晝夜溫差對茶樹生長及茶葉品質的影響
2023-12-18 09:11:08李慧麗
中國茶葉 2023年12期
關鍵詞:生長
李慧麗
成都錦城學院,四川 成都 610097
茶樹原產于亞熱帶森林環境中,具有喜溫耐濕的生長特性,尤其對生長環境的溫度有特定要求,溫度影響著茶樹的生長發育、光合特性及生化成分等,因此,研究溫度對茶樹生長發育的影響規律十分重要。目前,溫度對茶樹生長發育的影響研究已十分廣泛,甚至涉及到極端溫度下對茶樹品種轉錄組學和代謝組學的研究[1]。但探究晝夜溫差對茶樹新梢生長及生化成分合成影響的研究不多。關谷直正等[2]研究發現,當晝夜溫差大于10 ℃時,新梢萌展速度存在減慢趨勢。因而,探究晝夜溫差對茶樹新梢生長及部分生理生化特性影響,對調控茶樹生長發育、提高茶葉品質等具有重要意義。
1 材料與方法
1.1 試驗材料
試驗材料來自于四川省樂山市沐川縣一枝春茶業公司茶樹良種繁育基地(103°90'E,28°96'N),為生長勢一致的省級茶樹品種峨眉問春和川茶2號的1年生茶苗。
1.2 試驗方法
于2020 年11 月將茶苗從品比園移栽至塑料盆缽中,修剪至25 cm 高,以30 株為一個生物學重復,每個處理設3 個生物學重復,共90 株。茶苗適應7 d 后,分別放置于3 個智能人工氣候培養箱(LHP-300H 型,上海三發科學儀器有限公司)中,光照強度、溫濕度條件設置為:30 000 lx白光(LED燈光源),濕度80%/75%(光照/黑暗),光周期12 h/12 h(光照/黑暗),溫度分別設置為25 ℃/20 ℃、25 ℃/15 ℃、25 ℃/10 ℃(光照/黑暗),統一澆水施肥,并進行病蟲害防治,同時通過定期調換茶苗位置,以確保光照條件影響一致。試驗觀察時間共計45 d,并以15 d為一個時段將試驗劃分為前、中、后3個時期。
1.2.1 新梢生長速率及生長量的測定
觀測時間集中在每日上午8:00—10:00,分別于各處理中隨機選取生長勢一致的芽頭共25 個進行掛牌,芽頭萌動后,根據實際生長情況,每隔3~7 d 測量1 次,記錄掛牌茶芽的長度、展葉數,直至長至一芽三葉;測量各處理所有長至一芽三葉的新梢長度、質量,以及部分芽下第二、第三葉葉長及葉寬,以測試不同溫差處理新梢生長速率和生長量的差異。
1.2.2 光合色素含量測定
在不同溫差處理的第15、30、45 d,于上午11:00,分別取不同處理供試茶苗形態學頂端下第二、第三片成熟葉,于-80 ℃分別冷凍保存,在完成3 次取樣后進行集中測定,測定其光合色素含量,測定方法參照《植物生理生化試驗原理和技術》[3]。
1.2.3 氨基酸及兒茶素組分含量測定
待不同溫差處理茶苗的越冬茶芽展葉至一芽三葉時開始采樣。因新梢持續萌發,持續達到采摘標準,將采樣時間按處理時間分為3 個時間段:前期(1~15 d)、中期(16~30 d)和后期(31~45 d),將3 個不同時段采摘新梢樣用液氮速凍后放于冰箱中-80 ℃保存。待試驗結束時,將各時間段內樣品混勻,用于測定咖啡堿(CAF)含量、兒茶素組分及氨基酸組分。
兒茶素組分的測定:采用《茶葉中茶多酚和兒茶素類含量的檢測方法》(GB/T 8313—2018)中的高效液相色譜法(HPLC)測定(按照取樣量0.5 g等比縮小提取液體積,保持濃度不變)。
氨基酸組分的測定:按照《植物中游離氨基酸的測定》(GB/T 30987—2020)中的第三法——氨基酸分析儀法測定(將定容體積縮小至10 mL,提高濃度)。
CAF 測定參照《茶咖啡堿測定》(GB/T 8312—2013)。
1.3 數據分析與處理
使用Excel 2019 進行統計,SPSS Statistics 25進行顯著性分析(P<0.05)。
2 結果與分析
2.1 不同晝夜溫差對茶樹新梢生長的影響
由表1 可知,川茶2 號、峨眉問春兩個品種各處理新梢生長速度均隨晝夜溫差增大而減緩,峨眉問春品種5 ℃晝夜溫差下展至一芽三葉生長速度較10 ℃溫差、15 ℃溫差處理快4.14、7.58 d,川茶2號品種5 ℃晝夜溫差下展至一芽三葉生長速度較10 ℃溫差、15 ℃溫差處理的快1.92、6.64 d。峨眉問春品種整體新梢生長速度略快于川茶2號。
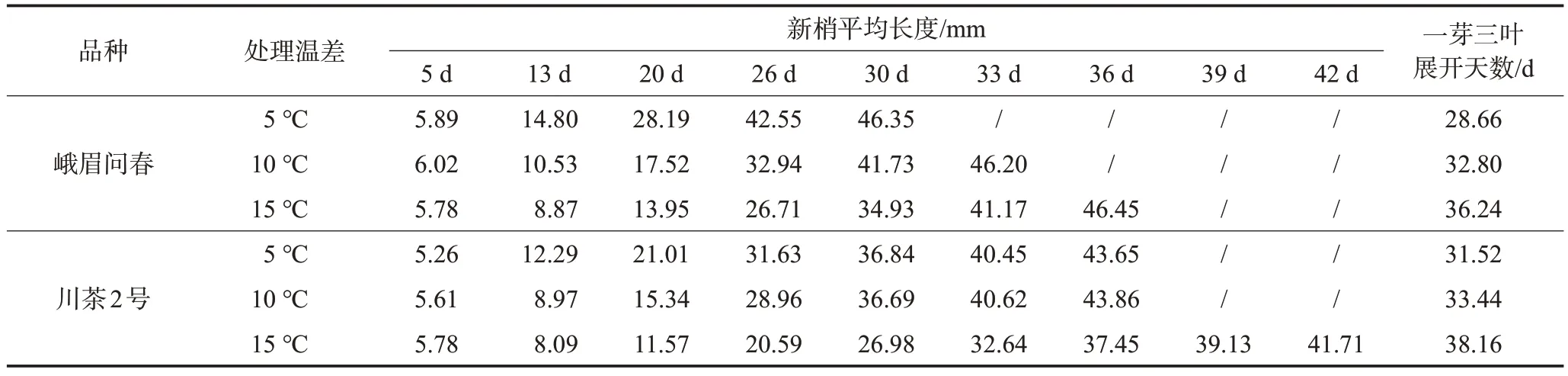
表1 不同溫差處理供試品種在不同處理時間的新梢長度變化
為進一步比較兩個品種不同晝夜溫差處理下茶苗新梢生長情況差異,以15 d 為間隔將整個試驗處理時長分為前、中、后3個時期,計算不同晝夜溫差處理下茶苗新梢日均生長量。從表2 可看出,峨眉問春品種的新梢生長量和生長速度總體超過川茶2 號品種。隨晝夜溫差增大,2 個品種均表現出新梢總生長量下降、生長速度減慢的趨勢。峨眉問春品種3個不同溫差處理下的茶苗日均生長量為1.49~2.07 cm/d,其中5 ℃溫差處理較10 ℃溫差、15 ℃溫差處理日均生長量增加35.29%、38.93%。川茶2號品種3個溫差處理的日均生長量為1.15~1.53 cm/d,其中5 ℃溫差處理較10 ℃溫差、15 ℃溫差處理日均生長量增加10.87%、33.04%。2 個供試品種前、中、后3 個處理時期各處理間的新梢日均生長量有一定差異,但一芽三葉整個生長期的平均生長量則3個處理差異不顯著。除此之外,5 ℃溫差、10 ℃溫差處理下新梢生長速度呈現“慢-快-慢”的S 型曲線,符合新梢生長的基本規律;而15 ℃溫差下由于新梢生長速度慢加之試驗處理時期有限,本試驗中界定的中、后期并不能完全對應新梢生長節律的中、后期,因而新梢日均生長量還未表現出下降趨勢。
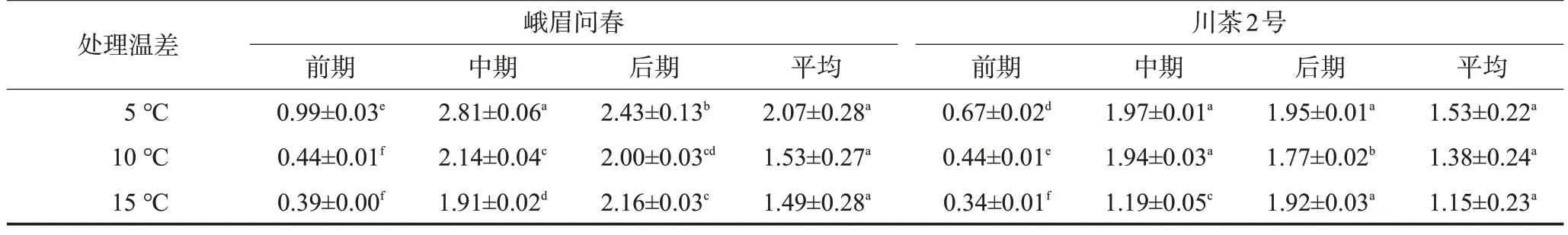
表2 不同溫差處理供試品種新梢日均生長量變化cm/d
綜上,晝夜溫差影響茶樹新梢生長,隨著溫差增大,新梢日均生長量總體下降,生長速度減緩,形成一芽三葉所需的時間增長。
2.2 不同晝夜溫差對茶樹新梢光合色素含量的影響
對兩個品種不同溫差處理下的茶苗光合色素含量進行測定,由于試驗前期處理時間較短,溫差對供試茶苗光合色素形成的影響尚不明顯,因此重點針對中后期茶樹光合色素含量進行分析。由表3可知,兩個供試品種處理中后期的葉片平均葉綠素a含量及葉綠素總量均表現出5 ℃溫差處理顯著高于15 ℃溫差處理,其中峨眉問春品種分別增加21.81%、12.69%,川茶2 號品種分別增加22.34%、24.81%;川茶2 號品種10 ℃溫差處理的平均葉綠素a 含量及葉綠素總量顯著高于15 ℃溫差處理,分別提高22.87%、19.08%,但峨眉問春品種差異并不顯著。
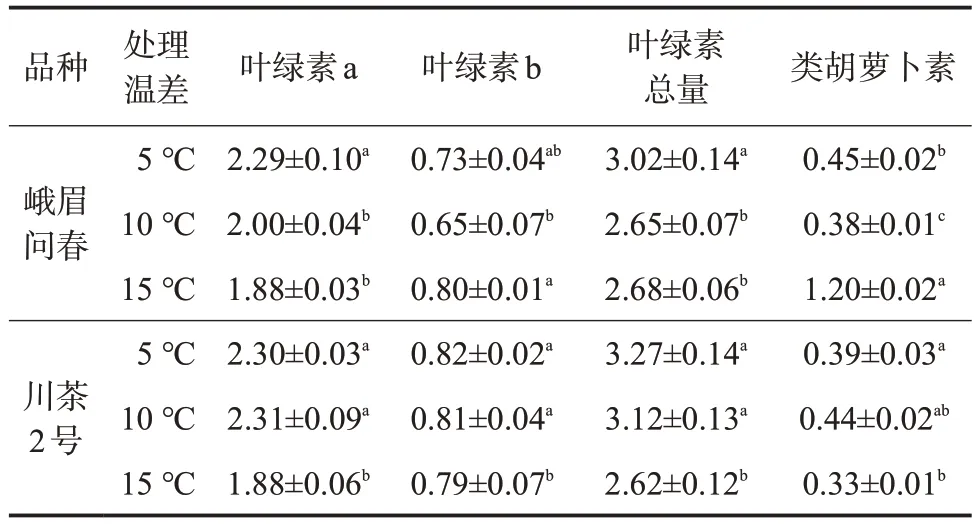
表3 不同溫差處理兩個品種光合色素平均含量mg/g
晝夜溫差對葉片葉綠素b及類胡蘿卜素含量的影響表現出品種差異性,在峨眉問春品種中,此二者含量均表現出溫差處理15 ℃>5 ℃>10 ℃;而在川茶2號品種中,此二者含量則表現出15 ℃溫差處理顯著低于5 ℃溫差處理,與葉綠素a及葉綠素總量變化趨勢一致,中、后期的平均葉綠素b含量溫差5 ℃、10 ℃處理較溫差15 ℃處理分別高3.80%、2.53%,平均類胡蘿卜素含量溫差5 ℃、10 ℃處理較溫差15 ℃處理分別高18.18%、33.33%。
綜上,晝夜溫差影響茶樹葉片光合色素的形成,具體表現為,在處理中、后期,較大的晝夜溫差下葉綠素a及葉綠素總量降低;對葉綠素b及類胡蘿卜素合成的影響,與茶樹品種特性有一定關聯。
2.3 不同晝夜溫差對茶樹新梢兒茶素組分的影響
由于峨眉問春品種新梢采摘時間較為集中,而川茶2號品種新梢樣品處理時間更長,為了數據的準確性,選擇川茶2號品種新梢進行后續兒茶素組分、氨基酸含量測定。
由表4 可見,在本試驗條件下,3 個處理的茶多酚含量和兒茶素總量隨溫差增大呈現先下降后上升趨勢,10 ℃溫差處理下含量最低,但處理間差異不顯著。進一步分析兒茶素各組分含量,除沒食子酸(GA)、表沒食子兒茶素沒食子酸酯(EGCG)含量隨著溫差加大而降低外,其他兒茶素組分均以10 ℃溫差處理下含量最低;10 ℃溫差處理的沒食子兒茶素(GC)、兒茶素(C)含量顯著低于其余兩個溫差處理,表兒茶素(EC)、兒茶素沒食子酸酯(CG)含量顯著低于5 ℃溫差處理,但與15 ℃溫差處理差異不顯著;表兒茶素沒食子酸酯(ECG)、沒食子兒茶素沒食子酸酯(GCG)、表沒食子兒茶素(EGC)含量則3個處理間差異均不顯著。總體來看,10 ℃晝夜溫差下茶苗新梢中以兒茶素類物質為代表的多酚類合成代謝相較于其余兩組處理受到抑制。
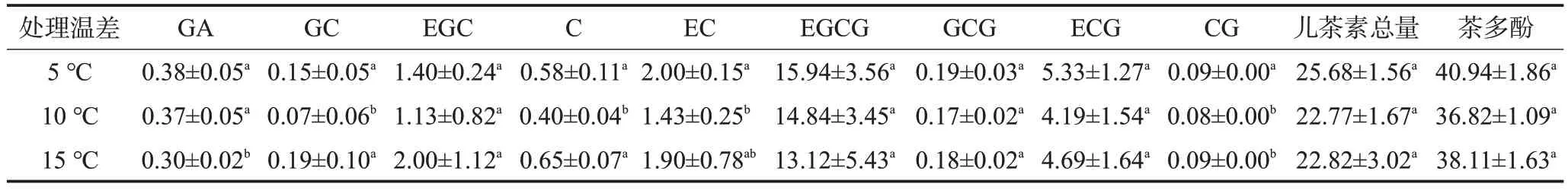
表4 川茶2號新梢兒茶素組分及茶多酚含量mg/g
CAF 合成代謝情況與之相反,10 ℃溫差處理的CAF 含量為(9.51±1.17)mg/g,顯著高于5 ℃溫差處理的(7.70±0.79)mg/g及15 ℃溫差處理的(6.66±0.40)mg/g,且3 個處理間差異均顯著。10 ℃晝夜溫差下CAF合成最為旺盛,而CAF作為含氮化合物,其合成離不開核苷酸、嘌呤代謝等氮代謝過程,由此可推出,在本試驗條件下,在以CAF 為物質合成中心的氮代謝通路中,以及在以茶多酚類物質合成為中心的碳代謝通路中,10 ℃溫差作為適中的晝夜溫差值更有利于前者的物質代謝活動。
2.4 不同晝夜溫差對茶樹新梢游離氨基酸組分的影響
于各個處理3 個時間段取樣測定天冬氨酸(Asp)、蘇氨酸(Thr)、絲氨酸(Ser)、天冬酰胺(Asn)、谷氨酸(Glu)、谷氨酰胺(Gln)、茶氨酸(The)、甘氨酸(Gly)、丙氨酸(Ala)、纈氨酸(Val)、半胱氨酸(Cys)、蛋氨酸(Met)、異亮氨酸(Ile)、亮氨酸(Leu)、酪氨酸(Tyr)、苯丙氨酸(Phe)、 γ- 氨基丁酸(GABA)、 賴氨酸(Lys)、組氨酸(His)、精氨酸(Arg)等20 種氨基酸組分含量及游離氨基酸總量并作顯著性分析,結果見表5。The 含量與游離氨基酸總量隨晝夜溫差增大呈現先上升后下降趨勢,其中5 ℃溫差、15 ℃溫差處理的The 含量較10 ℃溫差處理顯著降低46.76%、57.87%,5 ℃、15 ℃溫差處理的游離氨基酸總量較10 ℃溫差處理顯著降低26.68%、34.44%。這些結果表明新梢The含量與游離氨基酸總量隨晝夜溫差變化與茶多酚變化趨勢相反,即在茶樹中多酚類與氨基酸類物質的合成代謝可能存在相互抑制的現象。
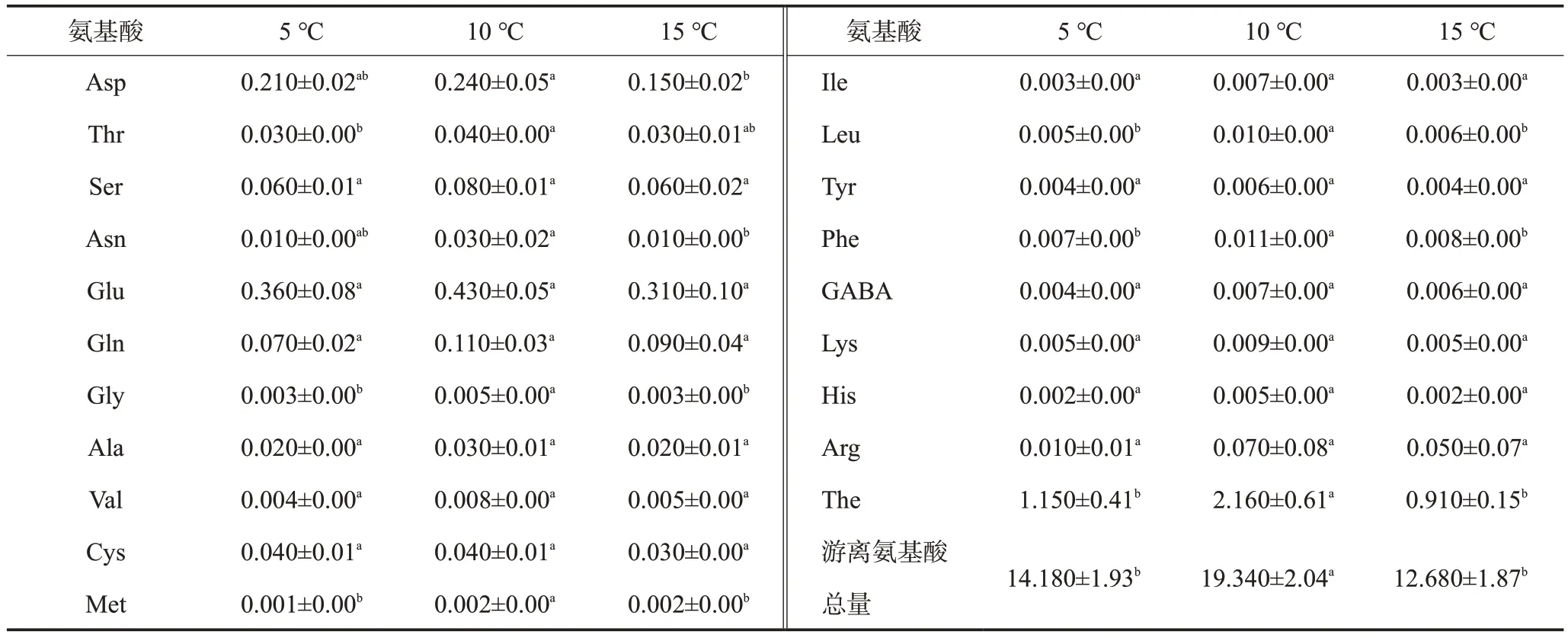
表5 不同溫差處理川茶2號新梢氨基酸組分含量顯著性分析 mg/g
分析3 個溫差處理的氨基酸組分含量,可發現,10 ℃溫差下大部分氨基酸組分(除Cys、Met外)含量均高于其他兩組處理。
3 小結與討論
本研究中,隨晝夜溫差增大,2個供試品種新梢生長速度、日均生長量均逐漸下降,形成一芽三葉所需時間均逐漸增長,即本試驗條件下,晝夜溫差越大,新梢生長速度越緩慢。在處理中后期,較大的晝夜溫差下葉綠素a及葉綠素總量會降低。茶樹新梢咖啡堿與游離氨基酸總量受晝夜溫差影響變化趨勢一致,10 ℃溫差處理含量最高,并與其他兩個處理差異顯著;兒茶素總量與茶多酚含量為10 ℃溫差處理最低,但3 個處理間差異不顯著。
本試驗條件下,作為茶樹重要含氮化合物的CAF 與游離氨基酸含量隨晝夜溫差增大均表現出先上升后下降的趨勢,而作為茶樹碳代謝重要產物的兒茶素及茶多酚含量則隨晝夜溫差增大表現出相反變化。10 ℃溫差處理最大程度地促進了碳代謝通路中游離氨基酸合成支路的代謝強度,使新梢酚氨比最低,這與金珊[4]對設施栽培下茶葉品質影響的研究結果一致,基于1—3 月茶園小氣候背景值觀察結果顯示,設施栽培茶園(晝夜溫差更大)相較于露天茶園酚氨比顯著降低。同時,有研究表明春梢(春季晝夜溫差更大)較秋梢酚氨比顯著降低[5]。本試驗結果中15 ℃晝夜溫差處理下,酚氨比最高,這是由于該溫差處理下游離氨基酸總量過低造成,過大的晝夜溫差會抑制氨基酸合成和轉運。
本試驗研究為理想模型,在控濕、控光的情況下,研究了單一因素晝夜溫差對新梢碳氮代謝的影響,且溫差處理時間有限,而在自然環境中濕度、光照、種植密度或間種等對茶樹新梢生長及代謝的影響是不可忽略的,如光照也影響茶樹光合作用以及碳、氮代謝的強度和方向,因此后期可將溫差與光照等因子結合進行試驗,從而分析不同光照與溫差對新梢碳氮代謝的影響,以及對比二者對功能性成分合成、各類代謝水平影響的強弱及協同(拮抗)作用。
其次,本試驗溫差處理采用的是白天溫度保持一定,通過降低夜溫來增大晝夜溫差的方式,但并未控制均溫一定,后期可通過增加溫差設置,保持均溫(最適溫度)一定的設計,進一步探討新梢或根部生長發育的晝夜溫差組合。
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
