含SH2結(jié)構(gòu)域蛋白酪氨酸磷酸酶2表達(dá)變化對(duì)肝纖維化大鼠肝組織中細(xì)胞外信號(hào)調(diào)節(jié)激酶1/2活性的影響
2023-12-09 12:10:58郝禮森潘恩亮季景秀苗笑佳蔣美鈺高瑩瑩
陜西醫(yī)學(xué)雜志 2023年12期
關(guān)鍵詞:模型
郝禮森,潘恩亮,季景秀,苗笑佳,蔣美鈺,王 薇,劉 甜,高瑩瑩
(華北理工大學(xué)附屬醫(yī)院消化內(nèi)科,河北 唐山 063000)
肝纖維化是肝臟受到損傷后的修復(fù)反應(yīng),可進(jìn)展為肝硬化,而肝星狀細(xì)胞是參與肝纖維化的最重要細(xì)胞[1-3]。含SH2結(jié)構(gòu)域的蛋白酪氨酸磷酸酶2(Src homology 2-containing protein tyrosine phosphatase 2,SHP2)是具有促癌作用的蛋白酪氨酸磷酸酶,其過表達(dá)可通過增強(qiáng)細(xì)胞外信號(hào)調(diào)節(jié)激酶1/2(Extracellular signal-regulated kinase1/2,ERK1/2)的活性促進(jìn)腫瘤細(xì)胞增殖,參與多種腫瘤的病理過程[4]。近年來研究發(fā)現(xiàn),SHP2除在多種腫瘤的病理過程中發(fā)揮重要作用外,也參與了某些非腫瘤性疾病的病理過程。研究發(fā)現(xiàn),在四氯化碳誘導(dǎo)的大鼠肝纖維化病程中,肝組織SHP2表達(dá)逐漸升高[5-6];下調(diào)體外活化大鼠肝星狀細(xì)胞SHP2表達(dá)可顯著抑制其增殖[7]。另有研究[8]發(fā)現(xiàn),ERK1/2信號(hào)通路參與了對(duì)活化肝星狀細(xì)胞增殖的調(diào)控。目前,SHP2表達(dá)變化對(duì)纖維化肝組織中ERK1/2的影響尚不清楚。因此,本研究探討SHP2過表達(dá)及低表達(dá)對(duì)肝纖維化大鼠肝組織中ERK1/2活性的影響。
1 材料與方法
1.1 實(shí)驗(yàn)動(dòng)物 健康雄性SD大鼠160只,體重350~400 g,購自北京華阜康生物科技股份有限公司,質(zhì)量合格證編號(hào):11501300046788。所有大鼠均按照清潔級(jí)要求飼養(yǎng),自由飲食和進(jìn)水。實(shí)驗(yàn)動(dòng)物倫理經(jīng)華北理工大學(xué)實(shí)驗(yàn)動(dòng)物倫理委員會(huì)審批。
1.2 主要試劑 表達(dá)綠色熒光蛋白(GFP)的空病毒Ad-GFP、表達(dá)野生型SHP2及GFP的腺病毒Ad-SHP2、表達(dá)GFP及靶向SHP2的RNA干擾序列——短發(fā)夾RNA(shRNA)的腺病毒Ad-shRNA/SHP2由上海漢恒生物科技公司協(xié)助構(gòu)建;兔抗SHP2多克隆抗體(批號(hào):ET1611-35,武漢Abclonal公司);小鼠抗磷酸化ERK1/2(p-ERK1/2)單克隆抗體(批號(hào):16443-1-AP,北京安諾倫生物科技有限公司);小鼠抗ERK1/2單克隆抗體(批號(hào):381050,北京安諾倫生物科技有限公司)。
1.3 實(shí)驗(yàn)方法
1.3.1 大鼠肝纖維化模型構(gòu)建:160只健康雄性SD大鼠隨機(jī)分為對(duì)照組、模型組、Ad-GFP組、Ad-SHP2組和Ad-shRNA/SHP2組,每組32只。將50%四氯化碳橄欖油混合液腹腔注射入模型組、Ad-GFP組、Ad-SHP2組及Ad-shRNA/SHP2組大鼠,每次1 ml/kg,2次/周,共8周[9]。對(duì)照組腹腔注射等量0.9%氯化鈉溶液。每周將滴度為2×109pfu/ml的Ad-GFP、Ad-SHP2及Ad-shRNA/SHP2腺病毒自大鼠尾靜脈分別注射入Ad-GFP組、Ad-SHP2組及Ad-shRNA/SHP2組大鼠[10]。在造模第2、4、6、8周,每組選取8只大鼠取適量肝組織,一部分用于Western blot及實(shí)時(shí)熒光定量PCR檢測(cè),另一部分于4%多聚甲醛固定后用于Masson三色及HE染色和免疫組織化學(xué)染色。
1.3.2 HE染色和Masson三色染色:采用HE染色觀察大鼠肝組織病理學(xué)變化。采用Masson三色染色觀察大鼠肝組織膠原沉積。
1.3.3 實(shí)時(shí)熒光定量PCR檢測(cè)SHP2和ERK1 mRNA表達(dá):提取肝組織總RNA,反轉(zhuǎn)錄合成cDNA。SHP2、ERK1及內(nèi)參3-磷酸甘油醛脫氫酶(GAPDH)引物序列由賽默飛世爾科技公司合成。其中,SHP2正向引物序列為5’-CTTGGTGAGTGAGTAGAT-3’,反向引物序列為5’-CTTACGGCAGAACATTAG-3’;ERK1正向引物序列為5’-GGCTCACCCTTACCTGGAAC-3’,反向引物序列為5’-GTCCAAAAGGACAGGGGGT-3’;GAPDH正向引物序列為5’-GAAAGCCTGCCGGTGACTAA-3’,反向引物序列為5’-AGGAAAAGCATCACCCGGAG-3’。在PCR儀上定量擴(kuò)增,采用2-ΔΔCt法計(jì)算大鼠肝組織SHP2、ERK1 mRNA相對(duì)表達(dá)量[11]。
1.3.4 免疫組化染色檢測(cè)SHP2蛋白表達(dá)水平:采用SP法進(jìn)行SHP2免疫組織化學(xué)染色,陰性對(duì)照用PBS代替一抗,棕褐色為陽性表達(dá)。用Image Pro Plus 6.0軟件分析染色圖像,以積分光密度值 (IOD)表示SHP2陽性表達(dá)水平。
1.3.5 Western blot法檢測(cè)ERK1和p-ERK1蛋白表達(dá):取約100 mg大鼠肝組織剪碎,置于EP管中,加入1 ml改良PIPA裂解緩沖液震蕩研勻,提取蛋白。測(cè)定蛋白含量后取一定量蛋白,添加適量上樣緩沖液,經(jīng)10% SDS-PAGE凝膠電泳后,用水浴式電轉(zhuǎn)儀轉(zhuǎn)至硝酸纖維素膜上,加封閉液4 ℃封閉2 h。分別加入小鼠抗GAPDH單克隆抗體(1∶5000稀釋)、小鼠抗ERK1/2單克隆抗體(1∶500稀釋)和小鼠抗p-ERK1/2單克隆抗體(1∶500稀釋),4 ℃過夜。TPBS漂洗后加入HRP標(biāo)記的山羊抗鼠IgG二抗(1∶5000稀釋),37 ℃孵育2 h。TPBS漂洗后加入ECL顯影液顯色成像,采用Image J 圖像處理軟件分析,ERK1及p-ERK1蛋白表達(dá)量用二者的IOD與GAPDH的IOD比值表示。
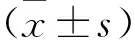
2 結(jié) 果
2.1 各組大鼠肝組織病理形態(tài)學(xué)及膠原沉積變化 見圖1。HE及Masson三色染色結(jié)果顯示,對(duì)照組大鼠肝組織無炎癥細(xì)胞浸潤(rùn),肝細(xì)胞及肝小葉正常,匯管區(qū)及小葉間隔有少量纖維沉積,肝小葉中央靜脈壁可見纖維染色;模型組、Ad-GFP組、Ad-SHP2組以及Ad-shRNA/SHP2組大鼠肝組織隨時(shí)間延長(zhǎng)逐漸出現(xiàn)炎癥細(xì)胞浸潤(rùn)、細(xì)胞變性壞死、正常肝小葉結(jié)構(gòu)消失,甚至出現(xiàn)假小葉,膠原沉積逐漸增加。在同一造模時(shí)間點(diǎn)比較,模型組與Ad-GFP組間上述變化無明顯差異;與模型組及Ad-GFP組比較,Ad-SHP2組上述病理變化及膠原沉積更明顯,而Ad-shRNA/SHP2組減輕。

圖1 各組大鼠造模第6周肝組織HE染色及Masson三色染色結(jié)果(×400)
2.2 不同造模時(shí)間點(diǎn)各組大鼠肝組織SHP2表達(dá)變化 見表1、2(圖2)。在不同造模時(shí)間點(diǎn)(第2、4、6、8周)對(duì)各組大鼠肝組織SHP2 mRNA表達(dá)及蛋白陽性表達(dá)IOD比較,模型組、Ad-GFP組、Ad-SHP2組及Ad-shRNA/SHP2組在各造模時(shí)間點(diǎn)明顯高于對(duì)照組(均P<0.05),Ad-GFP組與模型組在各造模時(shí)間點(diǎn)比較差異無統(tǒng)計(jì)學(xué)意義(均P>0.05),Ad-SHP2組在各造模時(shí)間點(diǎn)較Ad-GFP組及模型組升高(均P<0.05),而Ad-shRNA/SHP2組在各造模時(shí)間點(diǎn)較Ad-GFP組及模型組降低(均P<0.05)。
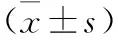
表1 各組大鼠肝組織不同造模時(shí)間點(diǎn)SHP2 mRNA表達(dá)比較
表2 各組大鼠肝組織不同造模時(shí)間點(diǎn)SHP2蛋白陽性表達(dá)IOD比較
2.3 不同造模時(shí)間點(diǎn)各組大鼠肝組織ERK1表達(dá)變化 見表3、4。在不同造模時(shí)間點(diǎn)(第2、4、6、8周)比較,模型組、Ad-GFP組、Ad-SHP2組及Ad-shRNA/SHP2組ERK1 mRNA及蛋白表達(dá)高于對(duì)照組(均P<0.05);但上述四組間比較差異無統(tǒng)計(jì)學(xué)意義(均P>0.05)。
表3 各組大鼠肝組織不同造模時(shí)間點(diǎn)ERK1 mRNA表達(dá)比較
表4 各組大鼠肝組織不同造模時(shí)間點(diǎn)ERK1蛋白表達(dá)水平比較
2.4 不同造模時(shí)間點(diǎn)各組大鼠肝組織p-ERK1蛋白表達(dá)變化 見表5(圖3)。在不同造模時(shí)間點(diǎn)(第2、4、6、8周)對(duì)各組大鼠肝組織p-ERK1蛋白表達(dá)比較,模型組、Ad-GFP組、Ad-SHP2組及Ad-shRNA/SHP2組在各造模時(shí)間點(diǎn)高于對(duì)照組(均P<0.05),Ad-GFP組與模型組在各造模時(shí)間點(diǎn)比較差異無統(tǒng)計(jì)學(xué)意義(均P>0.05),Ad-SHP2組在各造模時(shí)間點(diǎn)較Ad-GFP組及模型組升高(均P<0.05),Ad-shRNA/SHP2組在各造模時(shí)間點(diǎn)較Ad-GFP組及模型組降低(均P<0.05)。
表5 各組大鼠肝組織不同造模時(shí)間點(diǎn)p-ERK1蛋白表達(dá)比較

圖3 各組大鼠肝組織p-ERK1蛋白電泳圖
3 討 論
ERK是絲裂原活化蛋白激酶(MAPK)家族的重要成員,MAPK/ERK信號(hào)通路是一種進(jìn)化保守的信號(hào)級(jí)聯(lián),在細(xì)胞存活、分化、增殖及凋亡的調(diào)控中發(fā)揮重要作用,其活化可促進(jìn)細(xì)胞增殖,抑制細(xì)胞凋亡[12-14]。本研究利用腺病毒載體將靶向SHP2的shRNA及外源性野生型SHP2基因?qū)胨穆然颊T導(dǎo)的肝纖維化大鼠,發(fā)現(xiàn)SHP2低表達(dá)及過表達(dá)對(duì)大鼠纖維化肝組織中ERK1/2的表達(dá)沒有影響,但SHP2低表達(dá)使大鼠纖維化肝組織中p-ERK1/2的表達(dá)下降,而SHP2過表達(dá)使p-ERK1/2的表達(dá)升高。因?yàn)閜-ERK1/2為ERK1/2的活化形式,上述結(jié)果提示SHP2表達(dá)變化影響了大鼠肝纖維化過程中肝組織ERK1/2的活性,即SHP2低表達(dá)使大鼠纖維化肝組織中ERK1/2的活性減弱,SHP2過表達(dá)使ERK1/2的活性增強(qiáng)。SHP2是由原癌基因編碼的具有促癌作用的蛋白酪氨酸磷酸酶,其在腫瘤領(lǐng)域的研究[4,15-18]顯示,在多種惡性腫瘤如乳腺癌、肝癌、口腔鱗狀細(xì)胞癌組織中過表達(dá),通過增強(qiáng)ERK1/2活性促進(jìn)腫瘤細(xì)胞增殖及抑制腫瘤細(xì)胞凋亡參與腫瘤的病理過程,而下調(diào)SHP2表達(dá)可抑制腫瘤細(xì)胞ERK1/2的活性。本研究結(jié)果與SHP2在腫瘤領(lǐng)域的研究結(jié)果一致,提示肝組織SHP2表達(dá)變化可能通過調(diào)控ERK1/2的活性影響肝纖維化。
肝纖維化的病理特征是以膠原為主的細(xì)胞外基質(zhì)在肝內(nèi)的異常沉積。本研究應(yīng)用Masson三色染色檢測(cè)了大鼠肝組織膠原沉積的變化,發(fā)現(xiàn)SHP2低表達(dá)使大鼠肝組織的膠原沉積減輕,而SHP2過表達(dá)則加重了大鼠肝組織的膠原沉積。這提示SHP2表達(dá)變化可通過調(diào)控膠原沉積影響肝纖維化。已有研究[8,19-20]發(fā)現(xiàn),在大鼠肝纖維化病程中,肝組織p-ERK1/2的表達(dá)升高,ERK1/2信號(hào)通路活性增強(qiáng),抑制ERK1/2信號(hào)通路則可抑制肝星狀細(xì)胞增殖,促進(jìn)肝星狀細(xì)胞凋亡,減輕大鼠肝纖維化。結(jié)合本研究中SHP2表達(dá)變化對(duì)大鼠纖維化肝組織ERK1/2的影響,表明ERK1/2參與了SHP2對(duì)肝纖維化的影響,即肝組織中SHP2低表達(dá)可通過抑制ERK1/2活性使肝纖維化減輕,SHP2過表達(dá)可通過增強(qiáng)ERK1/2活性使肝纖維化加重。肝組織由肝細(xì)胞、肝竇內(nèi)皮細(xì)胞、肝星狀細(xì)胞、枯否細(xì)胞等多種細(xì)胞組成,其中肝細(xì)胞是主要細(xì)胞,而在肝纖維化病程中發(fā)揮重要作用的肝星狀細(xì)胞僅占約15%[21]。盡管已有研究[7]發(fā)現(xiàn),體外活化大鼠肝星狀細(xì)胞HSC-T6的SHP2表達(dá)下調(diào)時(shí)細(xì)胞增殖明顯受到抑制,但SHP2表達(dá)變化是通過影響肝臟哪種細(xì)胞的ERK1/2信號(hào)參與肝纖維化尚需進(jìn)一步探討。
綜上所述,大鼠肝纖維化組織中SHP2過表達(dá)可通過促進(jìn)ERK1/2磷酸化增強(qiáng)ERK1/2活性,而SHP2低表達(dá)則可通過抑制ERK1/2磷酸化降低ERK1/2活性。
猜你喜歡
童話王國(guó)·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
網(wǎng)絡(luò)安全與數(shù)據(jù)管理(2022年1期)2022-08-29 03:15:20
導(dǎo)航定位學(xué)報(bào)(2022年4期)2022-08-15 08:27:00
中學(xué)生數(shù)理化·中考版(2022年8期)2022-06-14 06:55:24
新世紀(jì)智能(數(shù)學(xué)備考)(2021年9期)2021-11-24 01:14:36
成都醫(yī)學(xué)院學(xué)報(bào)(2021年2期)2021-07-19 08:35:14
新世紀(jì)智能(數(shù)學(xué)備考)(2020年9期)2021-01-04 00:25:14
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2020年10期)2020-11-26 08:24:50
數(shù)學(xué)物理學(xué)報(bào)(2020年2期)2020-06-02 11:29:24
光學(xué)精密工程(2016年6期)2016-11-07 09:07:19

- 陜西醫(yī)學(xué)雜志的其它文章
- 肉瘤濾過性毒菌致癌同源體B1 V600E基因及生長(zhǎng)分化因子15表達(dá)變化與甲狀腺乳頭狀癌患者淋巴結(jié)轉(zhuǎn)移及預(yù)后關(guān)系研究
- 吉西他濱聯(lián)合熱療對(duì)胰腺癌ASPC-1細(xì)胞增殖、遷移、侵襲和凋亡的影響及機(jī)制研究
- 長(zhǎng)鏈非編碼RNA核富集轉(zhuǎn)錄子1對(duì)胃癌SGC7901細(xì)胞遷移、侵襲、自噬的影響及機(jī)制研究
- 膽綠素還原酶A rs699512和肝細(xì)胞溶質(zhì)載體有機(jī)陰離子轉(zhuǎn)運(yùn)蛋白家族成員1B1 rs4149015位點(diǎn)基因多態(tài)性與新生兒不明原因高膽紅素血癥易感性相關(guān)性研究
- 結(jié)核性胸膜炎合并肺部感染病原菌特點(diǎn)、耐藥性及影響因素分析
- 神經(jīng)精神性系統(tǒng)性紅斑狼瘡患者血清S100A8/A9、神經(jīng)絲輕鏈水平檢測(cè)及臨床意義