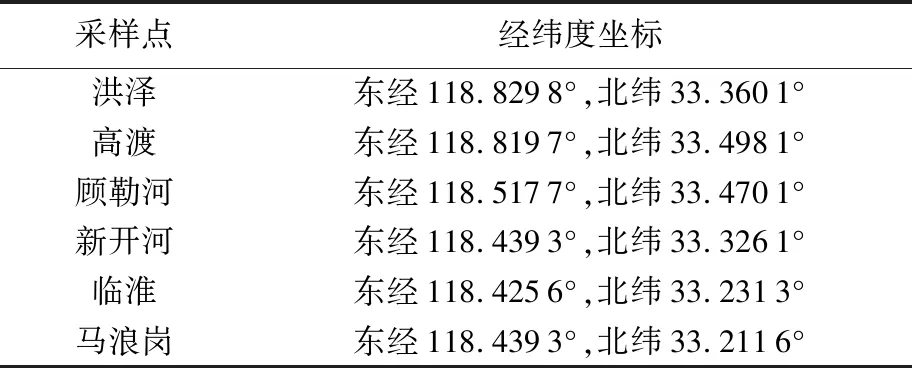
表1 洪澤湖翹嘴鲌各群體采樣點位置
1.2 DNA提取、PCR擴增和測序
采用TaKaRa公司試劑盒提取翹嘴鲌的基因組DNA,將DNA溶于TE溶液,于-20 ℃保存備用。用瓊脂糖凝膠電泳和紫外分光光度計檢測DNA質量和濃度。
線粒體Cytb基因擴增引物為L14724(序列為5’-CGTCAGTCCTTTACTTCGCA-3’)和H15915(序列為5’-AGGGCATACTCACGGGGTTG-3’)[14],引物由生工生物工程(上海)股份有限公司合成。PCR反應體系共50 μL,其中包括:2×Premix TaqTM25 μL(TaKaRa Taq 1.25 U/25 μL、0.2 mM dNTP、1.5 mM Mg2+、色素Marker、比重增加物和穩定劑)、上、下游引物各2 μL (10 μmol/L)、DNA模板2 μL (40 ng/μL )和ddH2O 19 μL。PCR擴增程序為:94 ℃預變性4 min;94 ℃變性40 s,55 ℃復性30 s,72 ℃延伸1 min,35個循環;72 ℃延伸5 min。PCR反應完成后,取4 μL產物經1%瓊脂糖凝膠電泳檢測,剩余的PCR產物送生工生物工程(上海)股份有限公司進行純化和雙向測序,測序引物與擴增引物一致。
1.3 數據處理和分析
使用BioEdit 7.0.9.0軟件[15]讀取序列,并參照測序圖譜人工校對序列中可能的錯誤。使用CLUSTALX 2.0軟件[16]對序列進行多重比對和排序。使用MEGA 7.0.26軟件[17]統計序列的堿基組成、突變位點數和突變類型,計算群體間的Kimura雙參數遺傳距離,利用鄰接法(neighbor-joining, NJ)構建單倍型系統發育樹,系統樹中節點的自舉置信水平應用自引導估計,循環次數為1 000次。采用Network軟件構建單倍型網絡圖,用以展示單倍型之間的進化關系。使用DanSP 5軟件[18]計算群體的單倍型數量、單倍型多樣性(haplotye diversity,Hd)、核苷酸多樣性(nucleotide diversity,Pi)、平均核苷酸差異數(K)等遺傳多樣性參數。采用Arlequin 3.5軟件[19]進行分子方差分析(AMOVA),計算群體間的遺傳分化指數(Fst),開展Tajima’sD及Fu’sFs中性檢驗和構建核酸錯配分布曲線,評估翹嘴鲌群體的歷史動態。
2 結果
2.1 Cyt b基因序列變異及遺傳多樣性
本研究獲得翹嘴鲌的Cytb基因全序列,其長度為1 141 bp。4種堿基的平均含量分別為A(29.4%)、T(26.6%)、C(29.4%)和G(14.6%),堿基A+T含量(56.0%)大于G+C含量(44.0%),具有明顯的堿基組成偏倚性。210條Cytb序列中共發現31個變異位點,其中有11個單突變位點,20個簡約信息位點。
6個群體210尾翹嘴鲌共定義了26個單倍型,整個群體的單倍型多樣性和核苷酸多樣性分別為0.735±0.032和0.001 9±0.000 1,平均核苷酸差異數為2.118(見表2)。6個群體中,馬浪崗群體(MLG)的遺傳多樣性最高(Hd為0.788±0.064,Pi為0.0021±0.0002),高渡群體(GD)的遺傳多樣最低(Hd為0.618±0.096,Pi為0.001 5±0.000 3)。洪澤湖翹嘴鲌的遺傳多樣性具有典型的高單倍型多樣性和低核苷酸多樣性模式。

表2 洪澤湖翹嘴鲌群體遺傳多樣性參數
2.2 群體單倍型組成及系統發育
6個群體定義了26個單倍型,其中洪澤群體擁有14個單倍型,高渡群體擁有11個單倍型,顧勒河和馬浪崗群體各擁有12個單倍型,新開河和臨淮群體各有10個單倍型。26個單倍型中,共享單倍型有18個,獨享單倍型有8個。其中,共享單倍型Hap3和Hap6為6個群體共有,個體數量分別有17個和105個,占整個群體的比例合計為58.1%;共享單倍型Hap1、Hap2、Hap4、Hap8、Hap11~12、Hap14~17和Hap19~24為部分群體共有,個體數量共有80個,占整個群體的比例合計為38.1%;獨享單倍型Hap5、Hap7、Hap9、Hap10、Hap13、Hap18、Hap25和Hap26為各自群體獨有,個體數量均只有1個,占整個群體的比例合計為3.8%(見表3)。整體來看,共享單倍型的個體數量多,占群體的比例高,暗示6個群體間存在廣泛的基因交流。
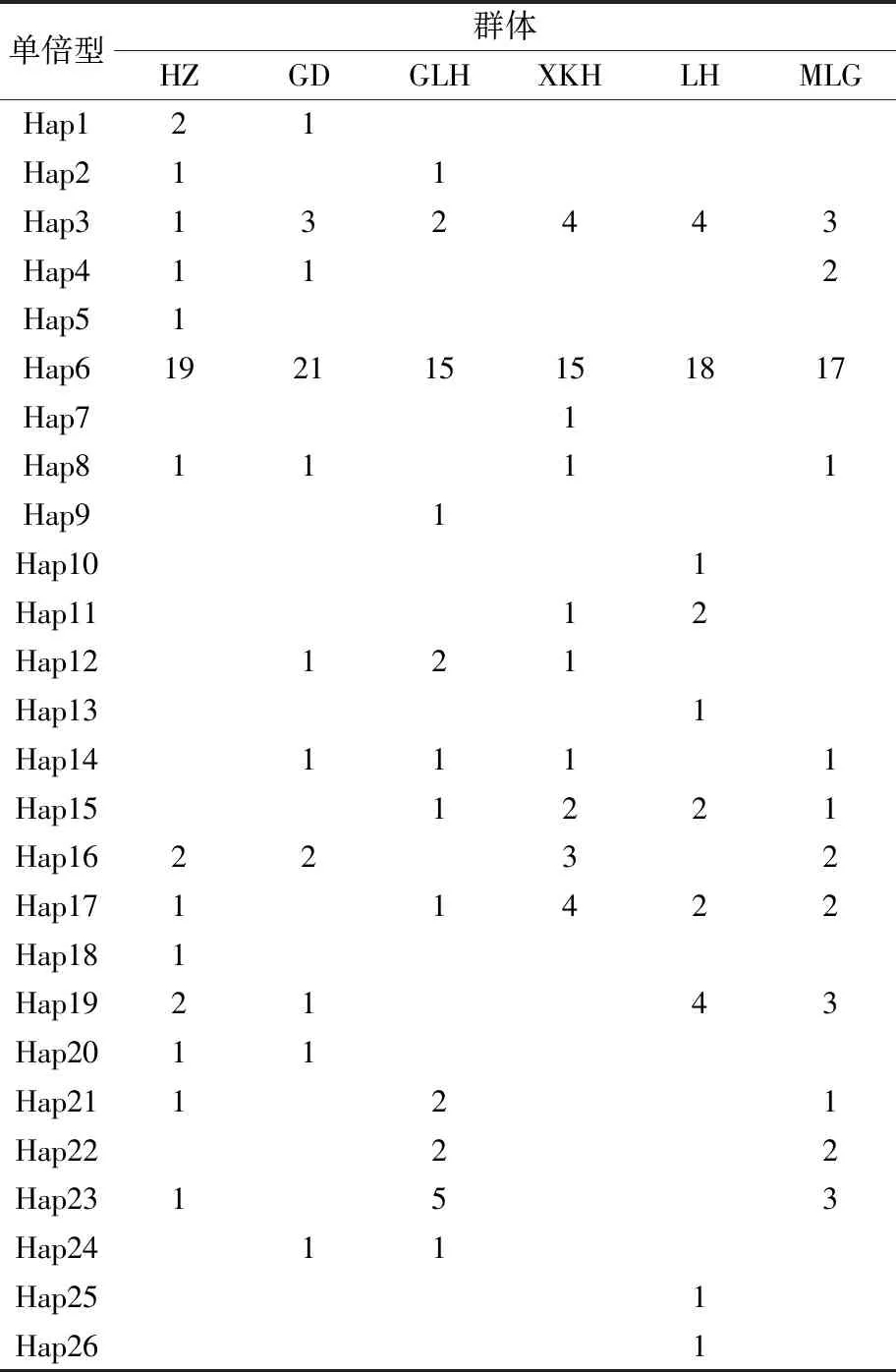
表3 洪澤湖6個翹嘴鲌群體單倍型組成
利用紅鰭原鲌(Cultrichthyserythropterus)和蒙古鲌(Cultermongolicus)作為外類群,基于鄰接法構建單倍型系統發育樹(見圖1)。結果顯示,所有單倍型聚為一個進化分支,分支的置信限水平為100%。單倍型在6個群體中呈混雜分布模式,沒有形成與群體地理位置相對應的譜系關系。單倍型網絡結構圖也得出類似結果(見圖2),說明6個群體間沒有明顯的遺傳分化。
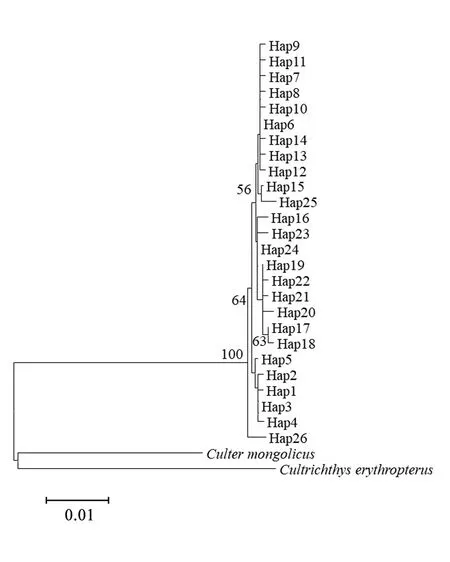
圖1 洪澤湖翹嘴鲌群體單倍型NJ系統樹
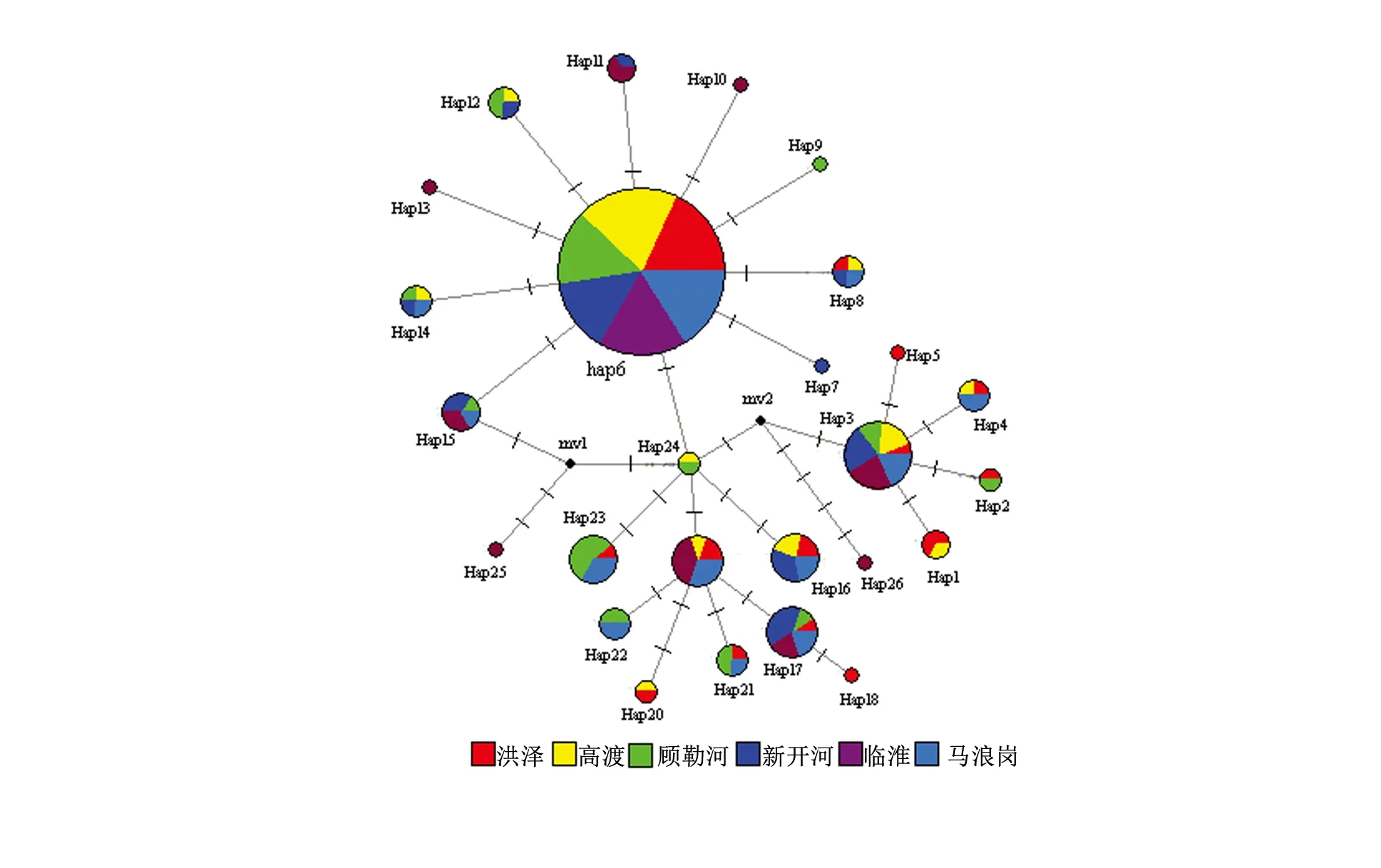
圖2 洪澤湖翹嘴鲌群體單倍型網絡結構圖
2.3 群體間遺傳結構
6個群體間的遺傳距離為0.001 6~0.002 0,遺傳距離均值為0.001 9,表明群體間遺傳差異較小,親緣關系較近(見表4)。分子方差分析結果顯示,群體內個體間的分子變異占比為100.33%,群體間遺傳變異占比為-0.33%,遺傳分化指數Fst值為-0.003 3(P=0.624 6)(見表5),表明整個翹嘴鲌群體沒有出現遺傳分化。兩兩群體間的Fst值為-0.017 7~0.018 2,且統計檢驗結果均不顯著(見表4),顯示群體間沒有形成顯著的遺傳分化。
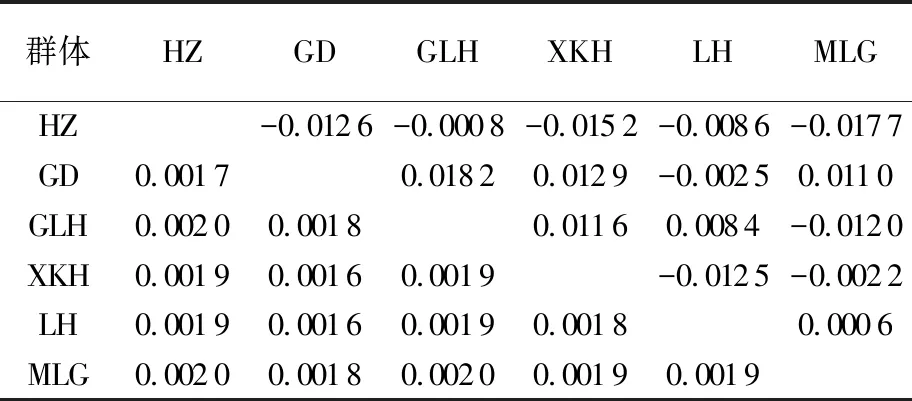
表4 洪澤湖翹嘴鲌6個群體間的遺傳距離(對角線下)和遺傳分化指數(對角線上)
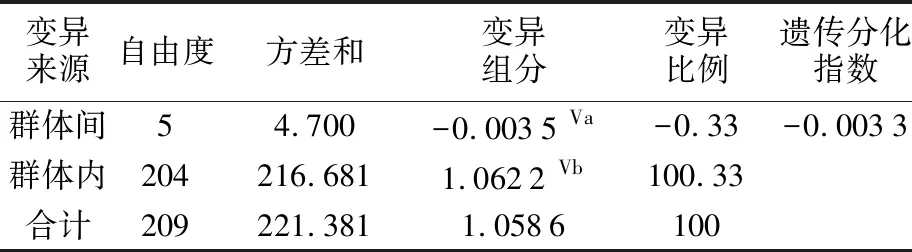
表5 洪澤湖翹嘴鲌群體分子方差分析
2.4 群體歷史動態
采用Tajima’sD、Fu’Fs中性檢驗和核苷酸錯配分布曲線分析翹嘴鲌群體的歷史動態。結果顯示,6個群體的Tajima’sD檢驗值統計均無顯著性差異(P>0.05),除臨淮群體(LH)外,Fu’Fs檢驗值統計均有顯著性差異(P<0.05)(見表6),且6個群體的核苷酸錯配分布曲線呈現出單峰模式(見圖3),表明6個群體顯著偏離中性進化。從整體來看,其Tajima’sD和Fu’Fs的檢測值分別為-1.683 2和-13.532 9,統計分析具有顯著性差異(P<0.05),且核苷酸分布曲線也具有典型的單峰模式(見圖3),表明翹嘴鲌在進化過程中發生了群體擴張事件。
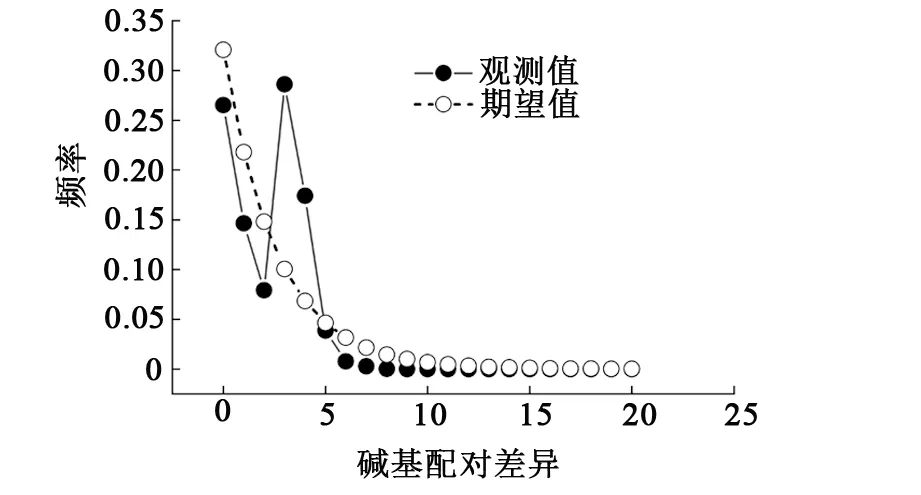
圖3 洪澤湖翹嘴鲌群體核苷酸錯配分布曲線
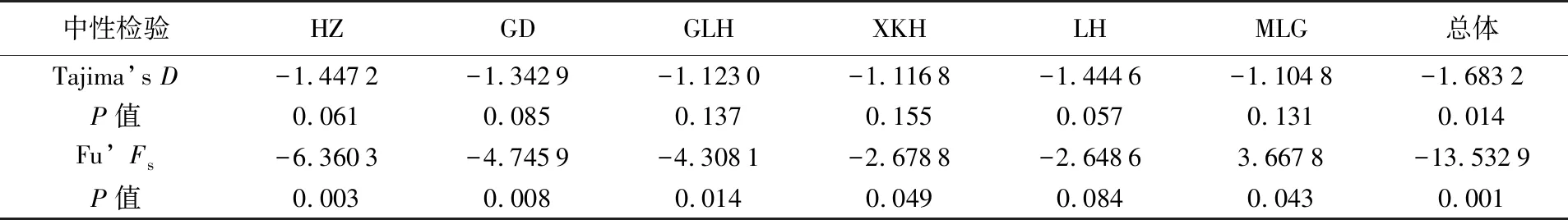
表6 洪澤湖翹嘴鲌群體的中性檢驗
3 討論
3.1 翹嘴鲌的遺傳多樣性
遺傳多樣性是生物多樣性的基礎,也是評價生物種質資源狀況及制定種質資源保護措施的重要依據。單倍型多樣性(Hd)和核苷酸多樣性(Pi)是衡量群體遺傳多樣性的重要指標,其數值越大,表明群體遺傳變異越豐富,遺傳多樣性也越高[20-21]。相比較而言,核苷酸多樣性考慮各種單倍型在群體中所占的比例,能更為精確地反映群體多態性程度[22]。本研究結果顯示,洪澤湖翹嘴鲌的平均單倍型多樣性和核苷酸多樣性分別為0.735±0.032和0.001 9±0.000 1,6個群體的單倍型多樣性在(0.618±0.096)~(0.788±0.064),核苷酸多樣性為(0.001 5±0.000 3)~(0.002 1±0.000 2)。Grant等[23]根據單倍型多樣性和核苷酸多樣性大小,將單倍型多樣性和核苷酸多樣性之間的關系分為4種類型:高Hd高Pi、高Hd低Pi、低Hd高Pi和低Hd低Pi。據此判定,洪澤湖翹嘴鲌的遺傳多樣性屬于高Hd(Hd≥0.5)低Pi(Pi<0.005)類型,這種遺傳多樣性模式暗示翹嘴鲌的遺傳多樣性較低[23]。多年來,受過度捕撈、環境污染、非法采砂等不利因素的影響,洪澤湖環境生態質量下降,翹嘴鲌漁業資源量明顯減少,其適宜的棲息環境大幅縮減,個體小型化和種群低齡化趨勢加劇,遺傳多樣性喪失。已有研究結果顯示,我國許多水體的野生翹嘴鲌資源遺傳多樣性具有高Hd低Pi共存現象[11,13,24-26],表明翹嘴鲌遺傳多樣性較低是一個普遍現象,需要采取措施恢復翹嘴鲌資源量及提高其遺傳多樣性。
3.2 翹嘴鲌的歷史動態
魚類群體的遺傳多樣性模式與群體的歷史動態有密切的聯系。Grant等[23]提出了一個較簡單的模式,即利用Hd、Pi來推測群體的歷史動態:當Hd≥0.5、Pi≥0.005時,表示群體穩定,具有較悠久的進化歷史;當Hd≥0.5、Pi<0.005時,表示群體是經歷瓶頸效應后迅速擴張而來的;當Hd<0.5、Pi≥0.005時,表明群體經歷了輕微的瓶頸效應,幾乎沒有影響到核苷酸變異;當Hd<0.5、Pi<0.005時,表明群體近期經歷了瓶頸效應。本研究結果顯示,翹嘴鲌群體的單倍型多樣性較高(Hd≥0.5),而核苷酸多樣性較低(Pi<0.005),由于積累核苷酸多樣性所需的時間比積累單倍型所需的時間漫長,表明洪澤湖翹嘴鲌群體是從一個有效種群數量較小的種群經快速擴張而來的,但是仍然沒有達到積累核苷酸變異所需要的時間。通常情況下,有2種方法可用于推測群體的歷史動態:(1)核苷酸錯配分布曲線。若分布曲線呈單峰分布,表明群體經歷了種群擴張;若分布曲線呈雙峰分布,表明群體沒有經歷過種群擴張,較為穩定[27-28]。(2)Tajima’sD和Fu’Fs中性檢驗。若中性檢驗值為負值且統計分析結果為顯著,表明群體可能經歷了種群擴張[29-30]。本研究結果顯示,洪澤湖翹嘴鲌群體的Tajima’sD和Fu’Fs中性檢驗值均為負值,統計檢驗結果具有顯著性差異(P<0.05);核苷酸錯配分布曲線呈典型的單峰型,均說明翹嘴鲌進化過程中偏離了中性選擇,經歷了顯著的種群擴張,形成了高Hd和低Pi的遺傳多樣性模式,這與已有的研究結果[11,13]一致。
3.3 翹嘴鲌的遺傳結構
通常采用遺傳分化指數(Fst)來評估群體間的遺傳分化程度。Wright[31]指出,若Fst值為0~0.05,表明群體間不存在分化;若Fst值為0.05~0.15,表明群體間存在中度分化;若Fst值為0.15~0.25,則為高度分化。分子方差分析結果顯示,遺傳變異主要發生在群體內個體間,整個群體的Fst值為-0.003 3(P=0.624 6),兩兩群體間的Fst值為-0.017 7~0.018 2,P>0.05,均表明翹嘴鲌群體間沒有出現遺傳分化,群體的分子系統發育樹和單倍型網絡結構圖也支持上述觀點。從群體的單倍型組成來看,群體間有多個共享單倍型,且共享單倍型的個體數量占比較高,說明群體間存在廣泛的基因交流。對于魚類而言,地理阻隔或同一水域中存在不同的棲息環境可能會限制基因交流,從而使群體間產生遺傳分化現象[32]。本研究中的6個群體雖處于洪澤湖的不同水域,水域生態環境也存在一定的差異[33-34],但群體所在的地理位置較近,沒有形成有效的地理隔離,而翹嘴鲌游泳能力強,活動范圍廣,也助推了群體間的基因交流。此外,近年來漁業主管部門在洪澤湖開展了翹嘴鲌增殖放流活動,大量人工養殖群體的加入容易導致群體遺傳結構的單一和均質化,這一現象在其他水域的增殖放流種類中均有發現[35-37]。根據翹嘴鲌的遺傳結構特點,應將6個群體作為一個進化單位進行管理和保護。
3.4 翹嘴鲌資源保護對策
對魚類群體遺傳多樣性和遺傳結構的研究可以為漁業資源管理措施及物種保護策略的制定提供重要的數據支撐。研究結果顯示,洪澤湖翹嘴鲌的遺傳多樣性具有高單倍型多樣性和低核苷酸多樣性共存現象,表明翹嘴鲌遺傳資源較為貧乏。本研究洪澤湖翹嘴鲌6個群體間沒有顯著的遺傳分化,因此可將洪澤湖不同水域的翹嘴鲌群體作為一個整體進行管理和保護。針對洪澤湖翹嘴鲌的遺傳多樣性現狀,應當采取措施,加大對翹嘴鲌的保護力度:(1)強化漁業資源管理,加大執法力度,嚴厲打擊非法捕撈,禁止電、毒、炸等作業方式;(2)控制環境污染,開展生態環境治理,恢復水生植被,構建人工魚巢,為翹嘴鲌生存和繁殖提供良好的生態環境;(3)科學規范地進行翹嘴鲌增殖放流,提高翹嘴鲌的成活率,開展放流群體和野生群體的遺傳評估,跟蹤監測翹嘴鲌資源變化,逐步提高翹嘴鲌的遺傳多樣性。
