CD4+T細胞極化在炎癥性疾病中作用的研究進展①
2023-12-28 10:23:14薛丹風江淑玲凌鑫萍南昌大學第一附屬醫院南昌330006
中國免疫學雜志 2023年12期
關鍵詞:小鼠
晏 偉 薛丹風 江淑玲 凌鑫萍 李 娜 (南昌大學第一附屬醫院,南昌 330006)
1 CD4+T細胞極化概述
CD4+T 細胞向特定T 細胞表型的特定細胞分化被稱為CD4+T 細胞極化,其在機體炎癥調節過程中發揮重要作用[1-2]。受外界環境及細胞因子等因素影響,CD4+T 細胞可極化為Th1、Th2、Th17 和Treg 等效應細胞。具體而言:①IL-12 上調T-bet,使CD4+T細胞極化為Th1細胞。Th1細胞會釋放IFN-γ、TNF-α及IL-2,從而加劇炎癥反應;②IL-4 促使STAT6 激活后,誘導GATA3表達,引發Th2極化。Th2細胞又可分泌IL-4及IL-13等發揮抑炎作用;③TGF-β、IL-1β、IL-6 等共同作用使RORγt 活化,進而實現Th17 極化。Th17 細胞會釋放IL-17 等促炎因子促進炎癥;④TGF-β 誘導Foxp3 表達進而促進Treg 極化。Treg細胞又可釋放IL-10、TGF-β 等抑炎因子來抑制炎癥。
因此,本綜述旨在總結并探討CD4+T 細胞極化在不同炎癥性疾病中的調控作用及機制,以期為后續展開相關研究提供參考和依據。
2 CD4+T細胞極化在炎癥性疾病中的作用
CD4+T 細胞極化與多發性硬化(multiple sclerosis,MS)、結腸炎、類風濕關節炎(rheumatoid arthritis,RA)及特應性皮炎(atopic dermatitis,AD)等多種炎癥性疾病有關,詳見表1,具體闡述如下。
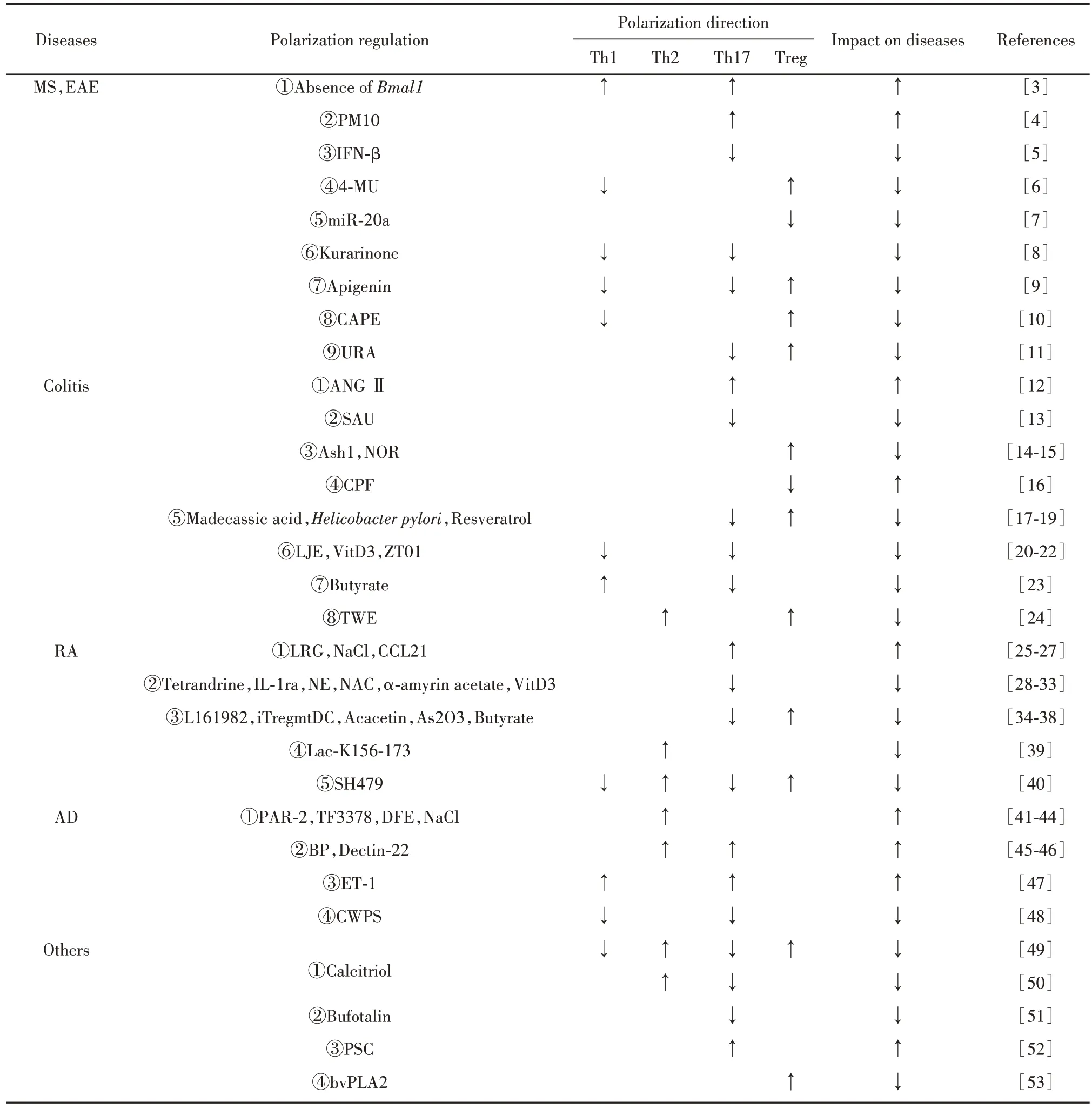
表1 CD4+T細胞極化在炎癥性疾病中的作用Tab.1 Role of CD4+T cell polarization in inflammatory diseases
2.1 CD4+T 細胞極化與MS MS 是一種以中樞神經系統損害為特征的自身免疫炎癥性疾病。實驗性自身免疫性腦脊髓炎(experimental autoimmune encephalomyelitis,EAE)是MS 的動物模型。Th1 和Th17 極化增強可加劇MS 及EAE 疾病進展。SUTTON 等[3]發現,小鼠髓系細胞時鐘基因Bmal1缺失能使分泌IL-1β 的CD11b+Ly6hi單核細胞擴增和浸潤,進而引起Th1 和Th17 反應增強,并產生更多致病性IFN-γ+/IL-17+T細胞,加重EAE。CORTESE等[4]研究顯示,空氣污染物顆粒物10(PM10)可誘導先天免疫細胞產生Th17極化細胞因子(IL-1β、IL-6)加劇MS炎癥。
常用藥干擾素-β(IFN-β)可通過活化的樹突狀細胞(dendritic cell,DC)限制CD4+T 細胞向Th17 極化,從而減輕EAE或MS的嚴重程度[5]。而4-甲基傘形酮(4-MU)可促使T 細胞從Th1 表型轉向FoxP3+Treg 表型,進而減輕EAE[6]。但近期有研究報道,miR-20a 能通過降低絲裂原活化蛋白激酶9(Map3k9)表達抑制Treg極化,減輕EAE[7]。
不僅如此,天然類物質也被廣泛應用于MS 及EAE 的治療。苦參素可抑制Th1 及Th17 極化延緩EAE 進展[8]。芹菜素可通過下調RelB 蛋白將Th1和Th17 表型轉變為Treg 發揮抗炎作用,其降低了Tbet、IFN-γ 及IL-17 表達,但增加了IL-10、TGF-β 和FoxP3 表達[9]。咖啡酸苯乙酯(CAPE)通過抑制NFκB 活化使CD4+IFN-γ+T 細胞比例下調、CD4+Foxp3+T細胞比例升高,進而在MS 中發揮抗炎功效[10]。果類成分鞣花酸可在腸道中形成烏蘿卜素A(URA),其可直接靶向芳烴受體(AhR),隨后抑制Th17 極化并阻礙IL-17、IFN-γ分泌,上調抑炎因子IL-10水平,最終減輕EAE[11]。
2.2 CD4+T 細胞極化與結腸炎 結腸炎疾病進展涉及CD4+T 細胞極化。血管緊張素Ⅱ(ANG Ⅱ)通過JAK2/STAT 信號通路,不僅直接促進Th17 極化,亦可誘導結腸上皮細胞釋放TGF-β1 及IL-6 間接促進Th17 極化,加劇結腸炎癥[12]。與之相反,茄子酮(SAU)不僅抑制Th17 分化,還可通過Blimp-1 途徑促使IL-10 產生,進而改善結腸炎[13]。H3K4 甲基轉移酶Ash1 通過增強TGF-β-Smad2/3 信號、去甲酚定(NOR)通過調節AhR/糖酵解軸及NAD+/SIRT1/SUV39H1/H3K9me3 信號通路,均可促進Treg 極化,預防并減輕結腸炎[14-15]。給予有機磷農藥毒死蜱(CPF)后,結腸炎小鼠可通過ACh 和AChRs 相互作用抑制Treg 極化,下調IL-10 和Foxp3 mRNA 表達,最終加重結腸組織損傷[16]。
調控Th17/Treg 動態平衡亦可改善結腸炎。XU等[17]發現,羥基乙酸通過PPARγ/AMPK/ACC1 信號通路降低結腸炎小鼠Th17 比例,提高Treg 比例,從而改善結腸炎。幽門螺桿菌定植可降低結腸炎小鼠IL-6 釋放、增加TGF-β 表達,隨后抑制Th17 反應并誘導Treg 反應,同時降低IL-17 水平,增加IL-10表達,最終在結腸炎中發揮保護作用[18]。ALRAFSA等[19]研究表明,白藜蘆醇誘導的微生物群對結腸炎治療有積極作用,表現出對CD4+Foxp3+T 細胞的極化促進及CD4+IFN-γ+T和CD4+IL-17+T細胞的減少。
此外,多種藥物還可通過抑制Th1/Th17 極化來防治結腸炎。一項探討金銀花水提物(LJE)灌胃給藥對小鼠結腸炎影響的實驗表明,LJE 可通過下調Th1及Th17相關炎癥細胞因子(IL-12、TNF-α、IFN-γ和IL-1β、IL-6、IL-17)預防結腸炎[20]。維生素D3(VitD3)、ZT01 均通過JAK-STAT 信號通路抑制Th1和Th17 極化,進而治療結腸炎[21-22]。與上述研究有出入的是,丁酸可通過差異化調節Th1 和Th17 分化控制T 細胞誘導結腸炎的能力:在Th1 極化條件下,丁酸促進T 細胞表達T-bet,而在Th17 極化條件下,其會抑制Th17 相關轉錄因子,而后促進Th1 發育但抑制Th17 發育[23]。塔希伯水提物(TWE)使巨噬細胞偏向M2表型后,可通過誘導Th2和Treg極化改善結腸炎[24]。
2.3 CD4+T 細胞極化與RA RA 是最常見的炎癥性關節炎,其動物模型為膠原誘導性關節炎(collagen induced arthritis,CIA)。富含亮氨酸的α2 糖蛋白(LRG)可通過增強TGF-β-Smad2 途徑促進Th17極化,加速CIA 進展[25]。JUNG 等[26]發現,RA患者滑液中Na+和IL-17 含量較高,且高鹽飼養的CIA 小鼠脾臟中Th17比例及腸道、滑膜中IL-17表達均較高,由此指出NaCl 可能通過影響Th17 細胞分化加重RA。VAN RAEMDONCK 等[27]研究顯示,趨化因子CCL21 在RA 滑膜組織中的大量表達可使IL-6 表達增加,進而驅動Th17極化,加重RA。
而抑制Th17 極化有益于RA 及CIA 的防治。粉防己堿可通過AhR途徑減少Th17細胞分化,達到改善小鼠CIA 目的[28]。LEE 等[29]發現人骨髓間充質干細胞(MSCs)表達的抗炎細胞因子白細胞介素1 受體拮抗劑(IL-1ra)可通過抑制IL-1RaKO 小鼠Th17極化改善其炎癥。同時,LIU 等[30]證明去甲腎上腺素(NE)可通過激活b2-AR/PKA 信號抑制Th17 極化,進而在CIA 中發揮抗炎作用。而N-乙酰半胱氨酸(NAC)、α-香菇蛋白醋酸酯通過抑制RORγt 轉錄、VitD3 通過miR-124 介導的IL-6 信號轉導,均可抑制Th17極化,進而改善RA及CIA[31-33]。
調控Th17 與Treg 的極化狀態亦可改善RA 及CIA。L161982 干預后,提高了CIA 小鼠Treg 水平、降低了小鼠血漿和組織中IL-17 水平、減少了Th17極化,最終減輕了CIA 嚴重程度[34]。成熟的耐受樹突狀細胞(iTre-gmtDC)可表達更高水平的FoxP3,在更大程度上抑制Th17 分化,并促使Treg/Th17 反應平衡,進而更有效地抑制CIA[35]。植物提取物金合歡素可通過增強Treg 反應、抑制Th17 反應修復Th17/Treg 失衡,從而降低CIA 的發生率[36]。同樣,三氧化二砷(As2O3)通過STAT3調節,不僅顯著抑制RA 患者Th17 分化、促進Treg 分化,也降低了IL-17、IL-6 和TNF-α 表達、增強了IL-10 和TGF-β 釋放,進而改善RA[37]。而丁酸可通過顯著抑制IL-1β、IL-6、IL-17 釋放、上調IL-10 表達及促進Treg 極化抑制CIA 炎癥[38]。多肽Lac-K156-173 可通過甘露糖受體(MR)調節DC 的分化、成熟和功能,進而上調Jagged1,隨 后 引 起Th2 極 化,顯 著 改 善 小 鼠CIA[39]。CHEN 等[40]研究指出,SH479 可通過JAK-STAT 信號通路將Th17/Th1 反應轉變為Th2/Treg 表型來治療RA,其可降低IL-17 和IFN-γ 水平并增強IL-10 和IL-4表達。
2.4 CD4+T 細胞極化與AD AD 是一種常見的以Th2 反應為主的慢性炎癥性皮膚病。蛋白酶激活受體-2(PAR-2)的激活會引起胸腺基質淋巴生成素(TLSP)產生及嗜堿性粒細胞聚集,進而促進Th2 極化,加劇AD[41]。而AD 患者皮膚中的金黃色葡萄球菌TF3378 亦可誘導Th2 免疫反應,增加AD 治療難度[42]。粉 塵 螨 提 取 物(DFE)首 先 激 活TLR1 和TLR6,隨后刺激角質形成細胞釋放IL-25 和IL-33,進一步引起Th2免疫應答極化,最終加劇AD的過敏癥狀及炎癥反應[43]。研究證明,AD患者皮膚中NaCl比例較高,其可通過滲透敏感轉錄因子NFAT5 和激酶SGK-1 誘導Th2 極化,塑造適宜AD 進展的微環境,加重AD[44]。
此外,HONG 等[45]發現煙霧中的苯并芘(BP)可通過Ahr/Arnt 軸極化Th2 及Th17 反應,在致敏階段誘導AD。LIANG 等[46]研究指出,Dectin-22 介導的過敏原致敏及增敏可使IL-33、IL-6等Th2和Th17相關炎癥因子上調,隨后引起TSLP 誘導Th2/Th17 極化,進而引發AD。NAKAHARA 等[47]證實AD 患者表皮中內皮素-1(ET-1)上調,ET-1可使DC-T細胞反應向Th17/Th1 極化轉變,隨后加重AD。值得慶幸的是,天然草藥提取物白蠟蟲粗多糖(CWPS)可通過抑制Th1/Th17 極化減輕AD 炎癥反應,在CWPS干預組中,Th1 及Th17 的比例、T-bet 和RoRγt 的基因表達、細胞因子(IFN-γ、IL-17A 和IL-6)的表達均明顯低于生理鹽水組[48]。
2.5 CD4+T 細胞極化與其他炎癥性疾病 除上述疾病外,CD4+T 細胞極化也與牙周炎等其他炎癥性疾病相關。BI 等[49]研究顯示,骨化三醇干預的大鼠牙周炎組織IL-17水平降低、IL-4和IL-10水平升高,外周血中Th2 和Treg 百分比增加、Th1 和Th17 百分比下降。該團隊還發現骨化三醇處理后,小鼠Th細胞表現出Th2 極化增強,GATA3 和STAT5 的基因及蛋白水平均提高;而Th17 極化降低,RoRγt 基因及蛋白水平也降低[50]。
蟾蜍林能抑制Th17 極化及IL-17、IFN-γ 分泌,進而改善慢性炎癥性自身免疫性疾病干燥綜合征(SS)炎癥反應[51]。另有研究指出,CD4+T 細胞在原發性硬化性膽管炎(PSC)中呈現出Th17 極化,傾向[52]。而蜂毒磷脂酶A2(bvPLA2)可誘導Foxp3+Treg極化減輕小鼠多發性關節炎[53]。
3 總結與展望
綜上所述,CD4+T 細胞極化在MS、結腸炎、RA及AD 等多種炎癥性疾病的發生發展及防治中發揮重要作用。總體而言,CD4+T 細胞向Th1 及Th17 極化會加重炎癥性疾病,而向Th2 及Treg 極化有利于減輕炎癥。但這并不絕對,如miR-20a 可通過抑制Treg 極化減輕EAE[7];丁酸可通過差異化調節Th1和Th17 分化控制T 細胞誘導結腸炎的能力[23]。而在AD 疾病中,因其還涉及一定的致敏過程等,發病機制更為復雜,Th2極化增強會加重疾病。
CD4+T 細胞的可塑性極強,其向某一亞型效應細胞群的極化是動態變化的。因此,上述多項研究通過外在的藥物干預人為調控CD4+T細胞的極化狀態,使其向有益于減輕和改善炎癥性疾病的方向發展。同時,CD4+T 細胞的極化也是一個極其復雜的免疫過程。不同藥物可通過促進或抑制同一極化方向達到相同的防治效果。而同一藥物在不同疾病中,其極化作用也可能不同[23,38]。提示在今后進行相關研究時,應具體問題具體分析。另外,CD4+T細胞極化涉及的途徑或靶基因尚未闡明,期待今后的研究能豐富相關內容。
最后,天然類物質在調控CD4+T 細胞極化以改善炎癥性疾病方面具有廣闊的前景,下一步研究可以此為切入點探討炎癥性疾病防治的新策略。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
東方少年·布老虎畫刊(2023年8期)2023-08-01 15:45:12
科學大眾(2021年6期)2021-07-20 07:42:44
科學(2020年3期)2020-11-26 08:18:30
學苑創造·A版(2020年9期)2020-10-13 09:41:02
娃娃樂園·綜合智能(2019年3期)2019-04-03 09:17:36
中成藥(2018年2期)2018-05-09 07:19:34
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
湖北師范大學學報(自然科學版)(2015年2期)2016-01-10 08:41:55
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
