潛在抗肝纖維化藥物與靶點相關(guān)信號通路研究進展
2023-12-18 04:10:08何雪茹付裕豪荀雪姣董占軍
臨床肝膽病雜志 2023年12期
關(guān)鍵詞:信號
周 鑫, 王 智, 何雪茹, 付裕豪, 荀雪姣, 李 穎, 董占軍
1 河北醫(yī)科大學(xué)研究生學(xué)院, 石家莊 050017; 2 河北省人民醫(yī)院藥學(xué)部, 石家莊 050057
肝纖維化是由非酒精性脂肪性肝炎、病毒性肝炎、過度飲酒和金屬中毒等引起的慢性肝損傷的傷口愈合的過程。不加干預(yù)的肝纖維化將會進展為肝硬化、肝癌,最終導(dǎo)致死亡[1]。全球約75%的肝癌發(fā)生在亞洲,僅我國就承擔(dān)了全球50%的肝癌負(fù)擔(dān)[2]。大量研究[3]表明,肝纖維化是可控的,甚至是可逆的。因此,及早地干預(yù)肝纖維化將極大地減輕我國的社會負(fù)擔(dān)和醫(yī)療壓力,延緩、逆轉(zhuǎn)肝纖維化已成為治療慢性肝炎的關(guān)鍵選擇。細(xì)胞外基質(zhì)(ECM)在肝臟內(nèi)的過度沉積是肝纖維化的一個標(biāo)志特征,作為其主要貢獻(xiàn)者的肝星狀細(xì)胞(HSC)在肝纖維化進行性發(fā)展中有非常重要的作用。肝纖維化是一個動態(tài)的過程,HSC 的活化與其他細(xì)胞之間的作用受多種信號通路之間復(fù)雜的相互作用影響[4]。同時,細(xì)胞的自噬與凋亡在HSC 的活化中也有多方面的影響。本文介紹了多條信號通路在肝纖維化進程中的影響并綜述了非編碼RNA 在肝纖維化進程中的不同作用,以期發(fā)現(xiàn)抗肝纖維化相關(guān)潛在候選目標(biāo)與新的治療靶點,為延緩、逆轉(zhuǎn)肝纖維化提供新的思路。
1 肝纖維化相關(guān)信號通路
1.1 TGF-β介導(dǎo)的信號通路 TGF-β是一種多功能細(xì)胞因子,調(diào)節(jié)巨噬細(xì)胞、未成熟造血細(xì)胞和樹突狀細(xì)胞等多種細(xì)胞的增殖、分化、凋亡、黏附和遷移。TGF-β通過經(jīng)典(基于SMAD)和非經(jīng)典(非基于SMAD)信號通路表達(dá)信號響應(yīng)。TGF-β激活下游介質(zhì),進一步激活幾種細(xì)胞內(nèi)信號通路以調(diào)節(jié)各種細(xì)胞功能。
1.1.1 TGF-β1/SMAD 信號通路與相關(guān)活性成分及藥物 TGF-β1 通過激活下游底物(包括SMAD2 和SMAD3)發(fā)揮其生物學(xué)作用,同時受到SMAD7表達(dá)的負(fù)反饋調(diào)節(jié)[5]。在肝纖維狀態(tài)下,SMAD2和SMAD3表達(dá)增加起正向調(diào)節(jié)作用,而SMAD7表達(dá)下降進行負(fù)調(diào)節(jié)。TGF-β/SMAD的信號傳導(dǎo)起始于一個異源傳導(dǎo)復(fù)合物,當(dāng)TGF-β1與TGF-β受體Ⅱ(TGFβRⅡ)相互作用時,異源傳導(dǎo)復(fù)合物被激活,誘導(dǎo)TGF-βRⅡ?qū)GFβRⅠ進行磷酸化,接著磷酸化TGFβRⅠ反過來磷酸化細(xì)胞質(zhì)介質(zhì)中的SMAD2/SMAD3,并且與SMAD4 形成異源三聚體復(fù)合物,逐漸向細(xì)胞核內(nèi)轉(zhuǎn)移,識別結(jié)合序列發(fā)揮轉(zhuǎn)錄因子作用[6]。
柚木具有抗炎保肝解熱鎮(zhèn)痛作用。柚木葉取物以劑量依賴性降低Ⅰ型α1 膠原鏈(COL1A1)、α-平滑肌肌動蛋白(α-SMA)、血小板衍生生長因子和組織金屬蛋白酶抑制劑1(TIMP1)的基因表達(dá),升高基質(zhì)金屬蛋白酶3(MMP3)的表達(dá)。同時柚木提取物在mRNA水平上呈劑量和時間依賴性,在蛋白水平上呈劑量依賴性,顯著下調(diào)TGF-β1 和SMAD2 表達(dá),上調(diào)SMAD7表達(dá),通過調(diào)節(jié)TGF-β1/SMAD2 通路和上調(diào)MMP3/TIMP1 比值逆轉(zhuǎn)肝纖維化[7]。芝麻酚是一種從芝麻中分離出的脂溶性木酚素類化合物,被證明對硫代乙酰胺(TAA)誘導(dǎo)的肝纖維化大鼠具有保護作用。芝麻酚以劑量依賴性方式顯著降低TGF-β1、SMAD3、磷酸化的SMAD3(p-SMAD3)和α-SMA mRNA的表達(dá)和蛋白質(zhì)濃度,上調(diào)SMAD7 mRNA 的表達(dá),同時以濃度依賴性方式顯著降低溶血磷脂酸受體1(LPAR1),LPAR3和結(jié)締組織生長因子mRNA 的表達(dá),通過抑制TGF-β1/SMAD3 信號通路和下調(diào)LPAR1/3 的表達(dá)發(fā)揮抗肝纖維化作用[8]。阿司匹林在體內(nèi)外以劑量依賴性方式下調(diào)α-SMA、Ⅰ型膠原蛋白、TGF-β1、p-SMAD2/3 的蛋白質(zhì)表達(dá)水平和α-SMA、COL1A1、TGF-β1 mRNA 水平,通過抑制TGF-β1/SMAD 信號減輕肝纖維化[9]。檸檬素通過上調(diào)SMAD7,減少SMAD2/3 的c 端磷酸化(p-SMAD2/3c)和核轉(zhuǎn)位,抑制TGF-β/SMAD 信號通路來減輕肝細(xì)胞EMT 和HSC 活化,從而抑制四氯化碳(CCl4)誘導(dǎo)的小鼠肝纖維化[10]。
1.1.2 TGF-β1/其他信號通路與相關(guān)活性成分及藥物 除規(guī)范的TGF-β1/SMAD 信號通路外,還存在由其他信號通路介導(dǎo)的TGF-β信號傳導(dǎo)。例如,絲裂原活化蛋白激酶(MAPK)通路,包括細(xì)胞外信號調(diào)節(jié)激酶、c-Jun氨基末端激酶(JNK)、p38 MAPK,以及NF-κB抑制劑(IκB)激酶(IKK)、磷脂酰肌醇-3激酶(PI3K)、蛋白激酶B(Akt)和Rho 家 族GTPases,這 些 受 體 激 活 的 非SMAD 轉(zhuǎn)導(dǎo)器以獨立或與SMAD 聯(lián)合的方式介導(dǎo)信號反應(yīng),并匯聚到SMAD上以控制SMAD活動[11]。
咖啡因阻斷了磷酸化的ERK、JNK、p38 和pSMAD3L 蛋白水平的顯著增加,通過阻斷MAPK 和TGF-β/SMAD3 信號通路,減輕實驗性非酒精性脂肪性肝炎和TAA 誘導(dǎo)的肝纖維化進展[12]。米糠油(RBO)富含多種生物活性植物化學(xué)物質(zhì),具有降低膽固醇、抗炎抗氧化活性等作用。RBO 顯著下調(diào)NF-κβ 信號通路,進而抑制一些炎癥介質(zhì)的表達(dá),包括環(huán)氧化酶2、IL-1β和TNF-α。RBO通過抑制TGF-β1、α-SMA、Ⅰ型膠原、羥脯氨酸、結(jié)締組織生長因子和黏著斑激酶(FAK)的生物學(xué)作用,通過抑制HSC 活化和調(diào)節(jié)TGF-β1 和FAK信號轉(zhuǎn)導(dǎo)之間的相互作用來減少肝纖維化[13]。五味子酯甲具有抗氧化抗炎抗細(xì)胞凋亡和心肌保護等多種藥理活性,降低了磷酸化的TGF-β-活化激酶1(p-TAK1)/TAK1、p-JNK/JNK、p-p38/p38、p-IκB/IκB 和NF-κB p65 蛋白表達(dá)水平,不同程度上抑制了α-SMA 和COL1A1 mRNA的表達(dá),通過TGF-β1介導(dǎo)的TAK1/MAPK和NF-κB通路的激活,在體內(nèi)外改善肝纖維化[14]。
1.2 Wnt/β-catenin 信號通路與相關(guān)活性成分及藥物Wnt 信號通路是一種比較保守的信號轉(zhuǎn)導(dǎo)通路,在調(diào)節(jié)細(xì)胞功能和生長發(fā)育方面發(fā)揮重要作用。Wnt蛋白首先與靶細(xì)胞膜上的卷曲蛋白和輔助受體低密度脂蛋白受體相關(guān)蛋白5/6結(jié)合,從而通過蓬亂蛋白的磷酸化將胞外信號傳遞到細(xì)胞質(zhì)內(nèi)。然后細(xì)胞質(zhì)內(nèi)的蓬松蛋白通過抑制糖原合成酶激酶-3β(GSK-3β)的激活,阻止β-catenin 的磷酸化或降解,導(dǎo)致細(xì)胞質(zhì)內(nèi)游離的β-catenin增加。當(dāng)細(xì)胞質(zhì)中游離的β-catenin達(dá)到一定水平時,進入細(xì)胞核與核內(nèi)淋巴細(xì)胞增強因子/轉(zhuǎn)錄T淋巴細(xì)胞因子(LEF/TCF)結(jié)合,形成β-catenin-LEF/TCF復(fù)合體,導(dǎo)致經(jīng)典Wnt信號通路下游靶基因轉(zhuǎn)錄[15]。
漆黃素顯著抑制與β-catenin 下降和GSK-3β 水平升高相關(guān)的wnt3a基因表達(dá)。此外,漆黃素還能減緩肝組織纖維化的進程,減少α-SMA和特異性周期蛋白-D1(cyclin D1)的肝臟表達(dá),通過抑制Wnt/β-catenin 信號通路減輕TAA 誘導(dǎo)的大鼠肝纖維化[16]。和厚樸酚(HNK)是從廣玉蘭中提取的一種生物活性化合物,具有潛在的抗肝纖維化作用。HNK 激活了CC-3 和GSK3β,下調(diào)了HSC 中的α-SMA、Wnt3a 和β-catenin 的表達(dá),通過激活GSK3β 和抑制Wnt3a/β-catenin 信號通路誘導(dǎo)細(xì)胞凋亡和抑制肝纖維化[17]。
1.3 PI3K/Akt/哺乳動物雷帕霉素靶蛋白(mTOR)信號通路與相關(guān)活性成分及藥物 PI3K/Akt/mTOR 信號通路是癌癥的關(guān)鍵通路之一[18]。大量的研究表明PI3K/Akt/mTOR 信號通路在肝纖維化中發(fā)揮著重要作用。不同的生長因子對應(yīng)不同的受體酪氨酸激酶(RTK)。首先生長因子介導(dǎo)的RTK 刺激PI3K 的活化。活化的PI3K 使磷脂酰肌醇4,5-二磷酸(PIP2)磷酸化并將其轉(zhuǎn)化為磷脂酰肌醇3,4,5-三磷酸(PIP3)。第二個信使PIP3 是Akt 的對接位點,而Akt 是PI3K 通路的核心介質(zhì)。隨后磷脂酰肌醇依賴性激酶14 與Akt之間的相互作用,導(dǎo)致Akt在thr308位點上進行磷酸化[19]。磷酸化的Akt通過結(jié)節(jié)性硬化癥1/2 復(fù)合體激活下游效應(yīng)蛋白mTOR,并上調(diào)各種轉(zhuǎn)錄因子,從而發(fā)揮其生物學(xué)活性作用。
多沙唑嗪是一種選擇性α1 腎上腺素能受體阻斷劑,被發(fā)現(xiàn)有抗肝纖維化作用。多沙唑嗪在體內(nèi)外以濃度依賴性方式抑制HSC 的活化,上調(diào)PI3K/Akt/mTOR 信號通路中的p-mTOR 的表達(dá)從而抑制自噬來減弱肝纖維化[20]。富馬酸替諾福韋二吡呋酯主要用于慢性乙型肝炎患者的抗病毒治療,通過持續(xù)顯著降低PI3K/Akt/mTOR 信號通路的磷酸化在TAA 誘導(dǎo)的肝纖維化模型中誘導(dǎo)HSC凋亡[21]。委陵菜酸是從委陵菜中提取的一種活性成分,委陵菜酸顯著降低了α-SMA、TGF-β、半胱天冬酶-3、Bax、IL-6 和TNF-α 的表達(dá),升高了Bcl-2 和IL-10 的表達(dá),通過抑制甘油磷脂代謝途徑以及PI3K/Akt/mTOR 和NF-κB 信號通路,緩解體內(nèi)肝纖維化進展[22]。
1.4 NF-κB信號通路與相關(guān)活性成分及藥物 NF-κB信號通路是一種高度保守的進化途徑,在調(diào)節(jié)免疫和炎癥反應(yīng)方面具有相當(dāng)重要的作用。PI3K/Akt 可調(diào)控NF-κB 通路/轉(zhuǎn)錄因子家族,而NF-κB 通路/轉(zhuǎn)錄因子家族可調(diào)控炎癥、細(xì)胞應(yīng)激、先天和適應(yīng)性免疫反應(yīng),進而調(diào)控肝細(xì)胞、肝巨噬細(xì)胞和HSC 的生存、增殖、遷移和侵襲[23]。
NF-κB 包括RelA(p65)、RelB、c-Rel、p105/p50 和p100/p52五個家族成員。Rel蛋白作為成熟蛋白合成,p50 和p52 分別由前體p105 和p100 的蛋白體降解產(chǎn)生。這些蛋白具有保守的Rel 同源結(jié)構(gòu)域,可與目標(biāo)DNA 序列結(jié)合,形成homo/異二聚體,在沒有任何刺激的情況下,通過IκB 定位于細(xì)胞質(zhì)。NF-κB 的活化通過不同的典型和非典型途徑都可以進行,在規(guī)范途徑中,抑制劑IκB被IKK復(fù)合物磷酸化,并受到泛素介導(dǎo)的蛋白酶體降解。游離的NF-κB 二聚體(主要是p50/p65 和p50/c-Rel)進入細(xì)胞核內(nèi)、并激活靶基因轉(zhuǎn)錄。在非典型途徑中,NF-κB 誘導(dǎo)激酶被激活,隨后p100被翻譯后加工為p52 亞基,與RelB 二聚,以及核定位,從而誘導(dǎo)基因轉(zhuǎn)錄[24]。
二氫楊梅素是騰茶提取物中的一種天然活性物質(zhì),二氫楊梅素抑制了促凋亡蛋白Bax、半胱天冬酶-3/9(CC-3/9)、磷酸化蛋白及其上游炎癥調(diào)節(jié)因子p-IKKα/β和p-IκBα、TGF-β1 和α-SMA 的表達(dá),促進抗凋亡蛋白Bcl-2 和Bcl-XL 的表達(dá)。通過抑制NF-κB 介導(dǎo)的炎癥和TGF-β1 調(diào)節(jié)PI3K/Akt 信號通路來逆轉(zhuǎn)TAA 誘導(dǎo)的肝纖維化[25]。甲氧基丁香酚(4-烯丙基-2,6-二甲氧基苯酚)是存在于人類飲食中的成分,被廣泛用作食品工業(yè)中的添加劑,顯著降低了Col-1、α-SMA、IL-6、TNF-α、一氧化氮合成酶和IL-8 的表達(dá),通過NF-κB/過氧化物酶體增殖物激活受體-?和NF-κB機制減弱肝纖維化和炎癥[26]。獐牙菜被用來治療包括肝臟疾病在內(nèi)的各種疾病,獐牙菜提取物顯著抑制CCl4誘導(dǎo)的大鼠肝纖維化模型中TGF-β、α-SMA、SMAD2/3、Col1A1、NF-κB、TNF-α和IL-1β mRNA和蛋白的表達(dá),恢復(fù)了肝組織中改變的鐵調(diào)素水平,通過抑制TGF-β/SMAD 和NF-κB信號通路減輕大鼠肝纖維化并恢復(fù)肝鐵調(diào)素水平[27]。西他列汀以其抗糖尿病活性而聞名,提高了Nrf2 及其靶基因的mRNA 表達(dá),顯著降低了NF-κB 的水平和蛋白表達(dá)以及TNF-α 的蛋白表達(dá),通過激活Nrf2和抑制NF-κB通路發(fā)揮抗肝纖維化作用[28]。
1.5 Hippo 信號通路與相關(guān)活性成分及藥物 Hippo途徑是一種進化保守的信號通路,在傷口愈合和免疫調(diào)節(jié)中發(fā)揮關(guān)鍵作用。河馬通路由一個信號網(wǎng)絡(luò)組成,這些信號最終指示轉(zhuǎn)錄調(diào)節(jié)因子YES 關(guān)聯(lián)蛋白(YAP)和PDZ結(jié)合基序轉(zhuǎn)錄共激活因子(TAZ)的功能。核YAP/TAZ 直接進行基因表達(dá),調(diào)控細(xì)胞增殖和存活等一系列行為[29]。YAP/TAZ的消耗會阻斷TGF-β誘導(dǎo)的肌成纖維細(xì)胞轉(zhuǎn)化和ECM 的產(chǎn)生,而活化的YAP/TAZ 的異位表達(dá)會促進這些情況。YAP 與TAZ 結(jié)合直接調(diào)節(jié)TGF-β 激活的SMAD 轉(zhuǎn)錄因子,為促纖維化信號傳導(dǎo)串?dāng)_提供了一種機制。
白藜蘆醇是一種存在于葡萄花生等作物中的生物活性成分,已被證明通過多種信號通路發(fā)揮抗肝纖維化作用。白藜蘆醇顯著降低了YAP和TAZ的mRNA和蛋白水平,增強Bax 蛋白表達(dá)并降低Bcl-2 蛋白表達(dá),部分通過Hippo通路抑制HSC活化[30]。N-乙酰-L-色氨酸(L-NAT)是N-乙酰色氨酸(NAT)的三種異構(gòu)體之一,以往研究發(fā)現(xiàn)其具有神經(jīng)保護抗炎保肝等作用。L-NAT 顯著降低α-SMA、Ⅰ膠原、TGF-β1、p-SMAD2/3、MMP9 和Bax 蛋白水平的表達(dá)。增加Bcl-2 和YAP1 的蛋白表達(dá),通過調(diào)控TGF-β1/SMAD 和Hippo/YAP1 信號抑制CCl4誘導(dǎo)的肝纖維化[31]。
1.6 MAPK 信號通路與相關(guān)活性成分及藥物MAPK 途徑是最保守的信號轉(zhuǎn)導(dǎo)途徑之一,在細(xì)胞增殖、分化、死亡和胚胎發(fā)生中具有關(guān)鍵功能。MAPK信號傳導(dǎo)由三種基于激酶的反應(yīng)組成,通過這些反應(yīng),由絲裂原激活的蛋白激酶激酶(MAPKK 或MEK)催化的上游激酶反應(yīng)激活,而上游激酶反應(yīng)又被另一種稱為MAPKKK(MAPKK 激酶)或Raf(以快速加速纖維肉瘤命名)的上游激酶磷酸化[32]。常規(guī)MAPK包括JNK、p38 MAPK 和ERK。已有研究表明MAPK 通路與肝纖維化之間有著密切關(guān)系。
香芹酚是一種單萜烯酚類的非選擇性TRPM7抑制劑,具有抗腫瘤、抗炎和抗肝纖維化作用,香芹酚顯著降低了CCl4誘導(dǎo)肝纖維化小鼠模型中α-SMA、Col1α1、p-ERK1/2、p-JNK1/2和p-p38的表達(dá),通過阻斷MAPK通路發(fā)揮抗肝纖維化作用[33]。HNK是從廣玉蘭中提取的一種生物活性化合物,具有潛在的抗肝纖維化作用。HNK降低了α-SMA、COL1A1、COL3A1和SMAD2/3的表達(dá),增加了微管相關(guān)蛋白1輕鏈3-Ⅱ(LC3-Ⅱ)和p62的表達(dá),通過抑制TGF-β1/SMAD 信號通路和激活p38 MAPK通路分別介導(dǎo)HSC的活化與自噬,起到抗肝纖維化作用[34]。所有肝纖維化信號通路與潛在抗肝纖維化藥物及活性成分詳見表1。
2 非編碼RNA(ncRNA)介導(dǎo)的肝纖維化信號通路
人類基因組中約80%的基因被主動轉(zhuǎn)錄為各種ncRNA,在多種疾病發(fā)病機制的重要途徑中起關(guān)鍵作用。ncRNA 包括長鏈RNA(lncRNA)、微小RNA(miRNA)、環(huán)狀RNA(circRNA)、小核仁氨酸(snoRNA)和Piwi相互作用的RNA(piRNA)。大量研究[35]表明,ncRNA 可以通過調(diào)節(jié)多個下游基因靶點和相關(guān)途徑的表達(dá)能力,調(diào)控肝纖維化進程(表2)。
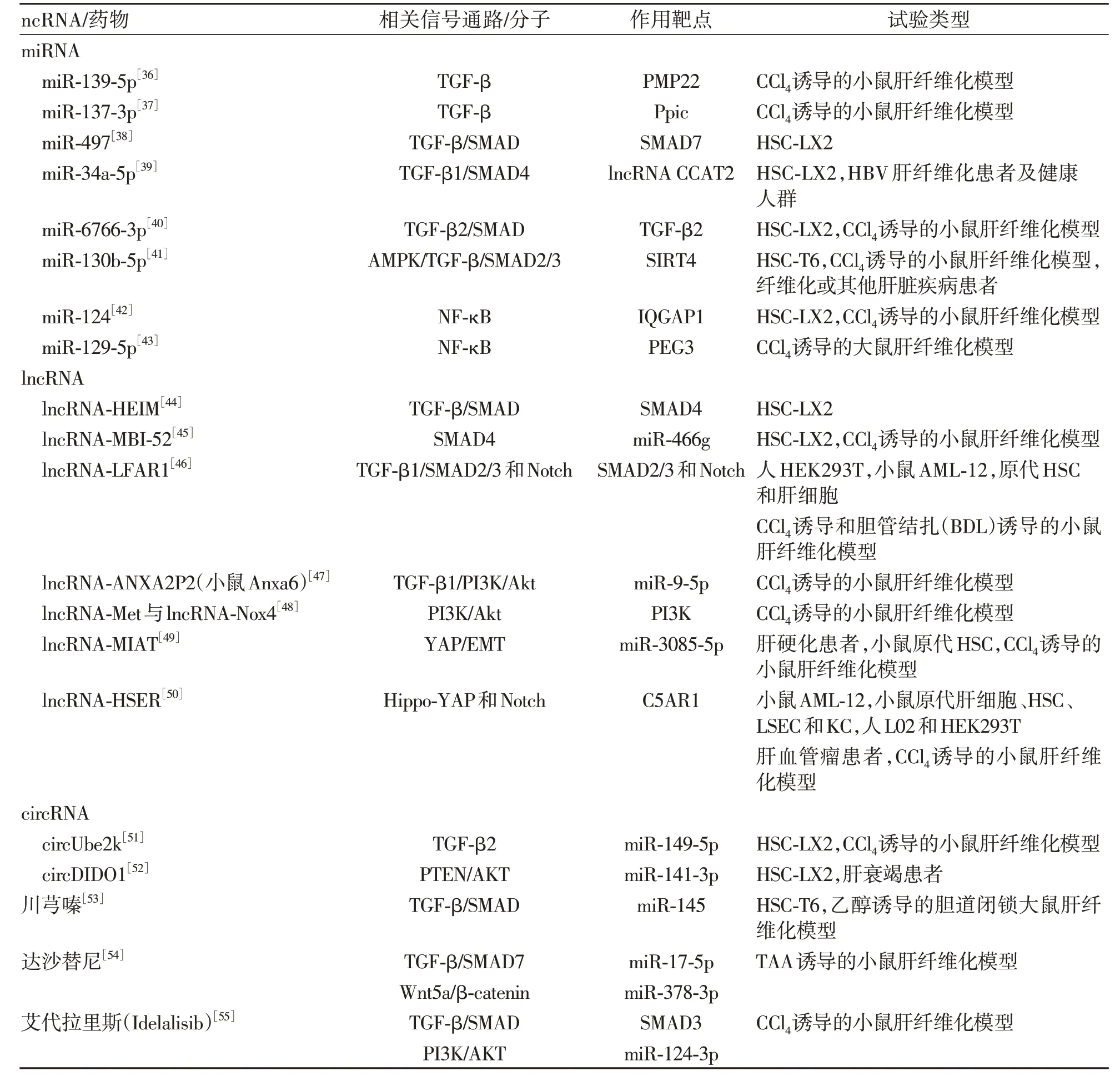
表2 ncRNA及藥物與肝纖維化相關(guān)通路與試驗類型總結(jié)Table 2 Summary of non-coding RNAs and drug and liver fibrosis-related pathways and trial types
2.1 miRNA 介導(dǎo)的肝纖維化信號通路及其靶點miRNA 是長度約22 個核苷酸的內(nèi)源性ncRNA,在轉(zhuǎn)錄后水平調(diào)節(jié)基因表達(dá)。外周髓鞘蛋白22(PMP22)首先從小鼠NIH3T3 成纖維細(xì)胞中分離出來,主要表達(dá)在周圍神經(jīng)的髓鞘化雪旺細(xì)胞中,研究[36]表明PMP22 在CCl4誘導(dǎo)的小鼠肝纖維化模型和TGF-β 激活的HSC 中顯著上調(diào),miR-139-5p 通過靶向抑制PMP22 阻礙TGF-β 誘導(dǎo)的HSC 的激活。肽基脯氨酰異構(gòu)酶C(Ppic)在骨髓中的造血祖細(xì)胞和胸腺中的T淋巴細(xì)胞前體細(xì)胞或胸腺細(xì)胞中表達(dá),以往研究表明Ppic 在環(huán)孢素(CsA)誘導(dǎo)的腎損傷起重要作用。在CCl4誘導(dǎo)的小鼠肝纖維化模型中,Ppic 敲低降低了TGF-β 誘導(dǎo)的小鼠HSC 活化,miR-137-3p 直接靶向并負(fù)調(diào)節(jié)Ppic 的表達(dá)調(diào)控TGF-β 引起的小鼠HSC 活化[37]。miR-497 在多種疾病過程中起到一定作用,是潛在的生物標(biāo)志物與治療靶點。miR-497 可以通過靶向SMAD7 增強TGF-β/SMAD 信號通路以促進肝纖維化[38]。miR-34a-5p 靶向并負(fù)調(diào)節(jié)lncRNA CCAT2 的表達(dá),通過TGF-β1/SMAD4 信號通路發(fā)揮抗肝纖維化作用[39]。三維人胚胎干細(xì)胞外泌體(3D-hESC-外泌體)中含有非編碼小RNA與蛋白質(zhì)等其他生物分子,已被證明可以增強組織再生并調(diào)節(jié)內(nèi)源性修復(fù)。3D-hESC-外泌體中的miR-6766-3p 通過抑制TGFβRII 表達(dá)來滅活SMAD 信號傳導(dǎo),從而減弱HSC 活化并抑制肝纖維化[40]。miR-130b-5p 的表達(dá)在多種肝損傷模型中上調(diào),miR-130b-5p通過AMPK/TGF-β/SMAD2/3信號通路調(diào)節(jié)SIRT4,從而促進肝纖維化[41]。含有IQ 基序的GTP 酶激活蛋白1(IQGAP1)是一種信號支架蛋白,通過促進細(xì)胞中的信號轉(zhuǎn)導(dǎo)來調(diào)節(jié)一系列細(xì)胞活動。研究[42]發(fā)現(xiàn)miR-124的表達(dá)在肝纖維化組織和TNF-α誘導(dǎo)的LX-2 細(xì)胞中下調(diào),同時伴隨著IQGAP1 的上調(diào)表達(dá),miR-124 的過表達(dá)可通過靶向IQGAP1 抑制NF-κB途徑來降低HSC 炎性細(xì)胞因子的分泌。父系表達(dá)基因3(PEG3)是一種編碼DNA結(jié)合蛋白的印記基因,是一種已知的轉(zhuǎn)錄抑制因子。miR-129-5p 可能調(diào)節(jié)纖維腺炎相關(guān)基因的表達(dá),miR-129-5p 在CCl4誘導(dǎo)的大鼠模型肝纖維化組織中表達(dá)較低,PEG3 表達(dá)較高。研 究[43]表明miR-129-5p 可以通過降 低PEG3 抑制NF-κB信號通路發(fā)揮抗肝纖維化作用。
2.2 lncRNA 介導(dǎo)的肝纖維化信號通路及其靶點lncRNA 是一類長度大于200 個核苷酸且不具有蛋白質(zhì)編碼能力的ncRNA。lncRNA-ENST00000519726(HEIM)是一種新鑒定的lncRNA。lncRNA-HEIM 的表達(dá)在HBV 感染后上調(diào),導(dǎo)致TGF-β 和SMAD4 的表達(dá)增加,并進一步促進HSC 的纖維化[44]。lnc-MBI-52在體內(nèi)和體外肝纖維化過程中均增加,lnc-MBI-52 可以通過抑制miR-466g表達(dá)來抑制SMAD4 表達(dá)發(fā)揮抗肝纖維化作用[45]。lnc-LFAR1 在HSC 中特異性上調(diào),通過激活TGFβR1/SMAD2/3 和Notch 信號通路促進肝纖維化[46]。小鼠Anxa6 敲低部分在TGF-β1 刺激的HSC 中 抑 制 了TGF-β1 誘 導(dǎo) 的HSC 活 化 并 阻 斷 了PI3K/Akt 信號激活,小鼠Anxa6/miR-9-5p/Anxa2 軸和PI3K/Akt 通路可能參與了lncRNA ANXA2P2(mouse Anxa6)對 肝 纖 維 化 的 作 用[47]。 lncRNA-Met 與lncRNA-Nox4是兩種新型lncRNA,共表達(dá)可以通過氧化還原過程和PI3K/Akt信號通路激活HSC,增加ECM的產(chǎn)生,并調(diào)節(jié)肝纖維化的過程[48]。lncRNA-MIAT的表達(dá)與miR-3085-5p 水平在肝硬化患者和活化的HSC 中呈負(fù)相關(guān),lncRNA-MIAT 至少部分通過miR-3085-5p/YAP/EMT 信 號 通 路 增 強HSC 的 活 化[49]。lncRNA-HSER 在肝細(xì)胞中特異性表達(dá),通過C5AR1-Hippo-YAP 和Notch 信號通路分別抑制肝細(xì)胞凋亡和肝細(xì)胞上皮間質(zhì)轉(zhuǎn)化[50]。
2.3 circRNA 介導(dǎo)的肝纖維化信號通路及其靶點circRNA 是由前體mRNA 反向剪接產(chǎn)生的一類內(nèi)源性ncRNA。circUbe2k 在CCl4誘導(dǎo)的肝纖維化小鼠和TGF-β1 刺激的LX-2 細(xì)胞中表達(dá)上調(diào),抑制其表達(dá)可以通過miR-149-5p/TGF-β2 軸抑制HSC 的活化和增殖[51]。circDIDO1 通 過 靶 向miR-141-3p 上 調(diào) 升 高PTEN 蛋白水平,降低LX2 細(xì)胞中p-AKT/AKT 的比例抑制HSC的活化[52]。
2.4 ncRNA 相關(guān)藥物與活性成分及其靶點 中藥生物堿單體川芎嗪在體內(nèi)和體外上調(diào)miR-145表達(dá)并抑制TGF-β/SMAD 信號通路發(fā)揮抗肝纖維化作用[53]。達(dá)沙替尼是第二代口服多靶點RTK抑制劑,上調(diào)miR-378-3p的表達(dá)抑制Wnt-10a,下調(diào)miR-17-5p升高WiF-1和SMAD-7 的表達(dá),通過抑制Wnt/β-catenin 和TGF-β/SMAD 通路抑制TAA 誘導(dǎo)的小鼠肝纖維化[54]。艾代拉里斯(Idelalisib)是首個高選擇性口服磷脂酰肌醇3-激酶δ(PI3K-δ)抑制劑,用于治療復(fù)發(fā)性慢性淋巴細(xì)胞白血病、復(fù)發(fā)性濾泡性淋巴瘤、復(fù)發(fā)性小淋巴細(xì)胞淋巴瘤。艾代拉里斯顯著降低了COL1和α-SMA 基因及蛋白水平的表達(dá),通過抑制TGF-β/SMAD3信號通路的活化抑制了HSC的活化,同時艾代拉里斯上調(diào)miR-124-3p的表達(dá)抑制PI3K/AKT通路促進細(xì)胞凋亡[55]。
3 小結(jié)
肝纖維化的進展涉及HSC、肝巨噬細(xì)胞在內(nèi)的多種細(xì)胞,通過多通路相互交聯(lián)作用,調(diào)節(jié)細(xì)胞活化、細(xì)胞自噬和細(xì)胞凋亡等多種形式。上文中提到的潛在活性成分及靶標(biāo)大多數(shù)處于前瞻性研究階段。近年來,隨著人們對肝纖維化認(rèn)識的逐步加深,已經(jīng)有越來越多的藥物及活性成分發(fā)現(xiàn)具有抗肝纖維化的潛力,新的肝纖維化調(diào)節(jié)方式也在逐漸被發(fā)現(xiàn)。目前,藥物治療仍是肝纖維化的主要治療方式。ACC 抑制劑Aramchol Meglumine、THR-β 激動劑Resmetirom、小分子半乳糖凝集素-3 抑制劑Belapectin 和GB1211、脂質(zhì)納米顆粒BMS-986263 等一批新藥已步入Ⅱ/Ⅲ期臨床研究,相信隨著對以往機制及新發(fā)現(xiàn)機制的深入剖析,一定能給藥物抗肝纖維化治療提供新的助力。
利益沖突聲明:本文不存在任何利益沖突。
作者貢獻(xiàn)聲明:周鑫負(fù)責(zé)文獻(xiàn)檢索,撰寫論文;王智、何雪茹、付裕豪、荀雪姣負(fù)責(zé)資料分析;李穎、董占軍負(fù)責(zé)指導(dǎo)撰寫文章并最終定稿。
猜你喜歡
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
媽媽寶寶(2019年10期)2019-10-26 02:45:34
中國生殖健康(2019年3期)2019-02-01 06:12:26
鐵道通信信號(2018年11期)2019-01-19 01:15:08
電子制作(2018年11期)2018-08-04 03:25:42
鐵道通信信號(2018年2期)2018-04-18 12:18:10
鐵道通信信號(2016年11期)2016-06-01 12:11:32
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
中國病理生理雜志(2015年8期)2015-12-21 12:38:06
