集胞藍細菌PCC 6803中非生物脅迫響應的核心基因分析
2023-12-19 05:17:02程亞蕊黃世鵬張天緣張建猛王鋒尖
高師理科學刊 2023年11期
程亞蕊,黃世鵬,張天緣,張建猛,王鋒尖
集胞藍細菌PCC 6803中非生物脅迫響應的核心基因分析
程亞蕊1,黃世鵬1,張天緣2,張建猛1,王鋒尖1
(1. 漢江師范學院 化學與環境工程學院,湖北 十堰 442000;2. 華中農業大學 生命科學技術學院,湖北 武漢 430070)
以集胞藍細菌PCC 6803的多組轉錄組數據為對象,鑒定藍細菌在非生物脅迫下共同表達的核心差異基因(CDGs)).結果表明,9個對氮、鐵、碳、磷缺乏響應的CDGs被鑒定.京都基因和基因組百科全書(KEGG)分析表明,氮代謝、ABC轉運蛋白和氨基酸代謝與營養缺乏有關.研究還鑒定了熱激處理下共同表達的CDGs,并且代謝通路ABC轉運蛋白和碳固定也與熱脅迫相關.同時構建了CDGs的蛋白質-蛋白質相互作用網絡,以預測基因在應對脅迫可能的響應機制.這些候選基因庫將有助于加速理解藍細菌在應激中的調控模式.
集胞藍細菌PCC 6803;非生物脅迫;核心差異基因;KEGG;蛋白互作
藍細菌屬于革蘭陰性的光合生物,通過捕獲太陽能來增強其高效的光自養代謝,進而固定無機碳和氮[1].藍細菌的光系統與高等植物相似,可在各種環境壓力下成功生存,因此其可以作為高等植物研究非生物脅迫下光合作用和信號轉導機制的優秀模型[2].由于集胞藍細菌PCC 6803(sp. PCC 6803)生長速度相對較快、基因組較小且易于修飾的特點,已成為研究各種應激響應的模式生物[3]71.組學測序技術已成功應用于研究藍細菌對脅迫的響應.前期研究通過整合轉錄組學、蛋白質組學和代謝組學數據,鑒定了集胞藍細菌對鹽脅迫響應的基因以及激活平衡滲透壓的基因[4].鹽濃度過高會破壞細胞內離子穩態,并導致活性氧的積累,進而導致光合系統的破壞和膜脂質的過氧化[5].藍細菌對3種不同光照(藍、橙、紅)適應過程的相關基因也通過轉錄組測序進行了鑒定[6].集胞藍細菌PCC 6803在缺鐵和氧化脅迫下的轉錄組數據也確定了該菌株在各種應激下的調控機制[7].
不同類型的脅迫不僅會導致藍細菌發生類似的生理變化,還可能誘導一系列相同基因的表達,并激活相關蛋白來響應非生物應激.基于不同的組學數據可以比較分析響應壓力源的共同基因和代謝途徑.集胞藍細菌應激的多轉錄組比較分析確定了其對己醇、苯甲醇和丁醇等響應的共同表達基因[8].多組氮缺乏的轉錄組數據的分析揭示了可能參與異形胞分化的新候選基因[9]2545.然而,尚未有多組轉錄組數據的比較分析來確定藍細菌對營養缺乏響應的共同基因和代謝途徑.
本文重新分析集胞藍細菌PCC 6803的3種不同營養限制和2種熱激處理轉錄組數據集,并確定細胞對應激因素的響應.通過生物信息學分析,確定脅迫響應的共同表達基因和代謝途徑,構建共同表達基因的蛋白質-蛋白質相互作用(protein-protein interaction,PPI)網絡,確定參與藍細菌抗脅迫的重要基因.這些候選基因庫將有助于加速理解藍細菌在應激中的調控機制,以期為藍細菌和植物對脅迫響應機制提供理論基礎.
1 材料與方法
1.1 轉錄組數據采集及分析
集胞藍細菌PCC 6803不同應激處理的轉錄組數據集從國家生物技術信息中心(NCBI,https://www.ncbi.nlm.nih.gov/sra)下載.本文共篩選4個不同的數據集(SRP102282,缺氮4 h,每個處理2個重復;SRP301698,缺鐵4 h,每個處理3個重復;SRP032228,缺碳20 h,缺磷12 h,42℃ 30 min,對照2個重復,其他無重復;SRP3328835,39℃ 1 h,每個處理3個重復),共21個樣本組成.通過FastQC處理原始數據獲得高質量的數據.Bowtie2用于將高質量數據與集胞藍細菌PCC 6803參考基因組進行比對[10].通過比較應激和對照樣品中的基因表達水平來鑒定共同表達的核心差異基因(core differential genes,CDGs).
1.2 CDGs的富集分析
通過比較不同數據集中的差異表達基因(倍數變化≥2.00和<0.05),并基于維恩圖分析應激響應的CDGs,然后對這些基因進行基因本體論(Gene Ontology,GO)、京都基因和基因組百科全書(Kyoto Encyclopedia of Genes and Genomes,KEGG)富集分析,以確定與營養缺乏和熱激有關的代謝途徑.使用clusterProfiler 3.8版進行GO和KEGG富集分析.
1.3 PPI網絡分析
基于STRING數據庫(https://string-db.org/cgi/input.pl)可以對已知和未知的蛋白質-蛋白質關聯數據進行收集和整合[11].可能的相互關系參考的標準是置信度得分為0.4,交互因素的最大數量為0.交互預測主要來自文本挖掘、實驗、數據庫、共表達、鄰域、基因融合和共出現.
2 結果與分析
2.1 與營養缺乏相關的CDGs鑒定
首先分析在氮、鐵、碳、磷缺乏下的轉錄組數據集.在營養缺乏的情況下,不同數據集的總體基因表達譜(見圖1a)是相似的,表明集胞藍細菌對脅迫的反應是相容的.由維恩圖(見圖1b)可見,在不同的樣品中共鑒定出1 132個差異表達基因,但不同數據集僅鑒定了9個在不同營養限制中保守的CDGs.這9個基因分別編碼缺鐵誘導的葉綠素結合蛋白、Fe2+轉運蛋白、50S核糖體蛋白L4、異天冬氨酰肽酶和未知蛋白等(見表1).每個數據集特有的數百個基因可能是由菌株對不同應激處理和生長條件的耐受水平差異引起的.

圖1 與營養缺乏相關CDGs的鑒定
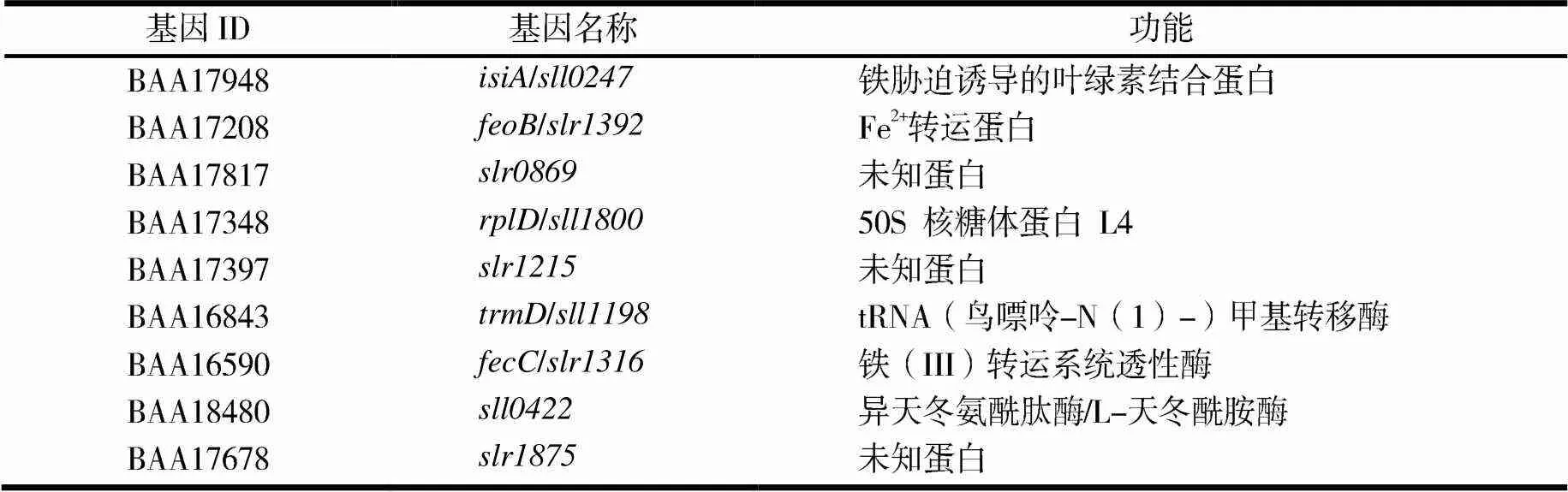
表1 不同數據集中響應營養限制的CDGs
對在不同數據集中檢測共同表達的CDGs進行了GO和KEGG富集分析(見圖2).在鐵和氮缺乏的情況下,對46個CDGs的GO分析(見圖2a)表明,所富集的生物過程與膜的組成、類囊體膜和質膜有關.KEGG分析(見圖2b)表明,ABC轉運蛋白是最顯著富集的代謝通路,其次是氮代謝,分別有35%,18%的CDGs參與這些途徑,表明轉運蛋白在營養缺乏的情況下發揮著關鍵作用.在氮、碳、磷缺乏的情況下,對48個CDGs的KEGG富集分析(見圖2c)表明,核糖體是最顯著的生物過程,其次是氨基酸代謝.在鐵、碳、磷缺乏的情況下,CDGs主要富集在ABC轉運蛋白和丙氨酸、天冬氨酸和谷氨酸代謝(見圖2d).這些結果表明,氨基酸代謝可能與營養限制有關,因為菌株需要更多的氨基酸來合成蛋白質以應對營養缺乏.
2.2 響應溫度的CDGs鑒定
對轉錄組數據集SRP332228(42℃,30 min)和SRP3328835(39℃,1 h)進行比較分析,以探究與溫度相關的基因.35個共同表達的CDGs被檢測(見圖3a).由缺氮、缺鐵和熱激下的CDGs(見圖3b)可見,該結果僅檢測到3個共同表達的CDGs,包括BAA17923(,未知蛋白)、BAA16628(,ABC轉運蛋白)和BAA16842(,鐵攝取蛋白A1),其在碳和磷缺乏下并未差異表達.該結果表明,在不同脅迫處理下,共有基因較少,可能是由于藍細菌本身的基因數量較少及不同數據集中菌株的生長狀態和脅迫處理不同.GO分析(見圖3c)主要富集在細胞質、ATP結合和蛋白水解.KEGG分析(見圖3d)顯示,ABC轉運蛋白是最富集的代謝途徑,其次是碳固定,該結果與營養缺乏下的分析一致,表明ABC轉運蛋白可能在各種脅迫下均發揮重要作用,這可能是由于菌株在應激時會增強營養物質的吸收和運輸.

圖3 與熱激相關CDGs的鑒定
2.3 CDGs的PPI網絡分析
為了探究在營養缺乏下9個共有表達CDGs的功能,基于STRING數據庫中尋找這些基因的PPI信息,每個PPI網絡由11個節點組成.部分CDGs的蛋白互作網絡見圖4.與IsiA相關的蛋白質是光系統I的亞基,與RplD相關性較高的蛋白質是核糖體亞基;與FecC相關性較高的蛋白主要與鐵轉運有關;與未知蛋白質(Sll0422,Slr0869,Slr1726,Slr1113)相互作用的蛋白質主要是未知蛋白質.這說明應對脅迫的基因并不是獨立發揮作用,而是與其具有類似功能的基因共同作用來抵抗不利條件.

圖4 CDGs的PPI網絡分析
3 討論
本文通過比較不同脅迫下的轉錄組數據集,以確定藍細菌對應激響應的共有基因和代謝途徑.與植物相比,集胞藍細菌的基因數量相對較少,只有3 000多個[3]1173.因此,本文通過比較轉錄組分析確定的共有表達的CDGs較少.念珠藻PCC 7120中通過將2組轉錄組數據進行比較分析來篩選與異形胞分化相關的基因,但該研究并未使用系統生物信息學的分析方法[9]2555.前期在集胞藍細菌中也未有相關的比較分析來確定其在非生物脅迫下的共有核心基因.
在營養缺乏的情況下,CDGs富集在ABC轉運蛋白、氮代謝和氨基酸代謝中.集胞藍細菌PCC 6803基因組中有50多個基因編碼ABC轉運蛋白,其功能主要涉及各種化合物和營養物質的運輸[12].植物在營養限制下會誘導蛋白質降解,激活呼吸途徑,并利用氨基酸作為替代底物來應對能量不足.氨基酸也可以作為信號分子來協調植物生長和脅迫響應[13],藍細菌也可能具有與植物類似的響應機制.在氮和鐵缺乏下,CDGs也富集到氮代謝.鐵攝取調節因子FurA參與念珠藻PCC 7120中的異形胞分化[14].總氮調節因子NtcA可以控制基因的表達以調節鐵的獲取,從而保護藍細菌免受缺鐵引起的氧化應激的影響[15].這表明藍細菌中的鐵和氮調節網絡存在相互交叉.響應溫度的CDGs的KEGG分析主要富集在ABC轉運蛋白和碳固定途徑.為了應對低溫脅迫導致的光合速率降低,香蕉會上調磷酸烯醇丙酮酸羧化酶,以提高固碳效率[16].因此,在不利條件下生長的植物可能會導致光合效率下降,并通過調節碳固定來應對能量不足.
PPI網絡顯示與IsiA高度相關的蛋白質均為PS I亞單位.基因與光系統II的具有序列相似性,前者參與鐵缺乏、氧化脅迫、熱激或鹽脅迫下的調節[17-19].這表明藍細菌中IsiA在脅迫下的調控發揮重要作用.大多數FeoB相關蛋白屬于ABC轉運蛋白,其在N-末端具有典型的ATP結合基序,其基因的缺失導致細胞利用亞鐵的能力顯著降低.Ssr2333是一種未知的蛋白質,但與FeoB的相關性最高,這表明Ssr2333可能也屬于ABC轉運蛋白.未知蛋白Sll0422和CphB(藻青素酶)具有最高的相關性,CphB可以水解-Asp-Arg鍵,其也與細菌的碳利用有關[20].這一系列基因在集胞藍細菌PCC 6803應對脅迫時的調控機制還有待進一步的驗證.因此,應構建并篩選基因的缺失和過表達菌株,觀察其在不同脅迫下的生長,進而判斷基因對藍細菌抗逆性的影響.若突變體的表型有明顯變化,則可通過轉錄組測序來進一步確定該基因在各脅迫下的調控機制.
綜上所述,本文從集胞藍細菌PCC 6803的不同轉錄組數據集中鑒定了響應不同非生物脅迫的CDGs.KEGG分析顯示,ABC轉運蛋白、氮代謝、氨基酸代謝和碳固定等代謝途徑與非生物脅迫相關.本文鑒定了9個在營養缺乏下共同表達的CDGs,并基于PPI網絡分析揭示了可能在藍細菌應激反應作為重要調節因子的假定蛋白質.此研究將為了解藍細菌對非生物應激反應的通用信號奠定堅實的基礎.
[1] Veaudor T,Blanc-garin V,Chenebault C,et al.Recent advances in the photoautotrophic metabolism of cyanobacteria:biotechnological implications[J].Life,2020,10(5):71.
[2] Jensen P E,Leister D.Cyanobacteria as an experimental platform for modifying bacterial and plant photosynthesis[J].Frontiers in Bioengineering and Biotechnology,2014(2):7.
[3] Kaneko T,Tabata S.Complete genome structure of the unicellular cyanobacteriumsp. PCC 6803[J].Plant and Cell Physiology,1997,38(11):1171-1176.
[4] Kl?hn S,Mikkat S,Riediger M,et al.Integrative analysis of the salt stress response in cyanobacteria[J].Biology Direct,2021,16(1):26.
[5] YANG Z,LI J L,LIU L N,et al.Photosynthetic regulation under salt stress and salt-tolerance mechanism of sweet sorghum[J].Frontiers in Plant Science,2020,10:1722.
[6] Luimstra V M,Schuurmans J M,Hellingwerf K J,et al.Blue light induces major changes in the gene expression profile of the cyanobacteriumsp. PCC 6803[J].Physiologia Plantarum,2020,170(1):10-26.
[7] CHENG Y,ZHANG T,WANG L,et al.Transcriptome analysis reveals IsiA-regulatory mechanisms underlying iron depletion and oxidative-stress acclimation insp. strain PCC 6803[J].Applied and Environmental Microbiology,2020,86(13):e00517-e00520.
[8] Mironov K S,Kupriyanova E V,Shumskaya M,et al.Alcohol stress on cyanobacterial membranes:New insights revealed by transcriptomics[J].Gene,2021,764:145055.
[9] Brenes-álvarez M,Mitschke J,Olmedo-verd E,et al.Elements of the heterocyst-specific transcriptome unravelled by co-expression analysis insp. PCC 7120[J].Environmental Microbiology,2019,21(7):2544-2558.
[10] Langmead B,Salzberg S L.Fast gapped-read alignment with Bowtie 2[J].Nature Methods,2012,9(4):357-359.
[11] Szklarczyk D,Franceschini A,Wyder S,et al.STRING v10:protein-protein interaction networks,integrated over the tree of life[J].Nucleic Acids Research,2015,43:D447-D452.
[12] Shvarev D,Maldener I.ATP-binding cassette transporters of the multicellular cyanobacteriumsp. PCC 7120:a wide variety for a complex lifestyle[J].FEMS Microbiology Letters,2018,365(4):fny012.
[13] Heinemann B,Hildebrandt T M.The role of amino acid metabolism in signaling and metabolic adaptation to stress-induced energy deficiency in plants[J].Journal of Experimental Botany,2021,72(13):4634-4645.
[14] González A,Bes M T,Peleato M L,et al.Expanding the role of FurA as essential global regulator in cyanobacteria[J].PLoS One,2016,11(3):e0151384.
[15] Kaushik M S,Srivastava M,Singh A,et al.Impairment ofgene revealed its role in regulating iron homeostasis,ROS production and cellular phenotype under iron deficiency in cyanobacteriumsp. PCC 7120[J].World Journal of Microbiology & Biotechnology,2017,33(8):158.
[16] LIU J,Taká? T,YI G,et al.Acceleration of carbon fixation in chilling-sensitive banana under mild and moderate chilling stresses[J].International Journal of Molecular Sciences,2020,21(23):9326.
[17] Kojima K,Suzuki-Maenaka T,Kikuchi T,et al.Roles of the cyanobacterial isiABC operon in protection from oxidative and heat stresses[J].Plant Physiology,2006,128(3):507-519.
[18] MA F,ZHANG X,ZHU X,et al.Dynamic changes of IsiA-containing complexes during long-term iron deficiency insp. PCC 6803[J].Molecular Plant,2017,10(1):143-154.
[19] Yousef N,Pistorius E K,Michel K P.Comparative analysis of idiA and isiA transcription under iron starvation and oxidative stress inelongatus PCC 7942 wild-type and selected mutants[J].Archives of Microbiology,2003,180(6):471-483.
[20] Hejazi M,Piotukh K,Mattow J,et al.Isoaspartyldipeptidase activity of plant-type asparaginases[J].Biochemical Journal,2002,364(1):129-136.
Analysis of core genes associated with abiotic stress response insp. PCC 6803
CHENG Yarui1,HUANG Shipeng1,ZHANG Tianyuan2,ZHANG Jianmeng1,WANG Fengjian1
(1. School of Chemistry and Environmental Engineering,Hanjiang Normal University,Shiyan 442000,China;2. School of Life Science and Technology,Huazhong Agricultural University,Wuhan 430070,China)
Multiple sets of transcriptome datasets ofsp. PCC 6803 was applied to identify core differential genes(CDGs)under abiotic stress.Results showed that 9 co-expressed CDGs responsive to nitrogen,iron,carbon and phosphorus deficiency were identified.The Kyoto Encyclopedia of Genes and Genomes(KEGG)analysis showed that nitrogen metabolism,ABC transporters,and amino acid metabolism were related to nutrient deficiency.The co-expressed CDGs were also identified under heat shock,the ABC transporters and carbon fixation were also related to heat stress.At the same time,a protein-protein interaction network of CDGs was constructed to predict the possible response mechanism of genes in response to stress.These candidate gene libraries will help accelerate the understanding of the regulatory patterns of cyanobacteria under various stresses.
sp. PCC 6803;abiotic stress;core differential genes;KEGG;protein-protein interaction
1007-9831(2023)11-0050-06
Q93
A
10.3969/j.issn.1007-9831.2023.11.010
2023-07-08
漢江師范學院重點項目(XJ2022A02);湖北省大學生創新創業訓練計劃項目(S202210518023);漢江師范學院一般項目(XJ2022030)
程亞蕊(1994-),女,河南信陽人,講師,博士,從事微生物多樣性和生物信息學研究.E-mail:chengyarui@hjnu.edu.cn
王鋒尖(1976-),男,江西高安人,教授,博士,從事微生物生態學研究.E-mail:13972500499@163.com
猜你喜歡
今日農業(2021年11期)2021-08-13 08:53:34
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
中學生數理化·七年級數學人教版(2019年10期)2019-11-25 07:33:58
中國生殖健康(2019年8期)2019-01-07 01:18:24
幸福(2018年33期)2018-12-05 05:22:46
電子制作(2018年18期)2018-11-14 01:48:24
中學生數理化·高一版(2018年9期)2018-10-09 06:46:50
湖南教育·C版(2018年3期)2018-06-05 16:54:36
山東工業技術(2016年15期)2016-12-01 05:31:22
海峽姐妹(2016年5期)2016-02-27 15:20:20
