縮骨鯽Cyt b基因和控制區序列克隆及系統進化分析
2023-12-02 12:10:47李強鄭鵬黃文煒彭蘇漢周江偉詹華偉藍昭軍桂林
湖南農業科學 2023年10期
李強 鄭鵬 黃文煒 彭蘇漢 周江偉 詹華偉 藍昭軍 桂林


摘 要:研究通過Sanger測序法對天然雌核發育的縮骨鯽(Carassius auratus var. Suogu)線粒體DNA的Cyt b基因和控制區進行擴增和測序,并與已發表的其他鯽屬魚類的Cyt b基因進行系統進化分析,探討縮骨鯽與其他鯽屬魚類的系統進化關系。結果表明:縮骨鯽線粒體DNA中Cyt b基因和控制區全序列分別長1 141和924 bp;其中,Cyt b基因的堿基含量為29.1%(T)、27.9%(C)、28.7%(A)和14.3%(G),控制區序列的堿基含量為32.6%(T)、20.7%(C)、32.6%(A) 和14.1%(G);2種序列中G+C含量均明顯低于A+T含量,且G含量偏低,顯示了與其他脊椎動物線粒體核苷酸堿基含量相似的特征;控制區序列可以分為終止序列區、中央保守區和保守序列區3個區域,均識別到了對應的保守序列;基于鄰接法(NJ)和最大簡約法(MP)構建縮骨鯽與其他鯽屬魚類Cyt b 基因的系統進化樹表明,縮骨鯽與紅鯽和淇河鯽的親緣關系最近,而縮骨鯽與野鯽、淇河鯽和紅鯽分布在同一個姐妹支系,進一步說明縮骨鯽可能起源于野鯽,是野鯽的一個地方種群。
關鍵詞:縮骨鯽;細胞色素b基因;控制區;系統進化分析
中圖分類號:Q953.3; S917.4 文獻標識碼:A文章編號:1006-060X(2023)10-0007-04
Sequence Cloning and Phylogenetic Analysis of Cyt b Gene and Control Region in mtDNA of Carassius auratus var. Suogu
LI Qiang1, ZHENG Peng1, HUANG Wen-wei1, PENG Su-han1, ZHOU Jiang-wei1, ZHAN Hua-wei1, LAN Zhao-jun2, GUI Lin1
(1. College of Life Science, Guangzhou University, Guangzhou 510006, PRC; 2. Shaoguan Fishery Technology Promotion Station, Shaoguan Fisheries Research Institute, Shaoguan 512006, PRC)
Abstract:The complete sequences of Cyt b gene and control region in mtDNA of natural gynogenesis triploid C. auratus var. Suogu were determined through Sanger sequencing method after PCR amplification. The phylogenetic analysis was performed by sequence alignment of the Cyt b gene between C. auratus var. Suogu and the other Carassius fishes in GenBank to explore their phylogenetic relationship. Results showed that the total length of Cyt b gene was 1 141 bp and the control region was 924 bp. The base composition of Cyt b gene was 29.1% T, 27.9% C, 28.7% A and 14.3% G, and control region was 32.6% T, 20.7% C, 32.6% A and 14.1% G. In these two sequences, the contents of G+C (42.2%, 34.8%) were significantly lower than those of A+T (57.8%, 65.2%), and the G base contents were low, which showed a similarity to those of other vertebrates. The sequence of control region was divided into three regions: termination sequence region, central conserved region and conserved sequence region, in which the corresponding conserved sequences were identified. The phylogenetic trees constructed by the neighbor-joining method (NJ) and maximum parsimony (MP) based on Cyt b gene in C. auratus var. Suogu and other Carassius fishes have revealed that C. auratus var. Suogu is closely related to C. auratus red var. and C. auratus var. Qihe. In addition, the C. auratus var. Suogu is distributed in the same sister branch with C. auratus auratus, C. auratus var. Qihe and C. auratus red var., which has indicated that the C. auratus var. Suogu may be originated from C. auratus auratus and belongs to a local population of C. auratus auratus.
Key words:Carassius auratus var. Suogu; cytochrome b gene; control region; phylogenetic analysis
收稿日期:2023–05–08
基金項目:廣州市科技計劃項目(201804010486);廣東省教育科學規劃課題(2021GXJK097)
作者簡介:李 強(1983—),男,廣東韶關市人,實驗師,主要從事淡水魚類生態與親緣地理研究。
通信作者:桂 林
縮骨鯽(Carassius auratus var. Suogu)是一種天然雌性核發育的三倍體鯽魚,屬鯉形目鯉科鯽屬,主要分布在廣東省韶關地區,為該地方特有的鯽魚品種[1]。與其他鯽魚形態特征相比,縮骨鯽的尾柄長明顯小于尾柄高,體高明顯比其他鯽魚品種突出[2],這一形態是漫長的環境適應進化的結果,為可遺傳性狀。由于縮骨鯽體短而高的特點,其含肉比例較高,且口感鮮美,是產區的重要經濟魚類。近年來,由于環境污染和過度捕撈等人為干涉,縮骨鯽的野生資源銳減,亟待加以保護和可持續開發利用。目前有關縮骨鯽的研究主要集中在生物學特性[1-2]、人工繁殖[3]、基因克隆[4-5]等方面,尚無縮骨鯽系統進化關系的研究報道。
線粒體DNA(mtDNA)具有結構簡單、無基因重組、進化速度比核基因快等特點,是研究魚類系統進化和遺傳多樣性的理想分子標記[6]。其中,細胞色素b基因(Cyt b)是mtDNA中結構和功能被研究得最為廣泛的基因之一,該基因進化速度適中,較適合研究種內和種間遺傳變異[7];另外,控制區是mtDNA中主要的非編碼區,屬于進化最快的片段,常作為分子標記來探討各種魚類的種群遺傳多樣性和種內遺傳分化[8]。該研究通過對縮骨鯽線粒體的Cyt b基因和控制區進行擴增和測序,與已發表的其他鯽屬魚類進行系統進化分析,探討縮骨鯽與其他鯽屬魚類的系統進化關系,旨在為縮骨鯽種質資源保護、研究和利用提供基礎數據。
1 材料與方法
1.1 試驗材料
試驗所用縮骨鯽來自廣東省韶關市水產研究所的試驗基地。隨機選取5尾縮骨鯽活魚樣品,樣品形態見圖1。
1.2 DNA 提取和PCR擴增
采用動物基因組DNA提取試劑盒提取縮骨鯽總DNA。PCR擴增反應體系(25 μL):PCR Master Mix 12.5 μL、Primer1 1μL、Primer2 1μL、模板DNA 3 μL、滅菌雙蒸水7.5 μL。反應在PTC100或PTC200型PCR儀上進行。PCR擴增程序: 95℃預變性3 min;95℃變性0.5 min,53℃退火0.5 min ,72℃延伸1.5 min ,循環35次;72℃ 延伸10 min。Cyt b基因克隆的引物為L14724和H15915[9],控制區序列克隆的引物為CR1和CR2[10],引物由生工生物工程(上海)股份有限公司合成。電泳檢驗合格的PCR產物委托生工生物工程(上海)股份有限公司進行純化并雙向測序。
1.3 數據分析
對測得的序列使用Sequence Analysis 5.2 軟件進行拼接。通過MEGA 7.0 軟件分析各序列的結構特征[11]。為了探討縮骨鯽在鯽屬魚類中的系統進化關系,從GenBank 數據庫下載鯽屬魚類的Cyt b 基因和控制區全序列,包括野鯽(C. auratus auratus)、淇河鯽(C. auratus var. Qihe)、彭澤鯽(C. auratus var. Pengze)、滁州鯽(C. auratus var. Chuzhou)、方正銀鯽(C. auratus gibelio)、方正銀鯽A系(C. auratus gibelio clone A)、方正銀鯽D系(C. auratus gibelio clone D)、方正銀鯽E系(C. auratus gibelio clone E)、蘭氏鯽(C. auratus langsdorfi)、日本白鯽(C. cuvierius)、萍鄉肉紅鯽(C. auratus var. Pingxiangnensis)、紅鯽(C. auratus red var.)和鯉魚(Cyprinus carpio),Genbank中的序列登錄號依次為JN105355、KJ476998、KJ874429、KM659025、GU138989、JF496198、JF496197、KX611160、AB006953、AB045144、HQ875340、KJ874430和AP009047。基于Cyt b 基因構建鄰接法(NJ)和最大簡約法(MP)的鯽屬魚類的分子系統進化樹。
2 結果與分析
2.1 線粒體DNA Cyt b基因和控制區序列分析
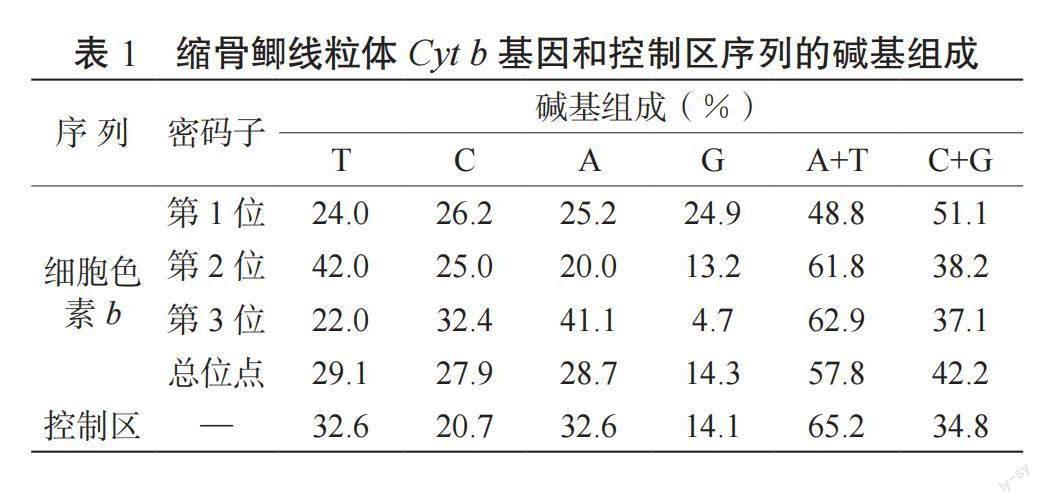
縮骨鯽線粒體Cyt b 基因和控制區序列經軟件拼接后的長度分別為1 141和923? bp。Cyt b 基因的起始密碼子是ATG,終止密碼子是T,共編碼380個氨基酸。Cyt b? 基因中T、C、A 和G 的含量分別為29.1%、27.9%、28.7%和14.3%,A+T含量(57.8%)明顯大于G+C含量(42.2%);控制區序列的T、C、A和G含量分別為32.6%、20.7%、32.6%和14.1%,A+T含量(65.2%)明顯大于G+C含量(34.8%)。縮骨鯽線粒體Cyt b 基因和控制區序列中,G含量顯著低于其他3種堿基,且具有明顯的AT偏好性。
2.2 線粒體控制區序列結構
縮骨鯽和其他硬骨魚類的控制區相似,位于tRNAPro和tRNAPhe之間,全長923 bp。通過與其他魚類控制區及各個區域關鍵序列和保守序列的比對,縮骨鯽的控制區也可以劃分為3個部分:終止序列區、中央保守區和保守序列區。由圖2可知,終止序列區位于控制區的5'端,是控制區變異最大的區域,識別出1個終止相關序列(TAS),其堿基序列為TACATATATGTA;中央保守區識別出了CSB-F(ATGTAGTAAGAGACCACC)、CSB-E(ATGATA GAATCAGGGACAT)和CSB-D(TTATACTGGCATCTGA TTAA)等關鍵序列;保守序列區識別出了CSB-1(ATATAAATGAATTATCGTAAGACATA)、CBS-2(CAAACCCCCCTACCCCC)和CBS-3(TGTCAAACCCC GAAACCA)等保守序列(圖2)。
2.3 基于鄰接法(NJ)和最大簡約法(MP)的縮骨鯽系統進化樹
基于鄰接法(NJ)和最大簡約法(MP)構建縮骨鯽與其他鯽屬魚類Cyt b 基因的系統進化樹,以鯉魚作為外類群,結果如圖3所示。2種方法構建的系統進化樹的拓撲結構相似,鯽屬魚類聚成Ⅰ和Ⅱ 2支。來自中國的鯽屬魚類組成了第Ⅰ支,而來自日本的蘭氏鯽和日本白鯽組成了第Ⅱ支,支系Ⅰ和Ⅱ之間的遺傳距離為6.7%,這說明蘭氏鯽和日本白鯽與中國的鯽屬魚類發生了明顯的遺傳分化。在Ⅰ支中,又可分為A和B個2姐妹群。A群包含了方正銀鯽系列、彭澤鯽、滁州鯽和萍鄉肉紅鯽;B群則包含野鯽、縮骨鯽、淇河鯽、紅鯽等物種,A群和B群之間的遺傳距離為1.8%,中國鯽屬魚類的遺傳分化程度較小。從B支系可知,縮骨鯽與淇河鯽和紅鯽聚成一支,這說明縮骨鯽與淇河鯽和紅鯽的親緣關系較近。
3 討 論
3.1 序列組成及控制區結構
魚類線粒體DNA是研究魚類系統發育和遺傳多樣性的理想分子標記[12]。目前,研究最多是Cyt b基因、COI基因、ATPase8/6基因和控制區序列。該研究首次獲得了縮骨鯽的線粒體Cyt b基因和控制區的全序列,它們的長度分別為1 141和923 bp。縮骨鯽這2種序列的堿基A+T含量均高于G+C,G含量顯著低于其他3種堿基,這些都與脊椎動物線粒體DNA的特點相一致[13]。Cyt b基因以單獨的T堿基作為終止密碼子,在其他動物類群中也常見[14]。一般認為不完全終止密碼子是由于蛋白質編碼基因3'端與tRNA基因重疊所致[15],不完全的終止密碼子將在轉錄過程中由mRNA 3'端的 Poly(A)尾補全[16]。控制區是脊椎動物線粒體基因組中主要的非編碼片段,是變異最大的區域[6]。縮骨鯽線粒體基因組控制區的長度為923 bp,在終止序列區識別出1個終止相關序列(TAS),中央保守區識別出了CSB-F、CSB-E和CSB-D等關鍵序列,保守序列區識別出了CSB-1、CBS-2和CBS-3等保守序列,這與劉良國等[10]報道的其他鯽屬魚類線粒體控制區結構相似。
3.2 縮骨鯽系統地位
我國的鯽屬魚類包括鯽和黑鯽2種,前者又可以分為鯽指名亞種和銀鯽亞種,銀鯽亞種主要分布在黑龍江和額爾齊斯河河水系,而鯽幾乎遍布全國各主要水系[17]。鯽適應性強、是遺傳背景復雜的類群,在長期的生態適應過程中形成了諸如方正銀鯽、滇池高背鯽、江西彭澤鯽、河南淇河鯽、貴州普安鯽、安徽滁州鯽、廣東縮骨鯽等許多不同的鯽種群[18]。廣東縮骨鯽主要分布在廣東省韶關地區的翁源縣等地,謝楠等[19]將其歸為銀鯽的地方類群。張輝等[20]用限制性內切酶對銀鯽、彭澤鯽、縮骨鯽和野鯽的mtDNA進行單倍型分析認為,縮骨鯽屬于野鯽的一個地方品系。而劉良國等[21]對廣東地區彭澤鯽、縮骨鯽和野鯽進行RAPD分析,結果表明彭澤鯽和縮骨鯽的親緣關系較近,推測它們可能共同起源于野鯽。
該研究使用NJ和MP 2種方法構建的鯽屬魚類Cyt b基因系統進化樹的拓撲結構基本一致。在系統進化樹上,方正銀鯽系列、彭澤鯽、滁州鯽、和萍鄉肉紅鯽聚成一支;野鯽、縮骨鯽、淇河鯽和紅鯽聚成另一支,這2支相互形成姐妹群,具有很高的支持率,分別代表了銀鯽類群和非銀鯽類群。這一結果也表明,縮骨鯽與紅鯽、淇河鯽和野鯽的親緣關系較近,與方正銀鯽、滁州鯽、彭澤鯽的親緣關系較遠,縮骨鯽可能起源野鯽,屬于野鯽的一個地方種群,而非銀鯽地方種群,這與張輝等[20]的研究結果相似。
參考文獻:
[1] 俞豪祥,張海明,林蓮英. 廣東雌核發育鯽魚的生物學及養殖試驗的初步研究[J]. 水生生物學報,1987,11(3): 287-288.
[2] 俞豪祥,張海明. “縮骨鯽”的生物學及養殖試驗初報[J]. 水產科技情報,1986,13(3): 21.
[3] 吳土金,鐘良明,谷平華,等. 縮骨鯽人工繁育技術研究[J]. 海洋與漁業,2016(9): 54-55.
[4] 楊 揚,許炎坤,舒 琥,等. 縮骨鯽MSTN基因的克隆與組織表達分析[J]. 淡水漁業,2017,47(2): 23-29.
[5] 楊 揚,周惠強,舒 琥,等. 縮骨鯽(Carassius auratus sogu var.)MC4R克隆、組織分布以及與生長相關SNPs的篩選[J]. 海洋與湖沼,2017,48(4): 830-837.
[6] 劉云國. 水產生物DNA分子標記技術[M]. 北京: 科學出版社,2009.
[7] 肖武漢,張亞平. 魚類線粒體DNA的遺傳與進化[J]. 水生生物學報,2000,24(4): 384-391.
[8] BEHEREGARAY L B,SUNNUCKS P. Fine-scale genetic structure,estuarine colonization and incipient speciation in the marine silverside fish Odontesthes argentinensis[J]. Molecular Ecology,2001,10(12): 2849-2866.
[9] XIAO W H,ZHANG Y P,LIU H Z. Molecular systematics of Xenocyprinae(Teleostei: Cyprinidae):taxonomy,biogeography,and coevolution of a special group restricted in East Asia[J]. Molecular Phylogenetics and Evolution,2001,18(2): 163-173.
[10] 劉良國,楊品紅,王文彬,等. 洞庭青鯽與其他六個鯽魚品系線粒體DNA控制區的比較分析[J]. 水生生物學報,2010,34(2): 378-387.
[11] KUMAR S,STECHER G,TAMURA K. MEGA7:molecular evolutionary genetics analysis version 7.0 for bigger datasets[J]. Molecular Biology and Evolution,2016,33(7): 1870-1874.
[12] 郭新紅,劉少軍,劉 巧,等. 魚類線粒體DNA研究新進展[J]. 遺傳學報,2004,31(9): 983-1000.
[13] HOCHACHKA P,MOMMSEN T P. Biochemistry and molecular biology of fish[M]. London: Elsevier Science Publishers,1993.
[14] CUI Z X,LIU Y,LI C P,et al. The complete mitochondrial genome of the large yellow croaker,Larimichthys crocea(Perciformes,Sciaenidae):unusual features of its control region and the phylogenetic position of the Sciaenidae[J]. Gene,2009,432(1/2): 33-43.
[15] WOLSTENHOLME D R. Genetic novelties in mitochondrial genomes of multicellular animals[J]. Current Opinion in Genetics & Development,1992,2(6): 918-925.
[16] OJALA D,MERKEL C,GELFAND R,et al. The tRNA genes punctuate the reading of genetic information in human mitochondrial DNA[J]. Cell,1980,22(2): 393-403.
[17] 中國科學院中國動物志編輯委員會主編. 中國動物志硬骨魚綱鯉形目(下卷)[M]. 北京: 科學出版社,2000.
[18] 汪留全,胡 王. 我國鯽品種(系)資源及其生產性能的初步分析[J]. 水利漁業,1997,18(5): 4-6.
[19] 謝 楠,劉 凱,馮曉宇. 鯽魚常見品種概述[J]. 杭州農業與科技,2016(4): 13-17.
[20] 張 輝,董新紅,葉玉珍,等. 三個三倍體鯽魚品系及野鯽mtDNA的比較研究[J]. 遺傳學報,1998,25(4): 330-336.
[21] 劉良國,趙 俊,陳湘粦. 彭澤鯽兩個雌核發育克隆與三個鯽魚品系的RAPD分析[J]. 淡水漁業,2005,35(2): 13-16.
(責任編輯:成 平)
