飼料磷脂水平對烏鱧幼魚生長性能、抗氧化能力及脂肪沉積的影響
2023-12-21 16:22:18李文飛王振杰陳乃松李松林
廣東海洋大學學報 2023年6期
關鍵詞:質量
張 玉,龔 埜,李文飛,王振杰,陳乃松,2,李松林,2
(1.上海海洋大學農業農村部魚類營養與環境生態研究中心,上海 201306;2.上海海洋大學水產科學國家級實驗教學示范中心,上海 201306)
磷脂(Phospholipid,PL)是魚類體內重要的結構和功能分子,參與細胞膜合成、能量代謝、信號轉導等多種生理過程,可通過促進脂肪酸運轉、增強脂肪酸氧化并抑制脂質生成等方式降低肝臟脂肪含量[1-2]。在飼料中添加適量磷脂可促進魚類的生長、存活、抗氧化和骨骼發育等方面的表現[3-5]。魚類對磷脂的需求量和利用效率會因不同發育階段和不同物種而變化。對于仔稚魚,由于其合成磷脂的能力有限,在其飼料中補充磷脂十分必要[6],添加適宜的磷脂可促進仔稚魚生長發育、提高存活率并降低骨骼畸形率[7-10]。而高水平的脂質攝入可能會導致魚類肝臟中脂質過度沉積[11],影響肝功能和其健康狀況[12-15]。飼料中添加磷脂可降低大黃魚(Larimichthys crocea)[16-17]、雜交鱧(Channa argus×Channa maculata)[18]和尼羅羅非魚(Oreochromis niloticus)[19]的肝臟脂肪含量。飼料中添加磷脂會顯著提高杜氏(Serioladumerili)[20]和斑點叉尾鮰(Ictalurus punctatus)[21]肝臟中的脂質積累。飼料磷脂的增加減少了脂肪酸氧化,增加了雜交石斑魚(Epinephelus fuscoguttatus♀×Epinephelus lanceolatus♂)肝臟脂質的積累[22]。因此,探討磷脂對魚類脂質代謝的調節機制,優化魚類飼料中磷脂的添加水平和比例,對于提高水產養殖效率有著重要意義。
烏鱧(Channa argus)是中國北方廣泛養殖的肉食性魚類之一,有很高的市場價值和需求[23],但在烏鱧的集約化養殖中,時常出現營養性代謝疾病,不利于烏鱧養殖產業化健康發展[24-25]。目前,有關烏鱧幼魚的營養需求和生理代謝研究較多[18,26-28],但烏鱧幼魚的磷脂需求及調節機制尚不清楚。本研究探討飼料中添加磷脂對烏鱧幼魚的生長、抗氧化以及脂質代謝的影響,分析烏鱧幼魚對磷脂的需求,旨在為烏鱧的健康養殖提供參考。
1 材料與方法
1.1 實驗飼料
在基礎配合飼料中分別添加質量分數0%、2%、4%、6%和8%的磷脂(各組飼料中磷脂水平實測值分別為1.37%、3.22%、5.36%、7.23%和9.15%),通過調整大豆油水平來維持脂肪含量的穩定,配制成5 種等氮等脂飼料,分別記為PL0、PL2、PL4、PL6 和PL8(表1)。實驗飼料氨基酸組成見表2。固體原料粉碎,過孔徑180 μm的篩,逐級混勻,與磷脂充分混合后再與魚油和豆油的混合物充分混勻。將混合的原料加水充分混勻后擠壓制粒,在60 ℃下烘干、密封,于-20 ℃保存備用。
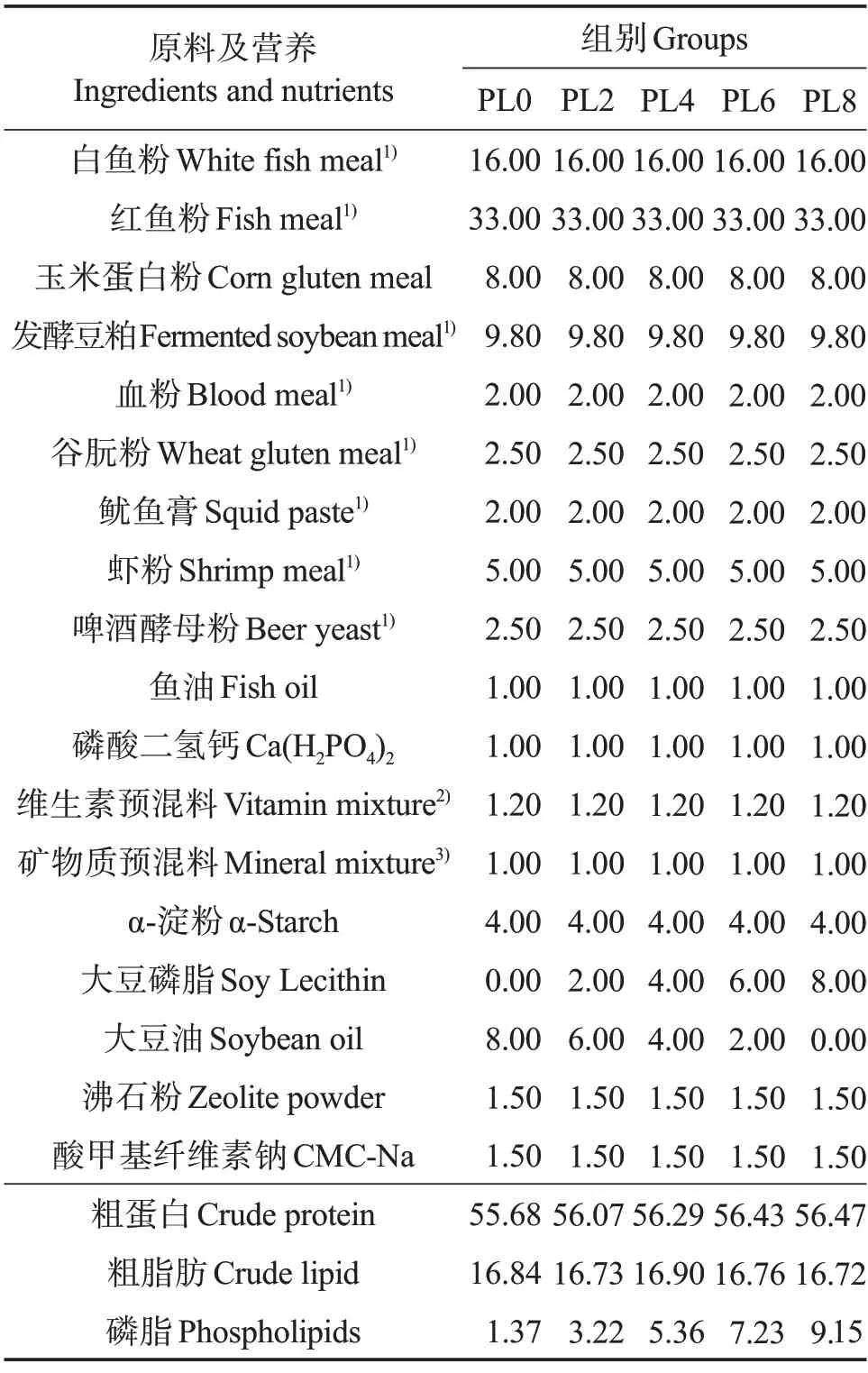
表1 實驗飼料配方及營養組成(干物質)Table 1 Formulation and chemical composition of experimental diets(dry matter) %
1.2 養殖管理
養殖實驗在上海海洋大學和上海農好飼料股份有限公司聯合實驗室的室內循環水養殖系統中進行。實驗魚用商品飼料(浙江欣欣天恩水產飼料有限公司)馴化2 周,停飼24 h,挑選規格相似、體格健壯的烏鱧幼魚[初始體質量(3.99 ± 0.01)g]隨機均分到15 個容量為800 L 的養殖桶中,每個養殖桶45 尾。各組每天6:00、10:00、14:00 和17:00 進行表觀飽食投喂,為期43 d。實驗用水經海綿和珊瑚砂過濾,并用紫外線消毒24 h,幼魚攝食后吸底和排污。養殖實驗期間采用自然光照周期,水溫(26 ±1)℃,溶解氧大于6.2 mg/L,pH 為7.2 ± 0.2,氨氮和亞硝酸氮質量濃度均≤0.1 mg/L。
1.3 幼魚生長性能統計及樣品采集
養殖實驗結束后,幼魚停飼24 h,用丁香酚麻醉,計數,稱總質量,計算生長性能指標:
存活率(SR,%)=終末數量/初始數量;
增重率(WGR)=(末均體質量-初均體質量)/初均體質量;
特定生長率(SGR)=(ln 末均體質量-ln 初均體質量)/養殖周期;
飼料系數(FCR)=攝食飼料質量/(末均體質量-初均體質量);
日攝食率(FI)=攝食飼料質量/[(初體質量+末體質量)/(2×養殖周期)];
臟體比(VSI)=內臟團質量/末體質量;
肝體比(HSI)=肝臟質量/末體質量。
每桶隨機取魚20 尾,6 尾保存在-20 ℃用于體成分分析。剩余14 尾魚中,6 尾用于測量肝體比和臟體比,8 尾剖取肝臟和腸道用于采集生化分析和基因檢測,所有樣品采集后于液氮中速凍,于-80 ℃條件下保存備用。
1.4 樣品分析
1.4.1 樣品及實驗飼料常規指標分析 實驗飼料和樣品(包括全魚、肌肉和肝臟)的水分含量采用105 ℃恒溫干燥法(GB/T6435—2014)測定,烘干樣用于測定粗脂肪、粗蛋白及粗灰分;粗脂肪含量采用氯仿-甲醇萃取法測定[29];粗蛋白質含量采用凱氏定氮儀(OPSIS,瑞典)測定(GB/T5009.124—2016);灰分含量通過將樣品在550 ℃馬弗爐中燃燒至恒重后測定(GB/T6438—2007)。磷脂含量通過鉬藍法測定[30]。脂肪酸相對含量(占總脂肪酸質量分數)用HP6890 氣相色譜儀(Agilents Technologies,Santa-Clara,CA,USA)測定,用于脂肪酸檢測的飼料、肌肉和肝臟樣品經過前處理變成的甲酯得到。
1.4.2 生化指標測定 取適量-80 ℃保存的肝臟樣品于離心管,加入生理鹽水,研磨,離心,取上清液,根據南京建成試劑盒說明書測定總蛋白(TP)含量、丙二醛(MDA)含量、超氧化物歧化酶(SOD)活力、過氧化氫酶(CAT)活力和總抗氧化能力(T-AOC)指標。
1.4.3 RNA 提取和實時定量PCR 使用TransZolUp(TransGen)提取肝臟樣品的總RNA,用PrimeScript?RT 試劑盒(Takara)將其逆轉錄成第1 鏈cDNA。用TransStart?TopGreenqPCR SuperMix(TransGen)在定量熱循環儀(Mastercycler EP Realplex,Eppendorf)中進行實時定量PCR。通過Primer Premier5.0設計特異性引物(表3),以經過驗證穩定的β-actin為內參基因,根據2-ΔΔCt方法計算基因相對表達量[28]。

表3 引物序列Table 3 Sequence of primers
1.5 數據統計與分析
結果表示為平均值± 標準誤。使用SPSS26.0統計軟件對數據進行單因素方差分析,數據差異顯著時,采用Duncan's多重檢驗法進行比較,顯著水平α=0.05。
2 結果
2.1 飼料磷脂水平對烏鱧幼魚生長性能與飼料利用的影響
由表4 可知,在飼料中添加磷脂對烏鱧幼魚的終末體質量(FBW)和SGR 有顯著影響,其中PL4組和PL6組均顯著高于PL0組(對照)(P<0.05),但對幼魚SR影響不顯著(P>0.05)。隨飼料中磷脂水平的升高,幼魚HSI 無顯著差異(P>0.05),PL8 組幼魚VSI 顯著小于其他組(P<0.05);FI 和FCR 先降低后升高,PL2 和PL4 組FI 顯著低于PL0 組(P<0.05),PL2組、PL4組和PL6組FCR 顯著低于PL0組(P<0.05)。
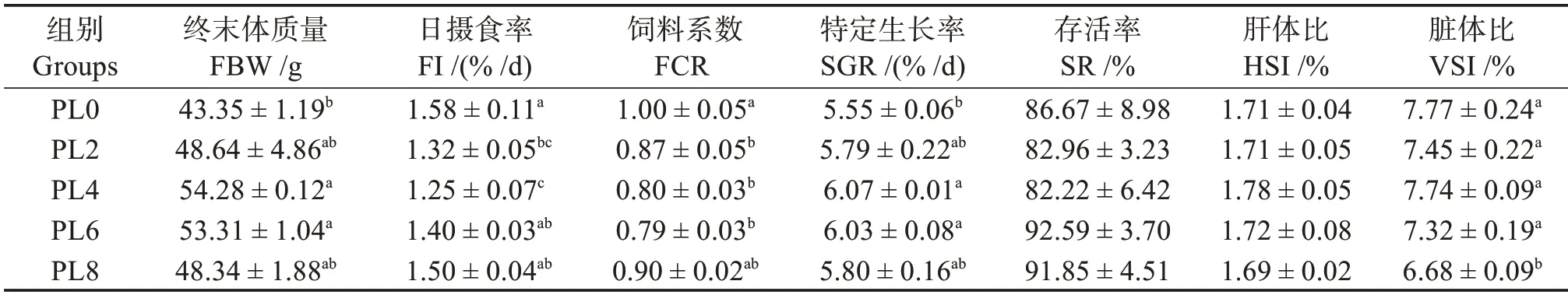
表4 飼料磷脂水平對烏鱧幼魚的生長性能與飼料利用情況的影響Table 4 Effects of dietary phospholipids on growth performance and feed utilization of juvenile snakehead
2.2 飼料磷脂水平對烏鱧幼魚的魚體、肝臟、肌肉營養成分的影響
由表5可知:隨著飼料中磷脂添加量的增加,魚體的粗脂肪含量呈下降趨勢,PL6 組和PL8 組顯著低于PL0 組(P<0.05),魚體水分含量則呈上升趨勢,PL6 組和PL8 組顯著高于PL0 組(P<0.05);肌肉粗蛋白含量呈下降趨勢,PL8 組肌肉蛋白含量顯著低于PL0組(P<0.05);肌肉水分呈先降后升變化趨勢,PL2 組和PL4 組顯著低于PL0 組,PL8 組升至PL0 組水平(P<0.05)。磷脂水平的提高,對幼魚肝臟組織中的蛋白含量無顯著影響(P>0.05),肝臟粗脂肪含量呈降低趨勢,PL0 組肝臟脂肪含量顯著高于其他處理組(P<0.05);肝臟水分含量則呈升高趨勢,PL6組、PL8組顯著高于PL0組(P<0.05)。
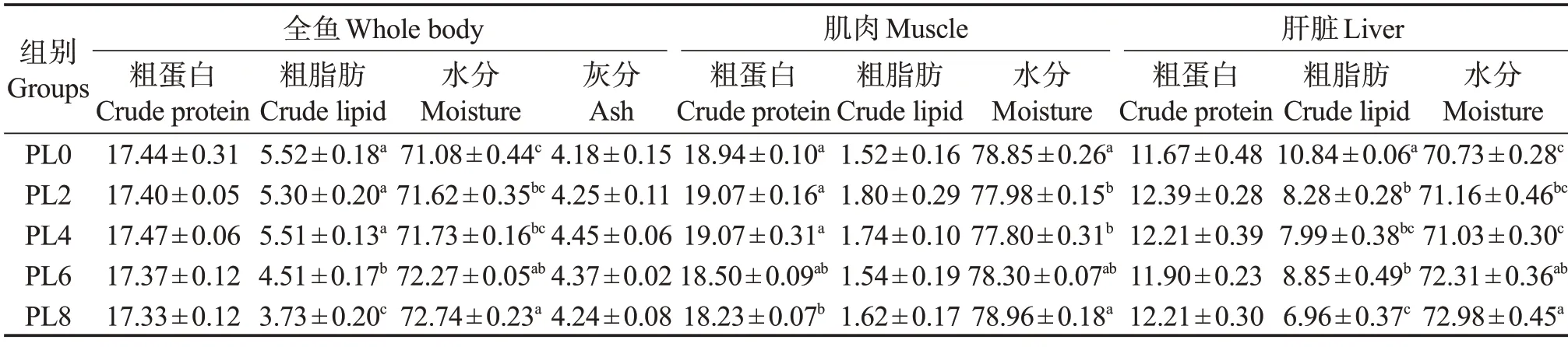
表5 飼料磷脂水平對烏鱧幼魚魚體、肌肉和肝臟成分的影響(濕基)Table 5 Effects of dietary phospholipids on composition of whole fish body,muscle and liver of juvenile snakehead(wet basis) %
2.3 飼料磷脂水平對烏鱧幼魚肝臟抗氧化酶活力的影響
由表6可知:隨著磷脂含量的升高,烏鱧幼魚肝臟MDA 含量呈降低趨勢,PL6和PL8組顯著低于其他組(P<0.05);肝臟SOD 活力呈升高趨勢,PL6 和PL8 組顯著高于PL0 組(P<0.05),PL8 組升至最高(P<0.05);T-AOC 則呈先升后降趨勢,PL2 組最高(P<0.05),PL8 組降至對照組水平。飼料中磷脂含量變化對CAT活力無顯著影響(P>0.05)。
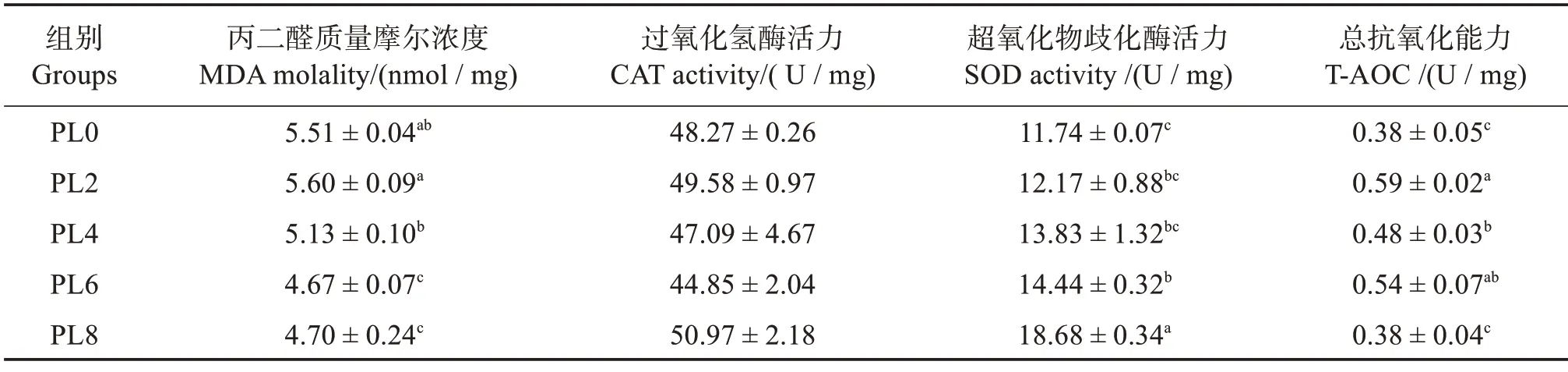
表6 飼料磷脂水平對烏鱧幼魚肝臟抗氧化酶活力的影響Table 6 Effects of dietary phospholipids on hepatic antioxidant capacity of juvenile snakehead
2.4 飼料磷脂水平對烏鱧幼魚肝臟、肌肉脂肪酸組成的影響
由表7可知:隨著飼料中磷脂含量的升高,烏鱧幼魚肝臟組織脂肪酸C14:0、C18:0、C16:1n7、C18:1n9和單不飽和脂肪酸(MUFA)相對含量無顯著變化(P>0.05);C16:0、飽和脂肪酸(SFA)、C20:1n9、C20:4n-6、C20:5n-3、C22:6n-3、n-3 多不飽和脂肪酸(n-3 PUFA)相對含量顯著升高(P<0.05),且PL8組最高;C18:2n-6c、n-6 PUFA 和C18:3n-3 相對含量則顯著降低(P<0.05),且PL8組最低。
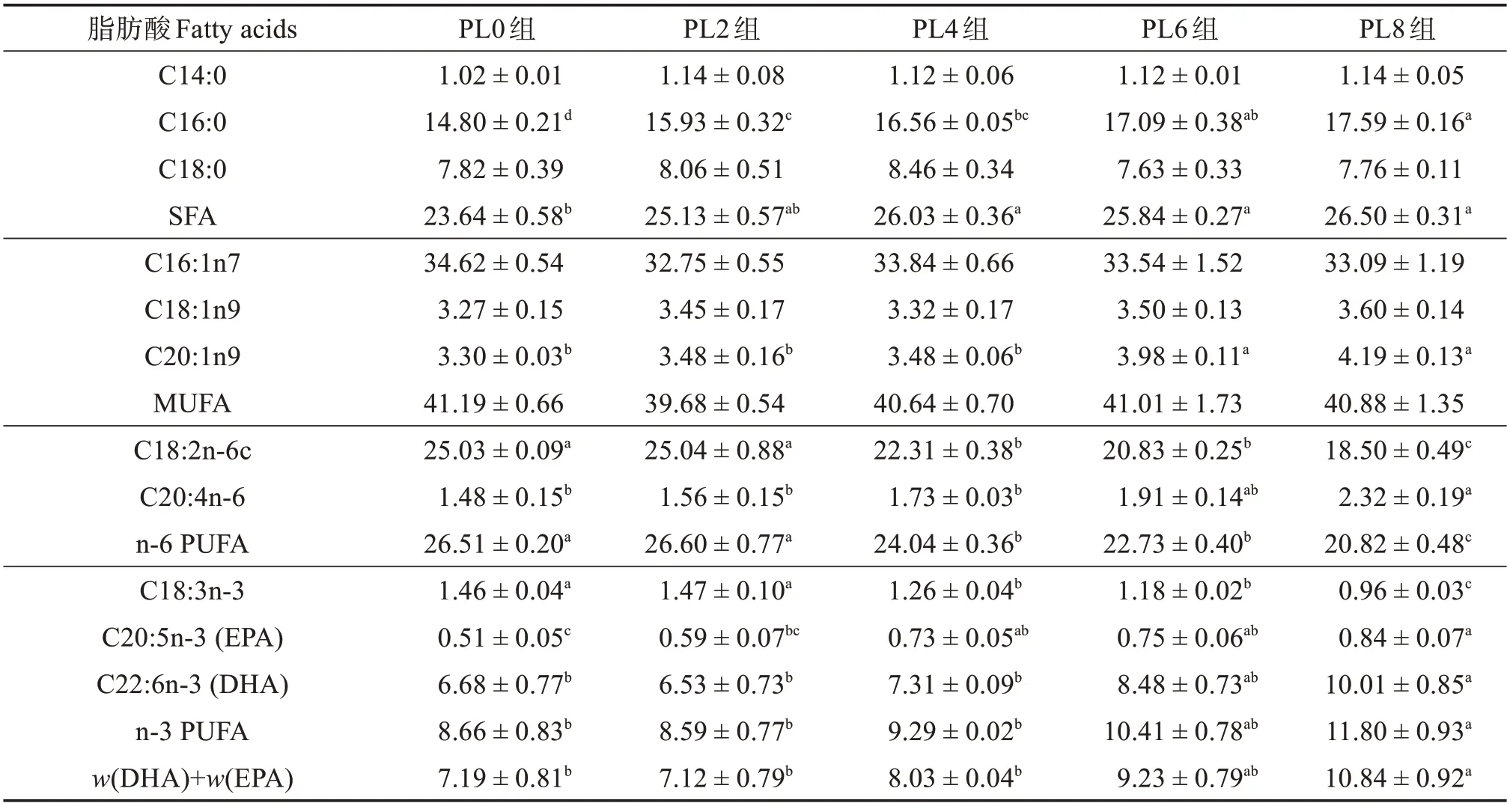
表7 各組烏鱧幼魚肝臟脂肪酸占總脂肪酸質量分數Table 7 Mass fraction of fatty acid to total fatty acid in liver of juvenile snakehead in different groups %
由表8 可知:在肌肉組織中,隨著飼料中磷脂含量的升高,C14:0、C18:1n9、C20:1n9 相對含量無顯著變化(P>0.05);但C16:0、C18:0、SFA、C20:4n-6、C20:5n-3、C22:6n-3、n-3 PUFA 相對含量值顯著升高(P<0.05),且PL8 組最高;C16:1n7、MUFA、C18:2n-6c、n-6 PUFA 和C18:3n-3 相對含量顯著降低(P<0.05),且PL8組最低。
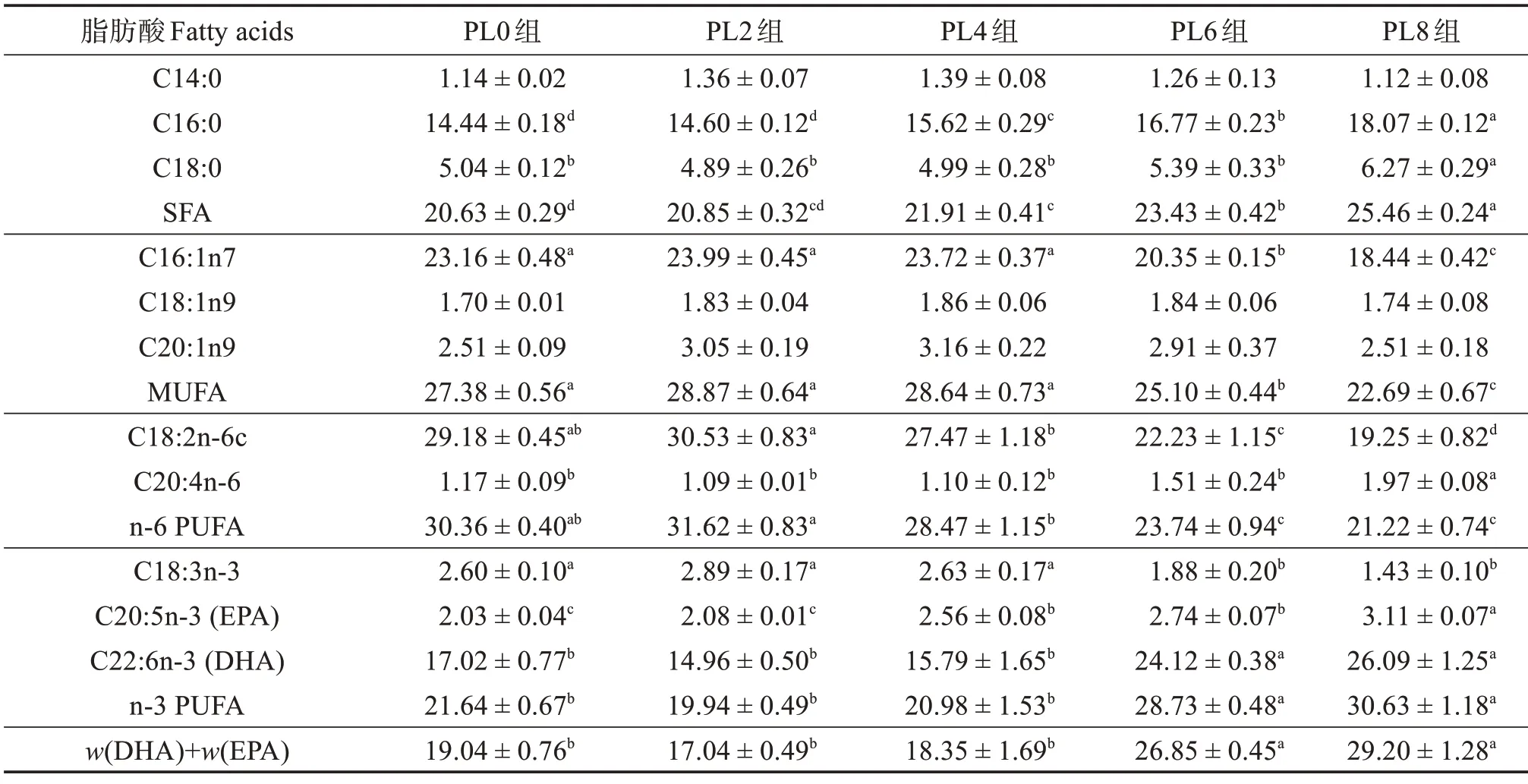
表8 不同飼料組烏鱧幼魚肌肉脂肪酸占總脂肪酸質量分數Table 8 Mass fraction of fatty acid to total fatty acids in muscle of juvenile snakehead in different groups %
2.5 飼料磷脂水平對烏鱧幼魚肝臟脂質代謝相關基因表達的影響
在脂肪生成相關基因中,飼料中磷脂水平的升高對fasn表達量無顯著影響(P>0.05);但顯著降低acc1的表達量,PL8 組顯著低于PL0 組(P<0.05)(圖1(a-b))。在脂肪分解相關基因中,PL4 組atgl基因表達量顯著高于其他組(P<0.05),PL0組mgl的表達量顯著低于除PL2 組外的各組(P<0.05),PL6 組的hsl表達量顯著高于其他組(P<0.05)(圖1(c-e))。在脂肪酸氧化相關基因中,PL6 組的aco表達量顯著高于PL0 組(P<0.05),同時,PL6 組cpt1和pparα的表達量顯著高于其他組(P<0.05)(圖1(f-h))。
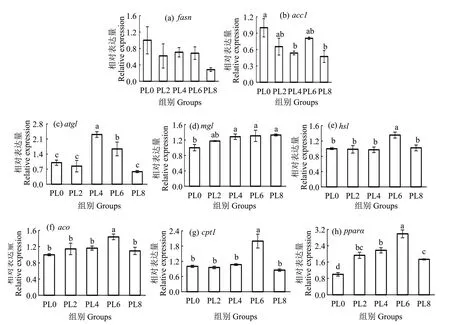
圖1 飼料磷脂水平對烏鱧肝臟脂質代謝相關基因表達的影響Fig.1 Effects of dietary phospholipids on relative expression of lipid metabolism related genes in liver of juvenile snakehead
3 討論
3.1 飼料磷脂對烏鱧幼魚生長的影響
磷脂是一種重要的結構性和功能性脂質[3]。魚類在幼體階段內源性磷脂合成能力有限[5],主要是腸上皮細胞的磷脂生物合成能力低,導致其對外源性磷脂的依賴性高[31]。磷脂可以增強腸道健康狀態,促進脂質從腸道轉運到身體其他部位,從而增加生長所需能量和促進脂肪酸的合成和沉積[32]。隨著魚類生長和發育,其對磷脂的需求量逐漸減少[4]。飼料磷脂對生長性能的影響在魚類生長早期階段更為明顯[17,33],為探究磷脂對烏鱧幼魚生長性能的影響,本研究實驗周期選擇為43 d。
在飼料中添加磷脂不會對大西洋鮭(Salmo salar)幼魚產生影響[34],大豆卵磷脂也不影響斑點叉尾鮰幼魚的生存和生長[20]。本研究中,隨著飼料中外源性磷脂含量的提高,烏鱧幼魚FBW 和SGR 顯著升高,FCR 顯著降低,可見添加適量的磷脂可增強烏鱧幼魚的食欲和飼料利用率,促進其生長發育。在牙鲆(Paralichthys olivaceus)[35]、虹鱒(Oncorhynchus mykiss)[33]、團頭魴(Megalobrama amblycephala)[36]和軍曹魚(Rachycentron canadum)[37]幼魚中也有類似結果。然而,當飼料中磷脂水平高于5.36%時,烏鱧幼魚生長性能開始下降,大黃魚[17]和雜交鱧[18]有類似結果,可能歸因于過量的磷脂造成營養失衡或者消化吸收障礙。
3.2 飼料磷脂對烏鱧幼魚脂質代謝的影響
在哺乳動物中,磷脂可通過減少脂肪酸的合成、促進脂肪酸氧化來減少肝臟脂肪的沉積[1]。在團頭魴[12]和大黃魚[38]等硬骨魚的研究中可證明脂質代謝相關基因在調節脂肪沉積中的作用。磷脂會通過抑制脂肪酸的合成,并促進脂肪酸氧化,來抑制某些魚類的脂質積累,如大黃魚幼魚[16]和雜交鱧[18]。本研究中,隨著飼料中大豆磷脂添加量的增加,烏鱧幼魚魚體和肝臟脂質含量顯著降低,說明磷脂可減少肝臟的脂肪沉積,在大口黑鱸(Micropterus salmoides)幼魚[39]中有相同結果。同時全魚和肝臟的水分含量顯著升高,魚的體脂百分比與體水分之間呈現很強的負相關,并且不受進食、生長和性腺發育的影響[40],在羅非魚[19]和牙鲆[20]中也發現了這種現象。此外,本研究檢測了烏鱧幼魚肝臟與脂肪生成(acc1、fasn)、脂肪分解(agt1、mgl和hsl)及脂肪氧化(aco、pparα和cpt1)相關的基因表達情況,結果顯示,隨著飼料中磷脂水平的增加,acc1基因的表達量顯著下降,而agt1、mgl、hsl、aco、pparα和cpt1基因的表達量均有不同程度的上升,表明磷脂能夠抑制烏鱧幼魚肝臟中脂肪生成途徑的活性,同時激活其脂肪分解或氧化途徑的活性,從而減少體內脂肪的積累。
3.3 飼料磷脂對烏鱧幼魚抗氧化能力的影響
過多的肝臟脂質沉積通常會對魚類抗氧化能力產生負面影響[13,41]。在草魚[1]、大黃魚[17]、團頭魴[36]等硬骨魚中,磷脂可以提高抗氧化能力。本研究中,飼料中添加磷脂顯著降低了烏鱧幼魚肝臟中MDA 的含量,提高了總抗氧化能力及SOD 的活力,減輕了幼魚肝臟的氧化損傷,說明飼料中補充磷脂有助于增強烏鱧幼魚抗氧化能力,減少了脂質過氧化。
3.4 飼料磷脂對烏鱧幼魚肝臟、肌肉脂肪酸組成的影響
通常養殖魚類的脂肪酸組成通常與實驗飼料中的脂肪酸組成有關。在本研究中,肝臟和肌肉中C18:2n-6、C18:3n-3 和EPA(百分比脂肪酸)的濃度相對低于實驗飼料,而樣品中的DHA 濃度顯著高于相應實驗飼料中的DHA。淡水和鮭科魚類可有效地將C18 PUFA 轉化為長鏈PUFA[42]。本研究中,魚類樣品和實驗飼料中脂肪酸含量的分析結果比較說明烏鱧具有長鏈PUFA 生物合成能力,并可進行EPA 與DHA 的轉換,在之前的一些研究中也有類似的結果[14,39]。同時,本研究中飼料磷脂含量顯著提高了EPA 和DHA 等長鏈PUFA 的濃度,但降低了C18 PUFA 濃度,這表明高濃度磷脂促進了C18 PUFA 向長鏈PUFA 轉化。值得注意的是,已證實n-3 長鏈PUFA 在緩解草魚[43]和大西洋鮭[44]等魚類脂質沉積方面的潛在作用,本研究n-3 長鏈PUFA 濃度的增加部分說明了磷脂對脂質積累的緩解作用。
4 結論
在飼料中補充大豆磷脂可提高烏鱧幼魚的生長性能,減少體內脂肪沉積,增強魚體抗氧化能力,并通過調節脂肪酸代謝相關基因表達,抑制脂肪酸合成并促進脂肪氧化分解緩解肝臟脂肪積累。基于生長指標,本研究中烏鱧幼魚飼料適宜磷脂添加水平為5.36%。
猜你喜歡
中學生數理化·中考版(2022年10期)2022-11-10 09:37:42
中學生數理化·八年級物理人教版(2022年12期)2022-02-14 07:08:42
中學生數理化·八年級物理人教版(2021年12期)2021-12-31 03:23:08
中學生數理化·中考版(2020年10期)2020-11-27 01:59:48
中國生殖健康(2019年2期)2019-08-23 08:12:08
石油化工建設(2018年6期)2018-04-22 03:16:54
產品可靠性報告(2017年7期)2017-09-05 09:49:12
中學生數理化·八年級物理人教版(2017年12期)2017-04-18 12:59:38
汽車觀察(2016年3期)2016-02-28 13:16:26
民生周刊(2014年7期)2014-03-28 01:30:54
