短期圍封和放牧條件下施氮對橐吾型退化草地植被的影響
2023-12-04 06:48:18張千山王玉琴宋梅玲余錢雯王宏生
草地學報 2023年11期
張千山, 王玉琴,2*, 宋梅玲,2, 周 睿,2, 余錢雯, 王宏生,2*
(1. 青海大學畜牧獸醫科學院/青海省畜牧獸醫科學院, 青海 西寧 810016;2. 省部共建三江源生態與高原農牧業國家重點實驗室, 青海大學, 青海 西寧 810016)
我國是草地資源大國,天然草地面積占我國土地面積的40%[1]。豐富的草地資源具有防風、固沙、氣候調節等重要作用,是維護我國生態系統安全的重要基石,但由于全球環境變化和人類活動的干擾,我國天然草地的面積正在不斷縮小,草地在加速退化,且已經嚴重影響了當地居民的正常生產生活[2-3]。目前,普遍認為長期過度放牧是導致草原植被大面積退化的主要原因[4-5],而繼荒漠化后,毒雜草的大肆蔓延,也成為了草地退化的主要表現形式之一,毒草治理已經成為了青藏高原退化草地恢復治理中的重要一環[6-7]。氮添加是改善草地營養物質平衡的重要管理措施,可以有效補充草地中的營養元素,緩解植物間的競爭壓力,有研究表明,施用30~40 kg·hm-2氮素可以有效促進退化草地的恢復及可持續利用[8],而圍封可以避免外界放牧干擾,使退化草場的植物得以一定程度上恢復繁衍,所以兩種措施常常結合使用以加快退化草地的治理和恢復[9-10]。
黃帚橐吾(Ligulariavirgaurea)是青藏高原上一種危害嚴重的毒草,常作為高寒草甸退化的指示物種,其大面積滋生蔓延已經嚴重危害了高寒草甸的健康發展[11]。目前的研究表明兩個重要機制影響了黃帚橐吾的快速擴散:一是黃帚橐吾體內含有一種具有特殊氣味的化感物質,可以抑制其它植物的生長,提高自身競爭能力[12];二是黃帚橐吾分布地土壤微生物功能多樣性的改變所引起的土壤氮素限制[13]。但目前關于氮素添加在黃帚橐吾型退化草地中是否對草地恢復有效果卻少有報道。因此,本研究以黃帚橐吾型退化草地為研究對象,在放牧和圍封條件下添加氮素,比較分析采取措施2年后草地群落組成、植被多樣性、地上和地下生物量以及黃帚橐吾密度的變化規律,明確氮素添加對黃帚橐吾型退化草地植被的影響,以期為黃帚橐吾型退化草地的恢復治理提供理論依據。
1 材料與方法
1.1 樣地概況
試驗點位于青海省黃南藏族自治州河南縣(34°44′18″ N,10°136′31″ E),平均海拔約3 589 m。該地屬典型高原大陸性氣候,年平均氣溫2.0℃,年均降水量579~616 mm,植物生長期為120~140 d。試驗地主要物種為黃帚橐吾、線葉嵩草(Kobresiacapillifolia)、矮生嵩草(Kobresiahumilis)、高原早熟禾(Poapratensis)、垂穗披堿草(Elymusnutans)等,伴生雜草主要為秦艽(Gentianastraminea)、蒲公英(Taraxacummongolicum)、莓葉委陵菜(Potentilla)。黃帚橐吾平均密度為133.6 株·m-2。
1.2 樣地設置
在研究區域內選取地勢較為平坦、植被分布均勻、黃帚橐吾為害嚴重的草地為試驗樣地。于2019年5月采用網圍欄建立平行生長季禁牧試驗區和放牧試驗區各3個,面積均為80 m×40 m。在各試驗區內隨機設置3個面積為17 m×10 m的小區進行添加氮素試驗,添加氮素量為40 kg·hm-2,氮素于2019年6月一次性施入,添加氮素選擇陰天將尿素(N含量46%)溶于水中在各小區內人工噴灑,并在不施氮素小區噴施等量清水。以不添加氮素作為對照,共4個處理,分別為放牧(G)、放牧+氮素(G+N)、圍封(E)、圍封+氮素(E+N)。
1.3 植被調查
分別于2020和2021年8月進行草地植被群落調查,在每個小區內隨機選取3個0.5 m×0.5 m的樣方,記錄樣方內所有植物物種名,用直尺量取每種植物的高度計算均值,以針刺法測定其蓋度,并統計每個樣方內黃帚橐吾的個體數。采集樣方框內所有地上植物組織,按物種分裝于信封袋中,帶回實驗室后低溫烘干,稱取烘干后的植物組織重量,計算每種處理下各物種的地上生物量以及總地上生物量[6]。在樣方內用直徑為7.0 cm的根鉆分別采集0~10 cm和10~20 cm土層的土壤,裝網兜后用清水反復漂洗,確保去除根系附著的雜質,經65℃烘干48 h,進行地下生物量的測定。
1.4 計算方法
按照測得的樣方數據(高度、蓋度、地上生物量)計算草地植物群落的多樣性[14]:
用Shannon-Wiener指數(H′)計算草地植物群落物種多樣性指數:
用Simpson指數(D)計算草地植物群落物種優勢度指數:
用Pielou指數(E)計算草地植物群落均勻度指數:
物種重要值(Ni)=(相對高度+相對蓋度+相對生物量)/3
物種豐富度指數(S)=物種數
式中,N為重要值總和,Ni為樣方內第i物種的重要值,Pi=Ni/N,Pi為物種i的相對重要值。
1.5 數據分析
使用Excel 2020對數據進行錄入、整理和作圖,運用SPSS 22.0軟件對相同年份不同處理間的植被群落多樣性指數、植物地上生物量和相同土層地下生物量等進行單因素方差分析,并進行重復度量方差分析方法進行檢驗(95%水平),圖表數據均采用平均值±標準誤表示。利用R Development CoreTeam,對所有指標采用PCA分析,根據各因子在PCA軸上載荷的大小判斷其作用大小,根據其夾角判斷其相關性,并根據主成分累積貢獻率,選擇重要主成分,利用得分公式求出不同處理草地植被綜合分值并排序。
2 結果與分析
2.1 添加氮素對草地群落重要值的影響
按功能群及重要程度分為以下四類(表1),莎草科、禾本科、豆科、雜類草(如黃帚橐吾、莓葉委陵菜、美麗風毛菊等),其中各處理間的主要物種組成基本相同,不同年份和處理間的優勢種群則有所差異。草地圍封后禾本科植物重要值普遍提升,其它各科植物重要值則有所下降,其中黃帚橐吾重要值下降較為明顯并于圍封第2年變為亞優勢物種,重要值<10%。在放牧草地添加氮素后,第1年的黃帚橐吾重要值明顯降低,但仍為優勢物種,其重要值>10%,而高原早熟禾重要值則明顯提高,但重要值仍低于黃帚橐吾;第2年,黃帚橐吾從優勢種變為亞優勢種,重要值>5%,而高原早熟禾重要值明顯提高成為優勢種,重要值>10%。在圍封草地添加氮素后,第1年的黃帚橐吾重要值有所上升,且高原早熟禾從優勢種變為亞優勢物種;第2年,圍封草地和圍封加氮素草地的黃帚橐吾均變為亞優勢物種,垂穗披堿草則成為唯一優勢物種,重要值>10%,且圍封加氮素草地的垂穗披堿草重要值高于圍封草地,其余物種均未發生優勢種的替換。相較于放牧加氮素草地,圍封加氮素草地的禾本科植物重要值提升更為明顯。
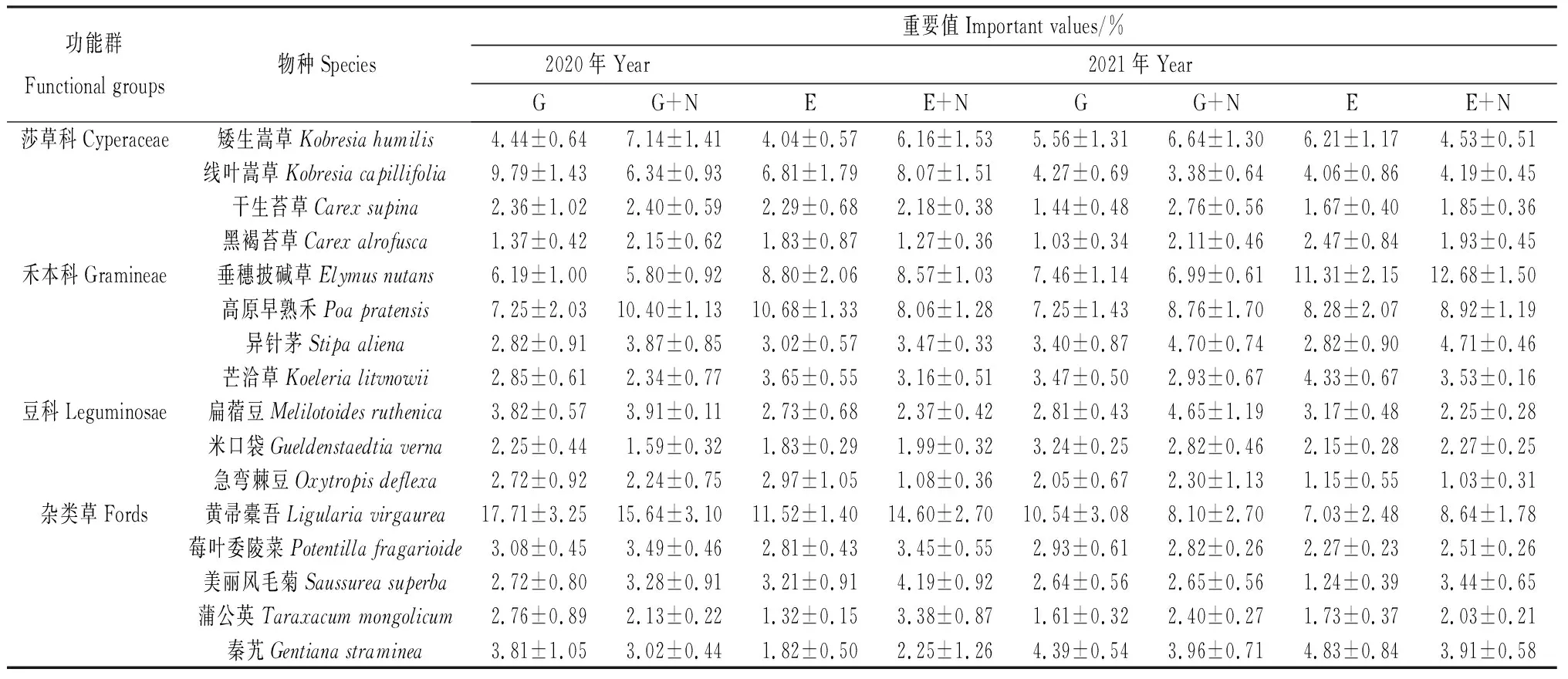
表1 添加氮素后草地群落主要物種組成及其重要值Table 1 Species composition and important value of grassland community after nitrogen addition
2.2 添加氮素對草地群落多樣性的影響
不同處理下退化草地豐富度指數、均勻度指數、優勢度指數和香農威納指數表現為(圖1):在放牧草地和圍封草地添加氮素1年后,圍封加氮素草地的香農威納指數和豐富度指數均為最高,圍封草地的均勻度指數和優勢度最高,放牧草地的各多樣性指數均為最低;在放牧草地和圍封草地添加氮素2年后,放牧草地的多樣性指數均為最高,圍封草地的香農威納指數和豐富度指數最低,但各處理的多樣性指數在處理間及年份間差異均不顯著,且處理和年份之間也無顯著交互作用(表2)。
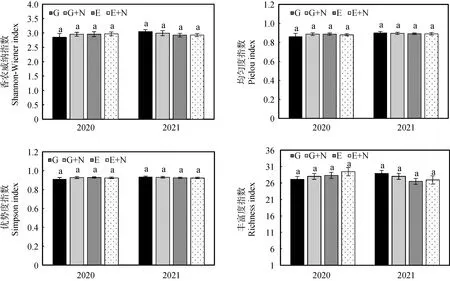
圖1 添加氮素后植物群落物種多樣性的變化Fig.1 Change of diversity indexes of grassland communities after nitrogen addition注:不同小寫字母表示同一年份不同處理間差異顯著(P<0.05)。下同Notes:Different lowercase letters indicate a significant difference among different treatments in the same year at the 0.05 level. The same as below
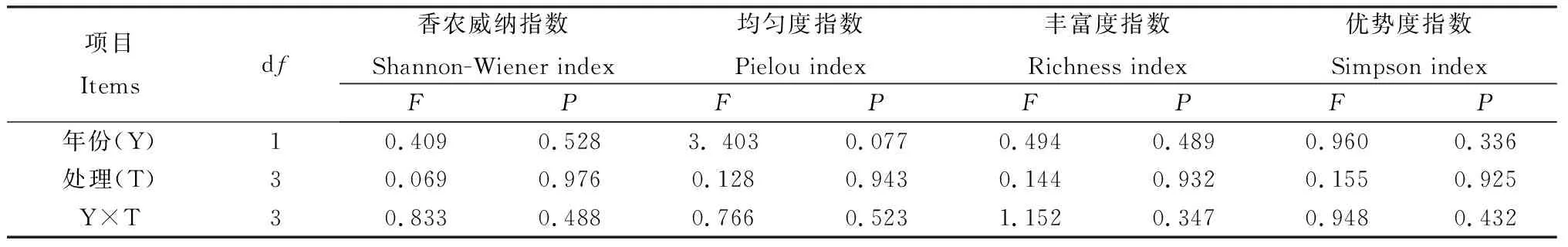
表2 添加氮素后不同年份植物群落物種多樣性的的重復度量方差分析結果Table 2 Results of repeated measures ANOVA on the effects of nitrogen addition on the grassland community diversity in different years
2.3 添加氮素對黃帚橐吾密度和生物量的影響
在放牧草地和圍封草地添加氮素對黃帚橐吾密度均無顯著影響。但在圍封草地添加氮素后,黃帚橐吾的密度和生物量均低于放牧草地添加氮素處理,且黃帚橐吾密度顯著低于放牧條件下添加氮素處理(P<0.05,圖2)。年份對黃帚橐吾密度具有顯著影響(P<0.05),但年份和處理間之間無顯著交互作用(表3)。
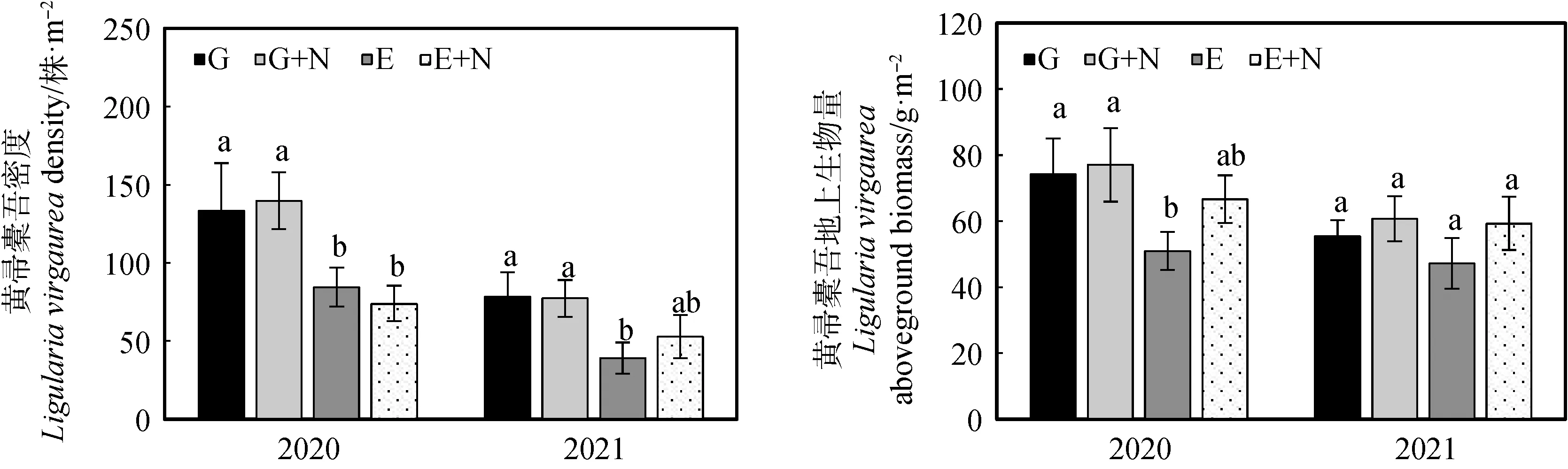
圖2 添加氮素后黃帚橐吾密度和地上生物量的變化Fig.2 Density and aboveground biomass of Ligularia virgaurea after nitrogen addition

表3 添加氮素后不同年份黃帚橐吾密度和地上生物量的重復度量方差分析結果Table 3 Results of repeated measures ANOVA on the effects of nitrogen addition on the density and aboveground biomass of L.virgaurea in different years
2.4 添加氮素對各功能群地上生物量的影響
比較各處理地上生物量得到(圖3),在放牧草地添加氮素后,莎草科植物地上生物量第2年顯著提高(P<0.05);禾本科植物地上生物量2年內均無顯著變化;豆科植物地上生物量第2年顯著降低(P<0.05);雜類草無顯著變化。在圍封草地添加氮素后,莎草科植物地上生物量在第2年顯著降低(P<0.05);禾本科植物地上生物量在第2年顯著提高(P<0.05);豆科植物地上生物量明顯降低但差異不顯著;雜類草地上生物量無明顯變化。相較于放牧草地,在圍封草地添加氮素后,莎草科植物地上生物量第1年明顯提高,但差異不顯著,第2年顯著降低(P<0.05);禾本科植物地上生物量第2年顯著提高(P<0.05);豆科植物地上生物量2年內均明顯降低,第1年顯著降低(P<0.05);雜類草地上生物量無明顯變化。另外年份對總地上生物量和禾本科、豆科、雜類草地上生物量均產生了顯著影響(P<0.05),不同處理對禾本科和豆科地上生物量產生了顯著影響(P<0.05),年份和處理間僅對莎草科地上生物量有顯著交互作用(P<0.05)(表4)。
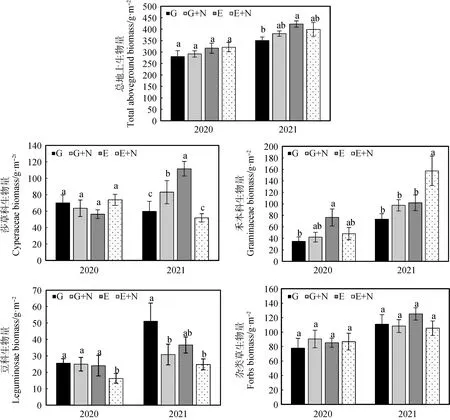
圖3 添加氮素后草地群落地上生物量的變化Fig.3 Effects of nitrogen addition on aboveground biomass of grassland communities
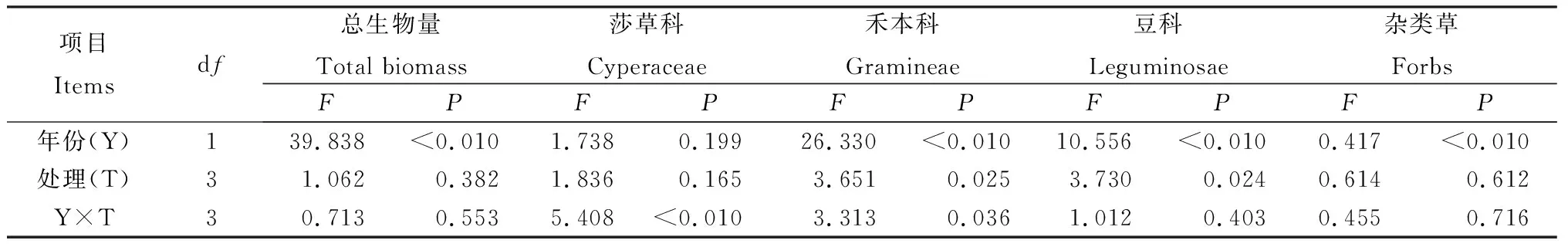
表4 添加氮素后不同年份草地地上生物量的重復度量方差分析結果Table 4 Results of repeated measures ANOVA on the effects of nitrogen addition on the aboveground biomass of the community and the four groups in different years
2.5 添加氮素對草地地下生物量的影響
在圍封草地添加氮素后,草地總地下生物量在第1年顯著提高(P<0.05),而在放牧草地添加氮素后草地總地下生物量無顯著變化。氮素添加對不同土層的影響不同,在0~10 cm土層中,放牧草地添加氮素后地下生物量在兩年內均無顯著變化;在圍封草地添加氮素后,第1年地下生物量顯著提高(P<0.05)。而在10~20 cm土層中,地下生物量表現為,在放牧條件下添加氮素第1年后地下生物量有降低趨勢,第2年無變化,在圍封條件下添加氮素,第1年地下生物量明顯增加,第2年則無明顯差異(圖4)。相較于放牧條件,在圍封條件下添加氮素后草地總地下生物量和0~10 cm地下生物量均顯著提高(P<0.05)。另外年份和各處理對地下生物量均無顯著影響,年份和處理間也無顯著交互作用(表5)。
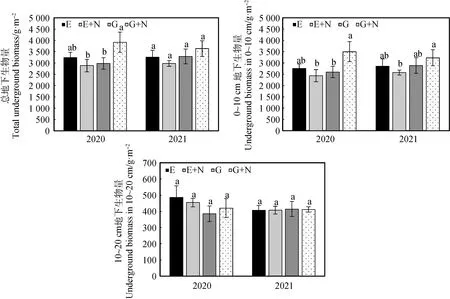
圖4 添加氮素后各土層的地下生物量的變化Fig.4 Underground biomass in different soil layers after nitrogen addition

表5 添加氮素后不同年份地下生物量的重復度量方差分析結果Table 5 Results of repeated measures ANOVA on the effects of nitrogen addition on underground biomass in different years
2.6 添加氮素后黃帚橐吾型退化草地植被的PCA分析
主成分分析表明(圖5):2020年PCA軸的累積解釋方差比例達46.6%,第一主成分累積貢獻率為26.5%,第二主成分累積貢獻率為20.1%。2021年PCA軸的累積解釋方差比例達56.1%,第一主成分累積貢獻率為34.3%,第二主成分累積貢獻率為21.8%。通過計算主成分綜合得分,并按照F值大小對個處理進行排序得到(表6),2020年、2021年綜合得分最高處理均是圍封,2020年排列順序為圍封>放牧+氮素>圍封+氮素>放牧;2021年排列順序為圍封>圍封+氮素>放牧+氮素>放牧。
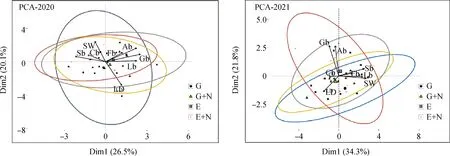
圖5 草地植被的PCA分析Biplot圖Fig.5 Biplot diagram of PCA analysis on grassland vegetation注:SW為香農威納指數;LD為黃帚橐吾密度;Ab為總地上生物量;Cb為莎草科生物量;Gb為禾本科生物量;Lb為豆科生物量;Fb為雜草生物量;Sb為總地下生物量注:SW, Shannon-Wiener index;LD, Ligularia virgaurea density;Ab, total above-ground biomass;Cb, cyperaceae biomass;Gb, graminaceae biomass;Lb, leguminosae biomass;Fb, forbs biomass;Sb, total subsurface biomass

表6 不同處理的主成分綜合得分及其排序Table 6 Scores and ranking of the principal component of different treatments
3 討論
3.1 添加氮素對草地主要物種的影響
重要值是物種在群落中地位和作用的綜合數量指標,用來表現不同植物的相對重要性,對研究群落演替具有重要意義[15]。而氮添加和圍封一直以來都作為恢復草地植被、減少毒雜草在草地所占比例的重要措施。本研究顯示,草地圍封后,禾本科植物的重要值普遍提高,在向黃帚橐吾型退化草地中添加氮素后可以有效提高禾本科牧草高原早熟禾的優勢地位。這是由于高大的禾本科植物本身具有更強的光資源競爭優勢,而添加氮素則可以有效補充土壤的養分[18],因此禾本科牧草的重要值快速提高。此結果與國內外退化草地添加氮素研究結果相符[19-20]。然而,在圍封條件下添加氮素后,禾本科高原早熟禾失去了優勢種地位。這是由于圍封排除了放牧干擾,植物生長主要受物種間競爭影響[21]。而土壤氮素作為黃帚橐吾的重要限制因素[10],在添加氮素后,短期內其重要值明顯提高,占據了禾本科高原早熟禾的生態位,從而使其跌落優勢種地位。而在各處理實施第二年,由于黃帚橐吾密度和生物量的降低,其在圍封和氮添加條件下均跌落優勢種地位。禾本科牧草作為黃帚橐吾的重要競爭物種[21],在黃帚橐吾重要值降低后逐漸占據優勢地位。此外,圍封可以有效降低黃帚橐吾密度和生物量[22],而添加氮素短期內對黃帚橐吾治理效果可能較為有限,在王宏生等[6]研究中也出現了相似結果。
3.2 添加氮素對草地物種多樣性的影響
物種多樣性一直以來都作為研究草地生態結構功能多樣性的重要指標,是物種組成、豐富度、均勻度和相互作用對生態環境影響的綜合體現[23]。在草地生態系統的研究中,國內外學者一般認為短期圍封有利于物種多樣性的增加[24-25],而對于添加氮素對物種多樣性的影響,學者們則存在較大分歧。有些學者認為,氮素會使物種多樣性降低[26-27],而另一些學者則認為,它對物種多樣性沒有顯著影響[28]。本研究表明,草地圍封對黃帚橐吾型退化草地多樣性無顯著影響,并且在圍封和放牧條件下添加氮素后黃帚橐吾型退化草地物種多樣性也無顯著變化。這可能是因為由于恢復年限較短,在Margalef[29]、李永宏等[30]研究中認為草原恢復演替中物種多樣性會呈現波動,但一般出現在恢復演替中后期,所以添加氮素后草地多樣性無顯著變化。另一方面,有研究表示在高寒退化草地中添加氮素會對草地產生負面影響[31-32]。在本研究中,兩年內圍封草地的得分均低于圍封加氮素的草地。然而,由于黃帚橐吾的化感作用會抑制其他植物種子的萌芽和生長[33],植物群落多樣性在兩年內與黃帚橐吾密度均呈現負相關關系。所以黃帚橐吾的密度降低有利于物種多樣性的增加,從而導致氮添加后黃帚橐吾型退化草地的物種多樣性沒有顯著變化。即在毒草型退化草地中草地多樣性的恢復受毒草種類和密度干擾[34]。但隨著草地恢復,黃帚橐吾密度降低對草地的影響也會減少,草地物種多樣性可能會發生變化。
3.3 添加氮素對草地生物量的影響
土壤氮素是草地生物量的重要限制性營養元素[35],添加氮素一直被國內外學者認為有利于草地植物生物量的提高[36-37],但由于草地類型、施氮條件、施氮水平的不同,其結果也存在差異。圍封草地可以通過避免牲畜采食、踐踏等干擾,同樣被認為是恢復退化草地生物量的重要方法[38]。本研究表明,在短期內隨著圍封年限的增加可以有效提高草地總地上生物量,但在圍封條件下添加氮素對草地總地上生物量則無顯著效果,且在圍封草地和放牧草地上添加氮素對草地總地上生物量的影響差異不顯著。這是由于圍封條件下植物生長壓力主要來自物種間的競爭,氮素添加雖然減緩了根系對養分吸收的競爭壓力[39],但使植物從原本的地下養分競爭變為地上的光資源競爭,高大的禾本科植物在此條件下具有較強的競爭優勢[40],從而顯著提高了生物量,但也使得其它科植物生物量明顯下降。因此,圍封條件下添加氮素并未使草地總地上生物量提高。這一結果與劉學敏等[41]在中度退化草地上的生物量變化相符合。同理,相較于放牧草地,圍封草地添加氮素可以有效提高禾本科植物地上生物量,但會使莎草科和豆科生物量顯著降低,而在放牧草地添加氮素,由于禾本科牧草較高位于上層,會受到家畜的優先采食使其生物量明顯降低[42],但上層植被量下降也使得禾本科和豆科等下層植被更容易獲取光照,在地下養分充足的情況下其生物量明顯提高[43],從而導致添加氮素后,圍封草地和放牧草地的各種群間生物量存在顯著差異但總地上物量無顯著差異。但在圍封條件下添加氮素可以顯著提高了地下生物量,這是因為添加氮素可以有效減緩根系的競爭壓力[44]。而在放牧條件下添加氮素則由于動物采食增大了地上部分對養分的需求,根據優先分配原則,植物會優先促進地上部分生長[45],因此地下生物量沒有顯著提高,這一結果與蒯曉妍等[46]同水平氮添加下的研究結果相同。隨著年份的變化禾本科、豆科、雜類草和總地上生物量均發生了顯著變化,但處理間未發生交互作用,這可能是因為受到降雨、氣溫等自然因素影響。
4 結論
綜上所述,氮素添加可以促進對速效養分利用率較高的禾本科牧草的生長發育,使其取得競爭優勢,從而使群落中其他牧草植物(包括有毒植物)的比例下降。在放牧條件下添加氮素能對黃帚橐吾型退化草地生產力起到恢復效果,而在圍封條件下添加氮素則對淺層地下生物量具有極大促進效果,但由于加大了物種間競爭,草地植被得分低于單圍封草地,具體的影響機制有待進一步研究。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:18:18
趣味(語文)(2020年3期)2020-07-27 01:42:46
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
作文與考試·初中版(2017年12期)2017-04-19 20:26:27
爆笑show(2016年7期)2017-02-09 09:36:13
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·少年版(2015年1期)2015-07-07 17:15:12
少兒科學周刊·兒童版(2015年1期)2015-07-07 04:12:52
