天然產物色胺酮的研究進展
2023-11-29 11:33:02張定堃
天然產物研究與開發 2023年11期
關鍵詞:小鼠
廖 維,宋 嬌,韓 麗,張定堃
成都中醫藥大學藥學院 西南特色中藥資源國家重點實驗室,成都 611137
色胺酮(見圖1),化學名為吲哚[2,1-b]喹唑啉-6,12-二酮(分子式C15H8N2O2),常溫下呈黃色針狀結晶。色胺酮屬于吲哚喹唑啉類生物堿,是傳統抗病毒中藥板藍根和解熱中藥青黛及其藥用植物馬藍、菘藍、蓼藍等的主要成分之一,某些微生物的代謝產物中也能檢測到[1-4]。現代藥理學研究表明色胺酮具有抗腫瘤、抗菌、抗炎、抗病毒、保護心血管等多種藥理活性[5]。色胺酮在腫瘤治療、細菌感染、炎癥損傷、病毒侵害等方面展現出的較高藥用價值,使之成為眾多學者關注的焦點。細胞毒性試驗與小鼠毒性試驗均證實其安全性較好[6-8]。本綜述對色胺酮的相關文獻進行系統梳理,綜合分析了色胺酮的來源、藥理作用、藥動學特征、毒性等方面的研究進展情況,旨在為色胺酮作用機制的深入研究提供更多最新、最全面的信息,最終為臨床上色胺酮類藥物的開發提供新的思路。
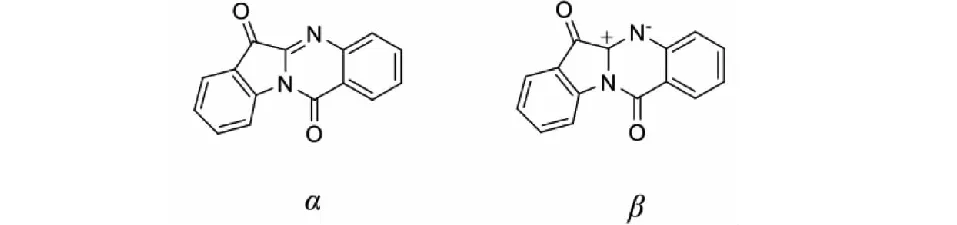
圖1 色胺酮的兩種構型Fig.1 Two conformations of tryptanthrin
1 色胺酮的來源
1.1 天然植物來源及其生物合成前體
色胺酮的發現不同于一般天然產物。大多天然產物是先從植物中分離出來,而后對其進行藥理與合成方面的研究。而色胺酮的人工合成比發現它是一種天然產物早了半個世紀,但產率極低,致使這個具有良好藥理活性的小分子化合物在很長時間內無人問津[9]。1997年,Masayuki等[10,11]從兩種蘭科植物Calanthediscolor和C.liukiuensis中分離出了一種糖苷(calanthoside)并發現其酶水解的主要產物是色胺酮。除菘藍、蓼藍、馬藍等藥用植物外(色胺酮含量分別為0.003%、0.006%、0.001%)[3,4],中藥青黛中也含有色胺酮(0.01%~0.006 5%)[1,2]。此外,色胺酮的含量受產地、采收時間及處理方式等因素的影響。以中國福建省為例,12批不同產地的馬藍中,產于南靖和溪的馬藍莖、葉中色胺酮總含量最高(244.56 μg/g)[12]。對不同生長時期板藍(Baphicacanthuscusia)中色胺酮的含量進行測定,發現色胺酮的含量變化趨勢不太穩定,整體看來十月份是其采收的最佳時期[13]。Oberthür等[14]檢測了不同方法處理后菘藍中色胺酮的含量,發現40 ℃烘干葉片中色胺酮含量最高(0.18%~0.33%),冷凍干燥葉片中的含量最低(0.034%~0.055%),其次是室溫干燥葉片(0.06%~0.207%),而經發酵的葉片中色胺酮的含量低于檢測限。提示色胺酮的產生是一個后熟過程,干燥和高溫可能促進了色胺酮的形成,且該過程是非酶催化的,因為烘干會使酶失活而無催化作用。隨后,有研究人員分別用烘干、陰干、凍干三種方法對馬藍進行處理,發現陰干法處理的馬藍根、莖、葉中色胺酮含量相對最高,凍干法處理的色胺酮含量最低[15]。其原因可能是,相對烘干法和凍干法,陰干法干燥溫度適中,干燥時間較長,在藥材未完全干透之前有利于色胺酮的后熟,致使其含量升高。
隨后,有學者探尋了色胺酮在植物體內的生物合成前體。分析色胺酮的結構,其可能是由靛紅與鄰氨基苯甲酸經脫水縮合而成。為此Wang等[9,16]選取了藥用植物菘藍作為研究對象,來驗證猜測的正確性。成分檢測發現,新鮮菘藍葉片中不含色胺酮,但有靛紅,葉片經烘干、晾干或研磨處理后靛紅消失,色胺酮生成。前體追加實驗結果顯示,追加靛紅或鄰氨基苯甲酸或者兩者都加均能大幅度提高色胺酮的含量,表明色胺酮與靛紅及鄰氨基苯甲酸存在生源關系。結合色胺酮和靛紅分布實驗,可以證實靛紅和鄰氨基苯甲酸是色胺酮的合成前體,同時也進一步證實了色胺酮的形成是化學催化而非酶催化。此外,關于植物中莽草酸酯和L-Trp生物合成途徑的現有知識,以及色胺酮的逆生物合成分析表明,鄰氨基苯甲酸和N-甲酰苯胺酸也可能是色胺酮的主要生物合成前體。2018年,Pedras等[17]基于同位素標記化合物中氘的引入,通過HPLC-DAD-ESI-MS對青黛葉片提取物進行分析,并使用ESI-MS光譜的峰值強度確定了各代謝物中摻入同位素的百分比。該同位素標記化合物并入色胺酮的總體實驗結果表明,鄰氨基苯甲酸和N-甲酰氨基苯甲酸是色胺酮主要的生物合成前體(見圖2)。
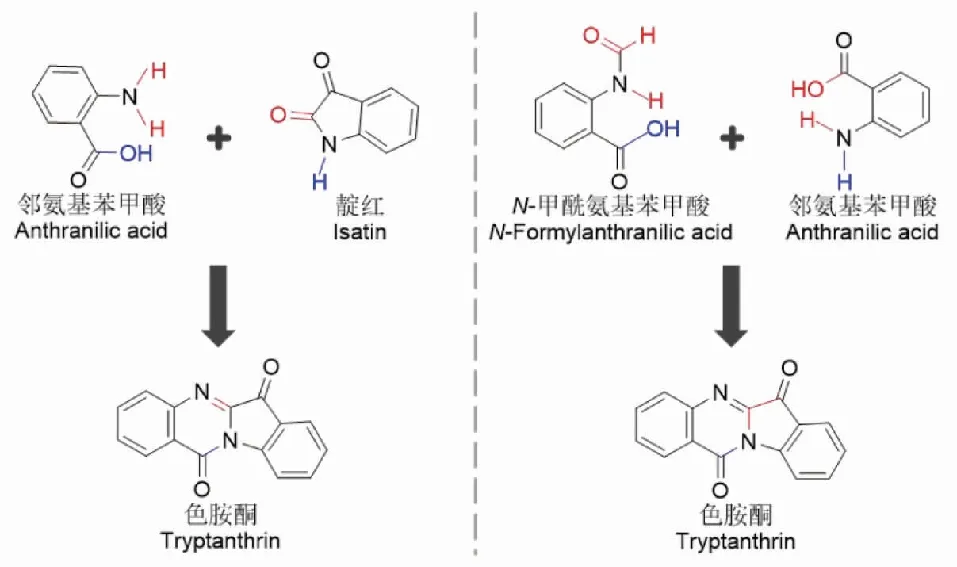
圖2 色胺酮的生物合成Fig.2 The biosynthesis of tryptanthrin
1.2 微生物途徑
除植物來源外,還能通過微生物發酵的方法獲得色胺酮。1971年,Schindler等[18]首次從過量L-色胺酸培養的解脂假絲酵母代謝產物中分離出了色胺酮。此外,擔子菌屬裂褶菌的發酵液、海洋微生物和大型生物中也能分離得到色胺酮及其衍生物[19]。有學者利用HPLC-DAD和LC-MS結合的方法,從海洋鏈霉菌Streptomycessp.B9173次級代謝產物中成功分離出了色胺酮[20]。隨后,Utkina等[21]從假儉魚中也分離得到色胺酮的衍生物ophiuroidine 1,這是首個在海洋無脊椎動物中發現的吲哚喹唑啉生物堿的例子,光譜數據確定其結構為4,8,9-三羥基吲哚[2,1-b]喹唑啉-6,12-二酮。微生物代謝產物中色胺酮的發現,可為色胺酮的合成生物學和代謝工程的研究提供新思路,以建立一個強有力的生物合成途徑,來滿足日益增長的需求。但目前天然產物在微生物宿主中的生物合成仍面臨挑戰,還需進一步研究,發展更新穎的合成生物學技術,進一步提升色胺酮生物合成的綜合效率。
1.3 人工合成
色胺酮在植物和微生物中含量極低,目前市場上主要通過人工合成獲得色胺酮,眾多合成方法如下[22-28](見圖3)。最初是通過靛紅自身反應得到色胺酮。陰極還原、高錳酸鉀水溶液處理或POCl3處理靛紅均能得到色胺酮,但存在產率低、反應條件不夠溫和等問題[22]。現最常用的方法是以靛紅和靛紅酸酐為初始底物合成色胺酮。靛紅和靛紅酸酐可在三乙胺的作用下,直接發生親核進攻和分子內的縮合脫水反應,以更簡潔的步驟得到色胺酮[24]。為提高實用性,Wang等[25]開發了一種新的色胺酮合成路徑,先將靛紅氧化成靛紅酸酐,再將靛紅酸酐與靛紅在堿性條件下得到目標產物,實現了無需催化劑參與、原子利用率高的合成目標,大大降低了反應成本,具有潛在應用價值。此外,以鄰氨基苯甲酸為底物,在二氯亞砜、羥胺和水等作用下,與靛紅發生環化也能得到色胺酮,產率可達85%[26]。隨后,Jia等[28]以靛紅和脒鹽作為底物,在室溫條件下,發展了一例TBHP/K3PO4協同促進的氧化環化策略,可用于合成喹唑啉酮類衍生物。相同的條件下,在缺少脒鹽底物時,靛紅極易發生自聚得到天然產物色胺酮,這為色胺酮的綠色合成提供了一條極具利用價值的新途徑。
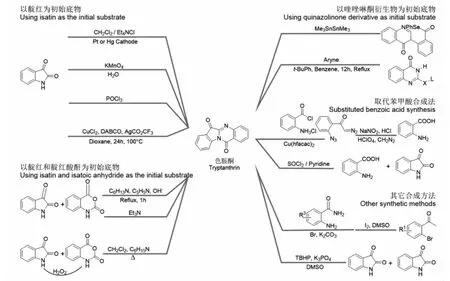
圖3 色胺酮的合成途徑Fig.3 The synthetic pathway of tryptanthrin
2 藥理作用
2.1 抗腫瘤作用
色胺酮類化合物是一種芳香烴受體激動劑,也是一種有效的IDO-1抑制劑,有成為抗腫瘤藥物的潛力[29]。天然產物色胺酮低毒性、強抗腫瘤活性的特點使之成為新藥開發研究的熱點之一,其抗腫瘤的作用機制也不斷被發現(見圖4)。

圖4 色胺酮的抗腫瘤作用Fig.4 The antitumor effects of tryptanthrin
2.1.1 白血病
青黛,在中國被廣泛用于白血病的治療,色胺酮作為青黛的活性成分也被證實具有抗白血病的作用。研究表明,色胺酮在體外對多種人類白血病細胞有較強的殺傷作用,低濃度的色胺酮(0.5 μg/mL)可誘導人類白血病細胞U-937分化為單核細胞/巨噬細胞,HL-60細胞分化為骨髓細胞/單核細胞,并增強CD11b的表達;高濃度的色胺酮(10 μg/mL)則可能通過caspase-3/Fas抗原途徑誘導細胞凋亡[30]。色胺酮在體外對人白血病細胞株K562也具有增殖抑制和誘導凋亡作用,且呈一定的濃度(3.12~50 mg/L)和時間(24~72 h)依賴性。相關分子機制可能是,一方面通過調節Bcl-2、Bax基因及蛋白的表達、降低線粒體跨膜電位、促使Cyt-c釋放至胞漿,刺激下游分子caspase-3的活化引起K562細胞發生凋亡;另一方面通過抑制融合基因bcr/abl及蛋白的表達,抑制K562細胞增殖和誘導凋亡。體內實驗發現,K562-SCID小鼠口服色胺酮(50 mg/kg)后病理癥狀得到明顯改善,髓組織中異型細胞明顯減少,脾臟淤血癥狀減輕,肝臟、肺、腎臟組織中淋巴細胞明顯減少,表明色胺酮對白血病具有治療作用[31,32]。在同系BALB/c小鼠體內,腹腔注射色胺酮(0.08、0.16 mg/kg)后,小鼠粒單核細胞白血病WEHI-3B JCS細胞的生長得到顯著抑制,其作用機制可能與引起細胞周期阻滯和誘導細胞分化有關[33]。
在人類和小鼠白血病細胞系如HL-60、U-937、K-562和WEHI-3B JCS,均證明色胺酮具有類似的抗增殖作用,表明其對白血病細胞的抗增殖作用不是物種或細胞系特異性的。提示色胺酮具有成為一種新的、有效的預防或治療白血病的先導化合物的潛力。
2.1.2 肝癌
色胺酮不僅能減緩氧化應激給肝臟帶來的危害,對肝癌細胞也有抑制作用。現普遍認為氧化應激是人體一種最基本的保護機制,但過度的氧化應激會導致細胞和組織損傷,引起高血壓、糖尿病、心血管等疾病。過度氧化應激是肝臟發病的病理生理基礎,可通過減少氧化應激或提高肝臟的抗氧化能力減少氧化應激帶來的危害。Moon等[34]發現色胺酮能有效抑制叔丁基過氧化氫(tBHP)介導的細胞毒性,使人肝細胞源性HepG2細胞免受氧化損傷和凋亡,包括活性氧(ROS)的產生、線粒體膜電位(MMP)的改變和細胞死亡,這種抑制可能與通過ERK/Nrf2途徑上調HO-1和GCLC蛋白表達有關。色胺酮還可抑制肝細胞生長因子(HGF)發揮抗肝癌的作用。HGF在腫瘤的侵襲、生長和轉移方面起著重要作用,與健康人相比肝癌患者體內HGF的表達明顯增加。具體而言,色胺酮對表皮成纖維細胞中的多種HGF誘導劑具有強烈的抑制作用,如乙酸肉豆蔻佛波醇酯(PMA)、表皮生長因素(EGF)、血小板生長因素(PDGF)和基本的纖維原細胞生長因素(bFGF)。與此同時,色胺酮可有效抑制EGF、PDGF和bFGF誘導的成纖維細胞增殖,且無毒副作用。但色胺酮不能減弱因PMA誘導引起的MAPK磷酸化作用,表明色胺酮抑制HGF的產生可能是通過MAPK活化的下游過程來實現的[9]。Liang等[35]在篩選抗腫瘤藥物過程中發現,色胺酮呈劑量依賴性(3~50 mg/mL)抑制人肝癌細胞BEL 7402的體外增長,當濃度為50 mg/L時抑制率可達88.02%。進一步用半數抑制濃度(8.2 μg/mL)的色胺酮培養1、3、5天后,BEL-7402細胞端粒酶活性A值分別為0.381、0.276和0,而未經藥物處理的肝癌細胞A值為1.60,表明色胺酮能抑制BEL-7402細胞的端粒酶活性,且抑制效果隨色胺酮作用時間增加而增加。此外,集落形成實驗也表明色胺酮具有逆轉腫瘤細胞向正常細胞轉化的能力。上述研究表明,色胺酮在肝癌的預防、治療與康復全過程均能發揮功效,且毒性較低,具有開發成抗肝癌新藥的前景。但上述研究均為細胞實驗,目前還缺乏體內試驗來證明色胺酮抗肝癌的有效性和安全性,因此還需要對色胺酮進行深入地研究,以充分發揮其抗腫瘤活性。
2.1.3 乳腺癌
乳腺癌是全世界女性發病率最高的惡性腫瘤,其術后復發率和轉移率極高,化療也易出現不同程度的副作用,如神經毒性、肝腎功能損傷、胃腸道反應等。因此從中藥中尋找療效好、毒副作用低的乳腺癌術后和放化療輔助藥物,具有重要的現實意義。色胺酮能上調E-cadherin蛋白水平、下調MMP-2和Snail在人乳腺癌MCF-7細胞中的表達,從而抑制MCF-7細胞的增殖、遷移和侵襲,還能抑制TGF-β1誘導的MCF-7細胞轉化。4T1乳腺癌小鼠灌胃色胺酮(25、50、100 mg/kg)后,食欲更好,肢體運動范圍更大,腫瘤生長得到顯著抑制,且不會引起體重和臟器系數的波動(P>0.05)。相關的藥理機制是下調小鼠腫瘤組織中NOS1、COX-2和NF-κB P65蛋白的表達,上調小鼠血清中IL-2和TNF-α,并恢復小鼠血清IL-10水平[56]。進一步的Label-free蛋白質組學實驗結果提示色胺酮可能通過調節與炎癥相關的蛋白發揮抗小鼠體內4T1乳腺癌的作用,與之密切相關的差異表達蛋白包括上調CD14、PTGS2,泛素蛋白連接酶E3和下調CD44、HSPA1A、MIF、NF-κB、RPS6KA4、HMGB1[36,37]。MAPK途徑在乳腺癌的增殖、生長和化療耐藥中起著重要作用,參與乳腺癌的侵襲和轉移過程。為考察色胺酮能否通過MAPK信號通路對MCF-7細胞增殖產生影響,Zhou等[38]將色胺酮與MAPK抑制劑PD98059、SB203580和SP600125聯合使用對MCF-7細胞進行處理。結果顯示,單獨給予色胺酮處理后MCF-7細胞活力明顯下降,細胞中p-ERK、p-p38 MAPK和p-JNK蛋白表達均有增加的趨勢;而聯合給予色胺酮和不同濃度的MAPK通路抑制劑后,細胞的增殖活力均有不同程度的上升,細胞中p-ERK表達有所下降,p-p38 MAPK和p-JNK表達變化不明顯。提示色胺酮對MCF-7細胞的抑制作用可能與MAPK信號通路激活有關,與ERK以及JNK通路的關系尤為密切。此外,Yu等[39]驚喜地發現色胺酮具有下調多藥耐藥性(MDR)的功能,通過下調MDR1基因的表達,發揮逆轉乳腺癌細胞阿霉素耐藥株耐藥性的作用。相對于人體內其他細胞而言,癌細胞生長同樣需要適應的環境,于是Zeng等[36]將目光投向癌癥與人體微環境之間的聯系—腫瘤微環境。結果表明,在體內和體外,色胺酮均能通過調節腫瘤炎癥微環境發揮抗乳腺癌作用。
綜上所述,色胺酮能從多個方面抑制乳腺癌的發生發展,主要機制是:抑制乳腺癌細胞的增殖、侵襲和遷移,促進凋亡,調節腫瘤炎癥微環境等。此外,色胺酮還表現出較好的MDR逆轉作用,且安全性較好。鑒于此,可考慮將色胺酮與臨床常用的某些一線化療藥物聯用,增強療效的同時克服耐藥。
2.1.4 皮膚癌
皮膚癌可分為基底細胞癌、鱗狀細胞癌和黑色素瘤三大類,多發于臉頰、鼻部、眼瞼皮膚等長期暴露于陽光的部位。其中黑色素瘤僅占皮膚癌總體的2%,但其惡性程度高,侵襲、遷移能力強,傳統的放化療治療效果不理想。早在1985年便有報道稱色胺酮對黑色素瘤細胞株B16有抑制作用[40]。Jayesh等[41]也發現從藥用植物Wrightiatinctoria(Roxb.) R.Br.葉子中分離出來的半純化組分DW-F5在體外和體內均能阻斷黑色素瘤生長。光譜分析表明,DW-F5由色胺酮、一種芳香化合物和一種未經鑒定的脂肪族成分組成。此外,Shankar等[42]還研究了色胺酮對非黑色素瘤皮膚癌(NMSC)的作用,并用瑞士白化小鼠皮膚癌變模型對色胺酮的功效進行評估。在小鼠模型中,色胺酮(0.5、1 mg)成功地使腫瘤發病率、大小和多樣性大幅度減少。經色胺酮處理后,小鼠毛囊細胞增殖和擴張受阻,表皮細胞中炎性細胞(中性粒細胞和淋巴細胞)浸潤減少,β-連環蛋白、細胞周期蛋白D1和c-Myc表達降低,但未觀察到小鼠皮膚或腫瘤中細胞凋亡水平的顯著變化。其機理是通過抑制炎癥和阻止毛囊細胞中的β-catenin和相關信號通路來抑制皮膚癌的發生,從而阻止其擴張和隨后的腫瘤發展。
2.1.5 腸癌、食管癌
色胺酮對一些較為常見的消化道惡性腫瘤如腸癌和食管癌也有一定的治療效果。Koya等[43]研究了口服色胺酮對azoxymethy(AOM)誘發的F344大鼠腸腫瘤的影響。結果顯示色胺酮組(50 mg/kg BW)小腸腫瘤發生率低于AOM對照組(5% versus 26%),表明色胺酮能降低腸道腫瘤的發生率,對腸癌有預防作用。順氯氨鉑(cDDP)是食管癌后期化療的主要藥物,但其耐藥性的產生極大地影響了化療效果。Huang等[44]發現色胺酮(16 mg/L)能增強順鉑對癌細胞的敏感度,抑制MDR1和GST-pi基因的表達逆轉食管癌細胞順鉑耐藥株Eca109/cDDP的耐藥性,表明色胺酮一種潛在的食管癌術后化療輔助藥物。
2.1.6 神經母細胞瘤
神經母細胞瘤主要起源于腎上腺,是兒童期最常見的顱外實體瘤,約占所有兒童癌癥的10%。傳統的放化療會有殘留病變、局部復發以及不良反應出現的可能,因此對于神經母細胞瘤的治療仍是兒科腫瘤學面臨的一個重大挑戰[45]。為尋找更安全有效的藥物,研究人員用N-myc擴增技術研究了色胺酮在體外對人神經母細胞LA-N-1細胞生長和分化的影響。結果表明色胺酮對人神經母細胞瘤細胞的生長抑制具有劑量(0~30 μmol/L)和時間(24、48、72 h)依賴性。其機制可能是誘導細胞周期阻滯于G0/G1期;增強乙酰膽堿酯酶活性和誘導LA-N-1細胞向神經元分化;降低LA-N-1細胞中N-myc的表達[46]。
2.2 抗炎
自古以來菘藍便有治療刀傷、潰爛、蛇咬、痔瘡等炎癥的記載,現多用于治療風濕性關節炎、皮炎、哮喘、結腸炎等疾病[47]。作為菘藍的活性成分,色胺酮的抗炎作用也不容忽視(見圖5)。體外實驗證明,色胺酮能抑制與炎癥相關的因子如NO、PGE2、PG、LTs的合成影響炎癥的反應進程[48,49]。此外,色胺酮(1~10 mmol/L)還能抑制肥大細胞的增殖,有用于治療肥大細胞介導的特應性皮炎的潛能。其作用機制是:抑制胸腺基質淋巴生成素刺激的HMC-1細胞中Ki67、MDM2和IL-13的表達;增加HMC-1細胞中PARP和caspase3的表達;降低TSLP誘導的TNF-α產生和IL-7Ra/TSLPR mRNA表達[50]。另一細胞實驗發現,色胺酮(10 mmol/L)可抑制LPS刺激的BV2小膠質細胞中促炎介質和細胞因子(如TNF-α、IL-6、IL-1b、iNOS、NO、COX-2和PGE2)的過度生成和TLR4和MyD88蛋白的表達,發揮抗神經炎癥的作用,這與色胺酮促使NF-κB通路失活,抑制p38 MAPK途徑的激活有關[6,51]。
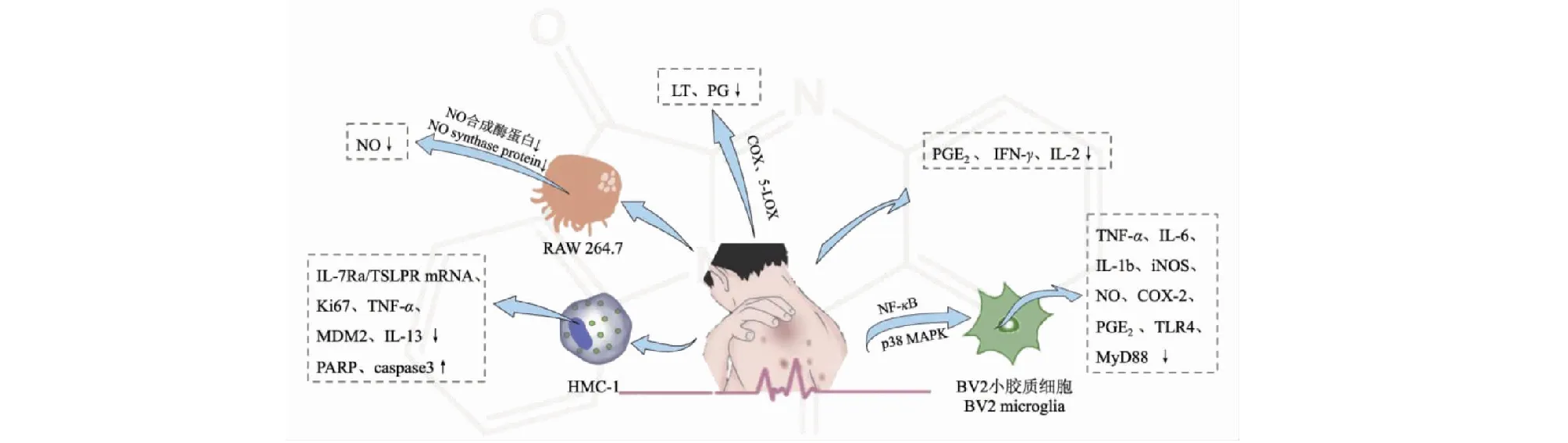
圖5 色胺酮的抗炎作用Fig.5 The anti-inflammatory effect of tryptanthrin
2.3 抗菌
抗生素的濫用導致多重耐藥、泛耐藥菌株不斷增加,可供臨床選擇的抗生素越來越局限。研發新型抗生素的周期較長,故從天然植物中挖掘毒副作用小的活性成分逐漸成為現代抗菌藥物研究的熱點。色胺酮是菘藍根抗菌作用的有效成分,且在體內外均有一定的抗菌作用。在體外,色胺酮能夠呈劑量依賴性抑制幽門螺桿菌(1.0~10 μg/mL),濃度為10 μg/mL時菌落形成被完全抑制。體內實驗發現,與溶劑對照組比,口服色胺酮(5 mg/只)能顯著抑制蒙古沙土鼠胃中的細菌菌落。此外,色胺酮與克拉霉素聯用時還能發揮協同效應,為降低抗生素的使用劑量提供可能性,從而減輕副作用[52]。色胺酮(亞-MIC 2 μg/mL)對霍亂弧菌也有較好的抑制作用,其靶點為群體感應(Quorum sensing)調節蛋白LuxO。溶血性實驗發現,色胺酮在12 h內溶血率至少為0.72%,在第12 h至少為2.78%(溶血<10%通常被認為是安全的),說明其亞-MIC濃度為非溶血性[7]。色胺酮對霍亂弧菌的抑制作用較好、且無溶血性,因此可以考慮將色胺酮與抗生素聯用,用于治療霍亂。色胺酮還能有效抑制各種肺結核菌株,其對結核分枝桿菌MTB H37Rv、M avium和M smegmatis具有較好的抑制活性,最小抑制濃度分別為1、4、6 mg/L,同時對多藥耐藥結核分枝桿菌菌株仍能保持較強抑制作用[53]。不僅如此,與空白組相比色胺酮(6.25~25 μg/mL)對革蘭氏陽性致齲菌和革蘭氏陰性厭氧牙周病菌均有顯著抑制作用,因此色胺酮還用于齲齒和牙周病的防治[54]。
色胺酮對許多真菌也表現出良好的抑制作用。隱球菌可從樹木和鳥類排泄物中分離出來,多通過散布在空氣中的繁殖體引起感染,并伴隨著誘發腦膜炎、肺炎等并發癥的風險。目前,隱球菌病是世界范圍內最嚴重的動物真菌病之一。一項關于中東地區隱球菌感染的流行病學調查發現,其在伊朗的樹木,鳥糞,動物和人類中的總體流行率分別估計為4.7%、20.4%、1.7%和2.8%[55]。研究表明在30 ℃時,鈣調神經磷酸酶抑制劑FK506和環孢霉素A誘導了色胺酮的抗隱球菌活性,最低抑菌濃度為2 μg/mL。值得注意的是,高溫環境中色胺酮仍能具有較強的抗隱球菌活性,具體表現在調節編碼細胞周期蛋白的基因和新生隱球菌孢子蟲的SBF/MBF復合物(CLN1、MBS1、PCL1和WHI5)的表達,誘導細胞周期停滯于G/S期。對新生隱球菌孢子蟲突變體數據庫的篩選進一步揭示了色胺酮與多種轉運蛋白和信號通路有關,如鈣轉運蛋白(Pmc1)和蛋白激酶a信號通路。提示色胺酮通過干擾細胞周期和信號傳導途徑對隱球菌產生了抗菌活性[56]。此外,色胺酮對犬小孢子菌、須毛癬菌、紅色癬菌、紫色癬菌、斷發癬菌、絮狀表皮癬菌、石膏樣癬菌、石膏樣小孢子菌、羊毛狀小孢子菌等皮膚類真菌也有較好的抑制效果[4]。
上述研究表明色胺酮具有活性強,抗菌譜廣的特點。色胺酮抗菌作用的發現為一些復雜致病菌的治療提供了新思路。可考慮單獨使用色胺酮,或將其與其它抗菌藥物聯合應用達到預防和治療細菌性疾病的目的。接下來還需對色胺酮的抗菌機制和科學合理使用進行深入研究,以期研發出以色胺酮為有效成分的抗菌新藥。
2.4 抗病毒作用
在短短20年間,冠狀病毒已經引發了包括COVID-19、嚴重急性呼吸系統綜合征冠狀病毒(SARS-CoV)、中東呼吸系統綜合征冠狀病毒(MERS-CoV)在內的3次傳染病大流行,給人類的健康帶來了極大的危害。盡管冠狀病毒具有極高的物種多樣性,但它們共享著關鍵的基因組元素,可依據病毒基因組信息和生物信息學特征指引發現潛在的有效藥物[57]。人冠狀病毒NL63(HCoV-NL63)是一種呼吸道病原體,感染后可引起上、下呼吸道感染,在兒童及免疫力低下的人群中的感染率較高。Tsai[58]等發現Strobilanthescusia(Nees) Kuntze葉的甲醇提取物能有效抑制HCoV-NL63感染細胞的細胞病變效應(CPE)和病毒產量,IC50為0.64 μg/mL,并確定其關鍵活性成分是色胺酮。在LCC-MK2和Calu-3細胞中,色胺酮表現出與細胞類型無關的抗HCoV-NL63活性,IC50分別為1.52 μmol/L和0.30 μmol/L。免疫熒光染色的感染性實驗結果表明,在兩種模式下,色胺酮阻止HCoV-NL63復制的早期(IC50=0.32 μmol/L)和晚期(IC50=0.06 μmol/L),特別是阻斷病毒RNA基因組的合成和類木瓜樣蛋白酶2的活性。此外,色胺酮(IC50=0.06 μmol/L)還表現出較強的殺病毒活性。值得注意的是,HCoV-NL63棘突蛋白(S蛋白)靶向ACE2受體,顯示出與SARS-CoV和SARS-CoV-2高度保守的序列和結構相似性,提示色胺酮有可能被作為一種生物活性劑來對抗SARS-CoV-2和其他人類冠狀病毒。3CLpro是冠狀病毒復制所必需的酶,也是SARS-CoV-2最具特征的藥物靶點之一,因此尋找以3CLpro為靶點的抑制劑對于COVID-19的防治具有重要意義[59]。Narkhede等[60]篩選出了包括甘草酸、色胺酮、大黃酸和小檗堿等在內的12種能發揮抗SARS-CoV-2感染作用的潛在候選藥物。分子對接結果顯示,色胺酮與3CLpro結合的親和力是-8.2 kcal/mol,通過與GLN 110形成兩個氫鍵以及與ILE 106、VAL 104的pi-alkyl烷基相互作用以及與ARG 105、GLN 107、THR 111、THR 292、PHE 294、ASP 153和SER 158的幾種范德華相互作用。它遵循利賓斯基的五定律,顯示出良好的藥物性質,沒有任何違反,提示色胺酮可用于抗COVID-19的探索性研究。
不僅如此,色胺酮對流感病毒也有潛在的預防作用。流感病毒分為甲、乙、丙三型,其中甲型流感病毒(IVA)最容易發生變異,流感大流行就是甲型流感病毒出現新亞型或舊亞型重現引起的。Xu等[61]對不同的IVA株,包括野生型和抗藥性IVA株,進行了細胞病變作用還原試驗(CEP)。意外地發現在A/Minfang/151/2000(野生型H3N2)病毒感染前給犬腎細胞注射色胺酮,其IC50=76.33±3.4 μmol/L與奧司他韋(IC50=54.45±14.24 μmol/L)和利巴韋林(IC50=62.45±16.24 μmol/L)相似,表明色胺酮對野生型IAV具有保護作用。神經氨酸酶(NA)是一種分布于流感病毒被膜上的糖蛋白,具有抗原性,能促進新生病毒的釋放。Zhang等[62]采用分子對接試驗研究了色胺酮對兩種流感病毒NA活性的影響(PDB密碼分別為3ti6和4mww)。發現色胺酮以顯著的結合能抑制NA(對3ti6編碼的結合能為-8.07 kcal/mol,對編碼4mww的NA的結合能為-7.52 kcal/mol),甚至優于奧司他韋(對3ti6編碼的結合能為-5.92 kcal/mol,對編碼4mww的NA的結合能為-7.12 kcal/mol),這表明色胺酮可有效地抑制NA,從而產生抗病毒作用。色胺酮可作為抗流感病毒的候選化合物,但還需進一步研究其體內功效、機制和結構轉化。
2.5 保護心臟
色胺酮可通過改善心肌缺血發揮保護心臟的作用。結扎冠狀動脈所致心肌缺血大鼠在灌胃色胺酮(0.10 g/kg)后,心肌收縮能力增強,心肌舒張順應性得到改善,左心室內收縮壓LVSP和左心室內壓最大變化速率±dp/dtmax明顯增高,與模型組比有明顯差異(P<0.05)。病理組織學檢測發現,給予色胺酮治療后心肌組織肌間隙纖維增生、慢性炎細胞浸潤和灶性壞死狀況均有一定改善,說明色胺酮對缺血心肌損傷有保護作用。但隨給藥劑量的增大,其對心肌的保護作用反而減弱,不呈量效依賴關系。這可能與色胺酮劑量增加,其對COX-2蛋白的抑制增強,導致副作用增大有關[63]。
2.6 抗血管生成
抗血管生成治療是一種通過血管生成抑制劑阻斷促血管生成因子,靶向抑制腫瘤血管生成的一種治療方法。血管生成因子眾多,常見的有血管內皮生長因子(VEGF)、表皮生長因子(EGF)、肝素、前列腺素E1/E2等。細胞試驗表明,色胺酮(50 μmol/L)能特異性作用于血管內皮細胞,抑制細胞增殖,這是通過下調內皮細胞中apelin的表達、抑制apelin啟動子活性和縮短mRNA半衰期來實現的[64]。色胺酮還能阻斷PKB或Akt和FAK途徑,使血管內皮細胞G2/M期阻滯,抑制血管內皮細胞的遷移和成管。在mRNA和蛋白質水平上,色胺酮(0~50 μmol/L)能呈劑量依賴性地降低細胞周期蛋白A、細胞周期蛋白B、細胞周期蛋白依賴激酶CDK1和CDK2的表達。值得注意的是,該研究是在原代血管內皮細胞用色胺酮預處理24 h后進行的,并且在沒有色胺酮的情況下進行遷移和成管實驗,提示色胺酮具有持續的抑制作用。
3 藥物動力學特征
藥代動力學性質是篩選新藥的重要指標之一,為給色胺酮的進一步研究和應用提供依據,學者們對色胺酮的藥代動力學特征進行了研究。Zhu等[65]對色胺酮(0.8~20 μmol/L)在Caco-2細胞中的吸收和轉運特性進行了考察,發現在該濃度范圍內色胺酮的吸收型Papp(AP→BL)和分泌型Papp(BL→AP)均無顯著變化,表明色胺酮的轉運不受濃度的影響;Papp(AP→BL)/Papp(BL→AP)大于1,表明色胺酮能被很好地吸收且以被動擴散為主;外排轉運蛋白P-gp和MRP2抑制劑的加入幾乎不影響色胺酮(4 μmol/L)的Papp,表明其外排不受P-gp和MRP2抑制劑的影響。
Zhang等[66]研究了色胺酮(80 mg/kg)在昆明小鼠體內的分布情況。小鼠灌胃2.5 h后,肝臟中色胺酮含量最高(3.54 g/g),其次是腎臟(2.12 g/g),肺(1.46 g/g),提示色胺酮與這些組織可能存在一定的關聯和靶向性;腦中含量最低(<0.2 g/g),提示色胺酮可能難以穿過小鼠血腦屏障。小鼠灌胃第12 h的Cmax是第1.5 h的Cmax的四十八分之一,表明色胺酮在體內無明顯蓄積。雄性SD大鼠體內初步藥代動力學研究表明,色胺酮(2 mg/kg BW i.v.)的半衰期為40.63±6.66 min,清除率為1.00±0.36 L/(h·kg)[67]。隨后,Wu等[68]研究了色胺酮在體內的藥代動力學過程。SD大鼠尾靜脈注射色胺酮(75 mg/kg)后,t1/2β為2.193 h,AUC(0-∞)為4.616 mg/(L·h),V1為33.794 L/kg,CL為29.76 L/(h·kg)。上述研究表明,色胺酮在體內具有吸收迅速、分布廣泛、代謝較快等特點,但還需進一步研究其作用靶點、代謝途徑,為色胺酮的臨床應用提供更多可靠的數據。
4 毒性
大量細胞毒性試驗表明,色胺酮不具直接毒性。Lee等[6]研究了色胺酮(2.5~20.0 μmol/L)對BV2小膠質細胞活性的影響,MTT結果顯示在沒有或存在LPS的情況下色胺酮均無細胞毒性。此外,顯微鏡下的形態學評估發現經色胺酮(2 μg/mL)處理的哺乳動物細胞系HEK-293細胞顯示出與PBS陰性對照組相似的紡錘形健康細胞,該結果進一步驗證了色胺酮非細胞毒性的特點[7]。急性毒性研究表明,小鼠灌胃色胺酮(25、50、80 mg/kg)后,體重略有減輕,但未見其他異常,無死亡。隨后,按同劑量連續給藥30 d,進行亞急性毒性研究。試驗期間動物的一般指標,如外觀、行為、對刺激的反應、分泌物排泄物等均無顯著異常,且未見小鼠死亡。小鼠臟器,如心、肝、脾、肺、腎、大腦等未見異常,臟器系數和空白對照組無顯著性差異(P>0.05)。結果提示,色胺酮毒性較低,安全性較好,是一種很有前途的替代療法藥物[8]。
5 總結
色胺酮的天然來源十分廣泛,在藥用植物馬藍、菘藍、蓼藍中均能分離得到色胺酮。現已證實色胺酮是一種后熟產物,且其合成過程是非酶催化的。色胺酮還是某些微生物如解脂假絲酵母、裂褶菌、海洋鏈霉菌的次生代謝產物。然而,自然界中色胺酮的含量很低,且存在提取工藝繁瑣、分離過程費時、提取率低等問題。因此色胺酮的化學合成是當前研究的熱點,靛紅和靛紅酸苷是常用的合成原料。與直接從植物中提取色胺酮相比,人工合成提高了色胺酮的產率,但合成路線較為繁瑣,反應條件較為苛刻,僅適用于實驗室少量合成。若想大規模生產,還需探索更簡捷、低成本的人工合成方法及工藝,色胺酮的仿生合成可作為未來的研究方向。
眾多藥理實驗表明,色胺酮具有良好的抗炎、抗菌、抗腫瘤、抗病毒等生物活性,且毒副作用低。色胺酮具有的藥理作用及在腸道中易于被吸收的特點使其在疾病的預防與治療方面具有廣闊的應用前景。然而,現今有關色胺酮的報道僅限于體外實驗或動物實驗,具有局限性,未來還需足夠的臨床試驗數據作為有力支撐,如何充分利用、開發具有市場價值的色胺酮類化合物產品,將會是今后的研究重點。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
東方少年·布老虎畫刊(2023年8期)2023-08-01 15:45:12
科學大眾(2021年6期)2021-07-20 07:42:44
科學(2020年3期)2020-11-26 08:18:30
學苑創造·A版(2020年9期)2020-10-13 09:41:02
娃娃樂園·綜合智能(2019年3期)2019-04-03 09:17:36
中成藥(2018年2期)2018-05-09 07:19:34
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
湖北師范大學學報(自然科學版)(2015年2期)2016-01-10 08:41:55
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
