玉曲河流域裂腹魚類游泳能力及其在魚道設計中的應用
2024-01-01 00:00:00劉瀚譚均軍王永猛李陽希王猛柯森繁石小濤譚紅林王淵洋李衛東蒲進金志軍
水生態學雜志 2024年4期

DOI:10.15928/j.1674-3075.202208150326
收稿日期:2022-08-15""""" 修回日期:2023-07-19
基金項目:國家自然科學基金(52179070);國家優秀青年科學基金(51922065);貴州省水利科技基金(KT202120);西藏玉曲河扎拉水電站過魚設施專題研究。
作者簡介:劉瀚文,1997 年生,男,碩士研究生,研究方向為生態水力學。E-mail:1840407588@qq.com
通信作者:譚均軍,1985年生,女,博士,副教授,主要從事水生態及環境研究。E-mail:tanjunjun52@163.com
摘要:探究玉曲河流域裂腹魚類的游泳能力,為過魚設施建設和魚類游泳行為學提供基礎參數。以該流域優勢種怒江裂腹魚(Schizothorax nukiangensis)、貢山裂腹魚(S. gongshanensis)為研究對象,在野外現場通過游泳能力測試水槽對裂腹魚的感應流速、臨界游泳速度、突進游泳速度及持續游泳速度、耐久游泳速度進行測定。結果表明,怒江裂腹魚和貢山裂腹魚的絕對感應流速相近,分別為(0.18±0.01) m/s和(0.18±0.06) m/s,絕對臨界游泳速度分別為(1.05±0.12) m/s和(1.11±0.14) m/s,絕對突進游泳速度分別為(1.45±0.26) m/s和(1.40±0.21) m/s,相對臨界游泳速度分別為(8.47±0.95) BL/s和(9.21±2.45) BL/s;二者的相對突進游泳速度均突破了10倍體長(BL),分別為(11.90±2.31) BL/s和(10.80±3.16) BL/s;怒江裂腹魚和貢山裂腹魚最大持續游泳速度均為0.63 m/s,最大耐久游泳速度分別為0.93 m/s和0.73 m/s。當以此2種裂腹魚為主要過魚對象時,建議魚道入口及豎縫處流速為1.05~1.40 m/s,過魚設施內部整體平均流速設計為0.24~1.05 m/s,休息池主流流速為0.18~1.05 m/s。研究結果可為西藏玉曲河流域魚類資源保護及過魚設施設計提供參考。
關鍵詞:怒江裂腹魚;貢山裂腹魚;感應流速;臨界游泳速度;突進游泳速度;魚道設計;玉曲河
中圖分類號:S956""""""" 文獻標志碼:A""""""" 文章編號:1674-3075(2024)04-0125-09
玉曲河為怒江中游左岸一級支流,干流總長444.3 km,流域面積9 379 km2,分布有2目3科7屬15種魚類(王龍濤,2015),天然落差達3 122 m,蘊藏有豐富的水能資源,扎拉水電站為玉曲河干流的第5梯級電站,計劃總裝機容量1 015 MW。水電站建設在為當地帶來經濟效益和社會效益的同時,大壩也阻斷了河流的縱向連通性(馮克義,2019),尤其對洄游性魚類產生了重大影響(Tan et al,2022)。
過魚設施作為一種有效協助魚類恢復其洄游通道的生態補償措施被廣泛應用,其設計依據則是流域過魚對象的游泳能力參數(侯軼群等,2020;Katopodis,2005)。為使魚類順利進入過魚設施,魚道進口誘魚水流流速不應過小,應大于臨界游泳速度且小于突進游泳速度(丁少波等,2020);魚類在到達過魚設施的內部后,為使其不失去趨流性,過魚設施整體的平均流速應大于魚類的感應流速且小于臨界游泳速度(周小波等,2017);在過魚設施休息區中,其設計流速應介于感應流速與臨界游泳速度(李志敏等,2018)。目前,與過魚設施設計相關的魚類游泳能力研究多為單一魚種,而過魚對象往往為多種,針對多種對象的過魚設施設計參數選取的研究較少。
本文以扎拉水電站上下游的魚類洄游習性、棲息地適應性、魚類保護等級、經濟價值及歷史地位等為依據,并兼顧其他魚類,確定西藏玉曲河流域的主要過魚對象為鯉科(Cyprinidae)、裂腹魚亞科(Schizothoracinae)、裂腹魚屬(Schizothorax)的怒江裂腹魚(S. nukiangensis)和貢山裂腹魚(S. gongshanensis);怒江裂腹魚主要分布在怒江干流,貢山裂腹魚主要分布于怒江上游及支流,為怒江水系特有種。有研究顯示,裂腹魚亞科是鯉科各亞科中受威脅最嚴重的類群,主要分布于青藏高原及周邊地區,是該區域內重要的經濟魚類,其棲息地受人類活動的影響較大(曹亮等,2016);此外,惡劣的高原環境決定了裂腹魚具有生長緩慢(陳鋒和陳毅鋒,2010)、繁殖能力低(劉潔雅,2016)等特點,種質資源的脆弱性導致其資源量一旦遭受破壞,將很難恢復(李雷等,2019),隨著對青藏高原開發程度的加大,裂腹魚類保護的形勢將更加嚴峻。本研究在野外測試其感應流速、臨界游泳速度、突進游泳速度、持續及耐久游泳速度,旨在為怒江水系魚類資源保護及電站過魚設施建設提供技術支撐。
1"" 材料與方法
1.1"" 過魚對象
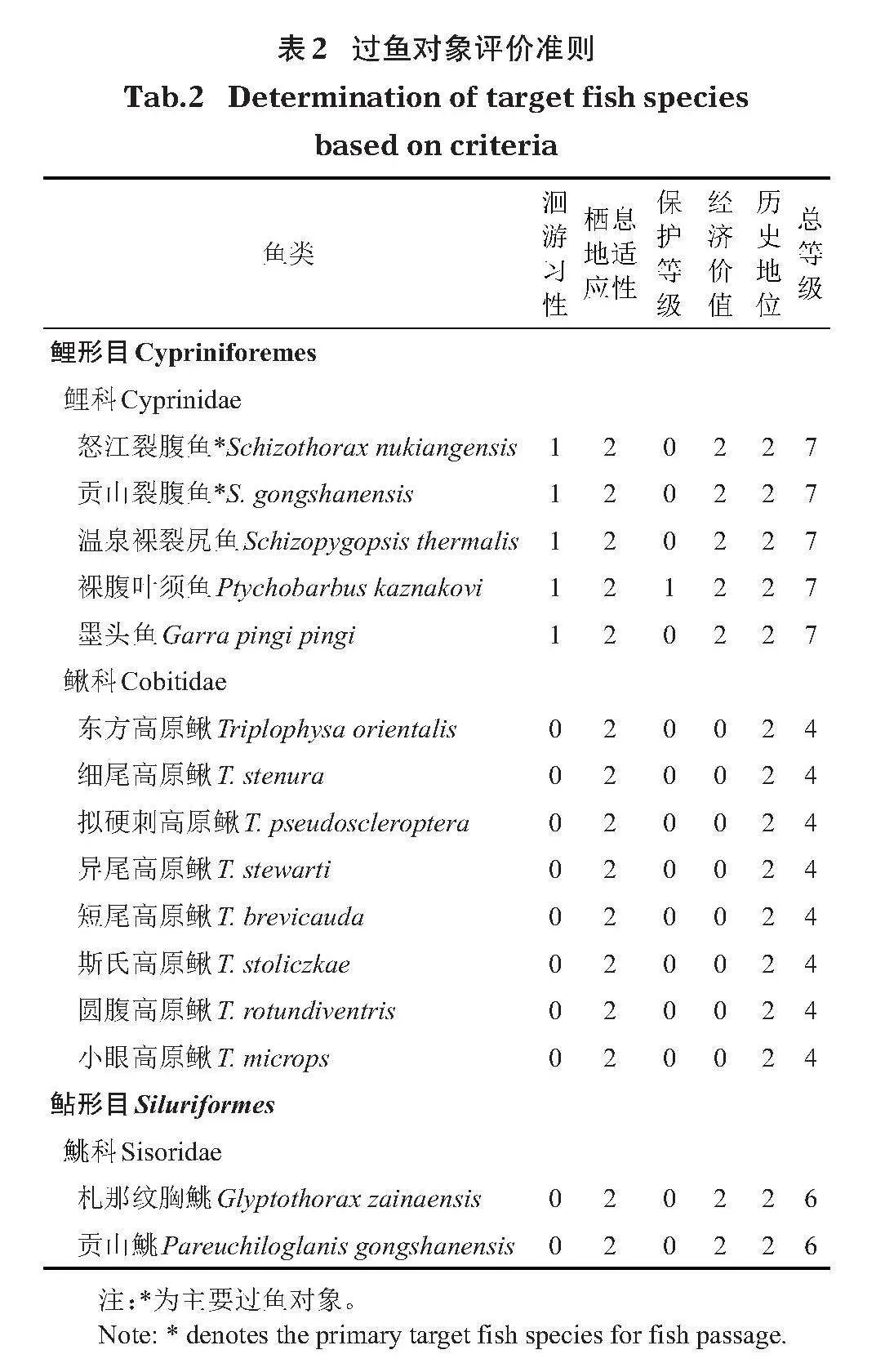
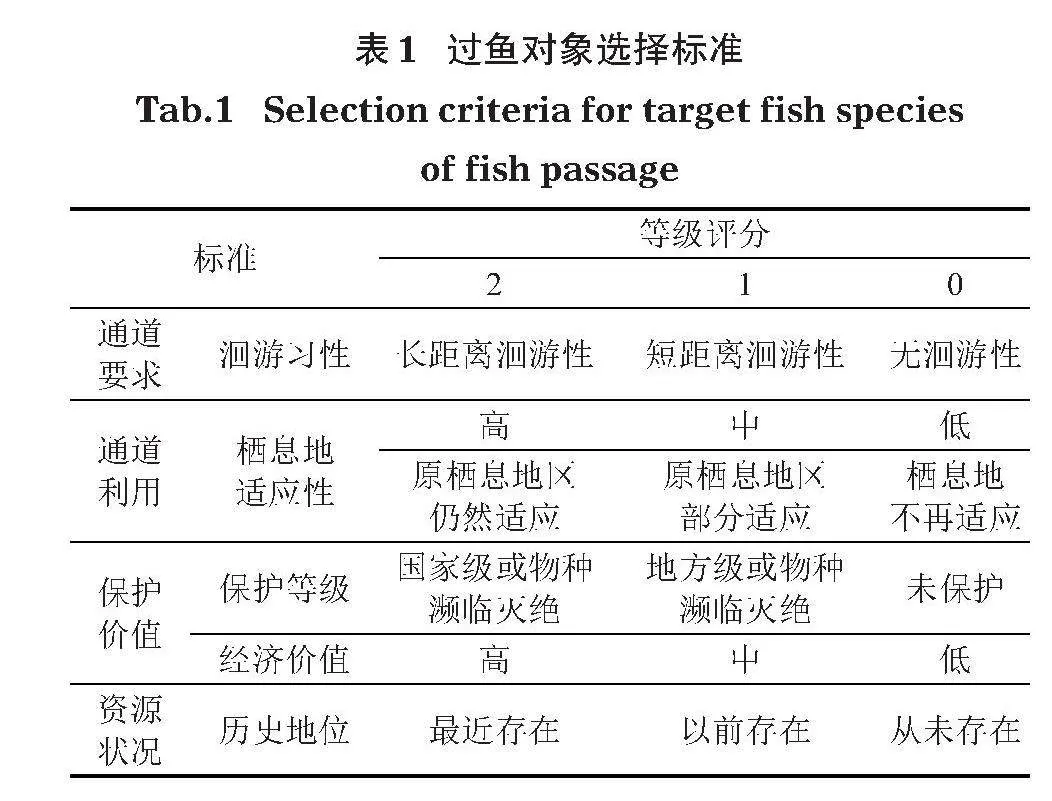
通過查閱《西藏玉曲河扎拉水電站環境影響報告書》(大唐玉曲河水電開發有限公司,2018)及《西藏魚類及其資源》(西藏自治區水產局,1995)等文獻資料,統計出西藏扎拉水電站上下游水域有魚類15種。根據蔡露等(2018)針對過魚對象劃分的5個標準,每個標準采用0、1、2進行分級(表1);主要過魚對象評價準則見表2。
結合環評要求及過魚對象評價,總等級為7的可作為主要過魚對象,但由于墨頭魚在該流域較為稀少,此次未捕獲,裸腹葉須魚和溫泉裸裂尻魚捕獲數量較少,故以怒江裂腹魚和貢山裂腹魚為主要過魚對象,其他裂腹魚類以及鰍科、鮡科魚類則作為兼顧過魚對象。
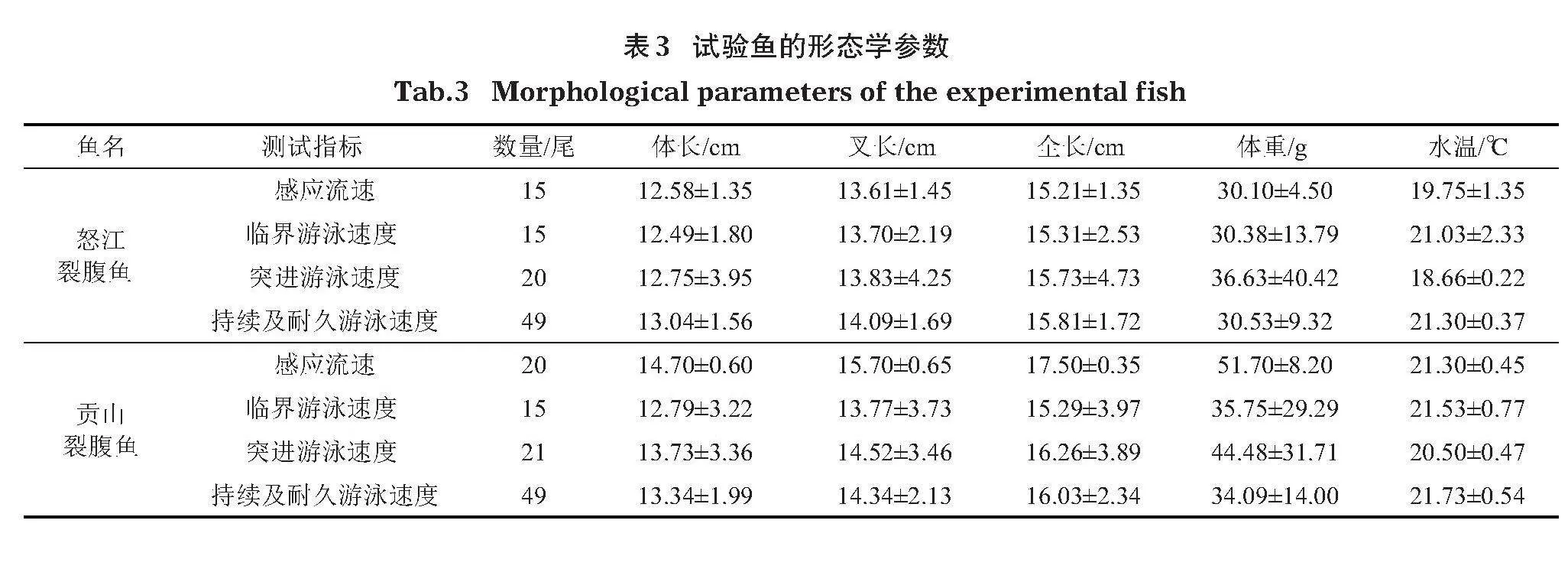
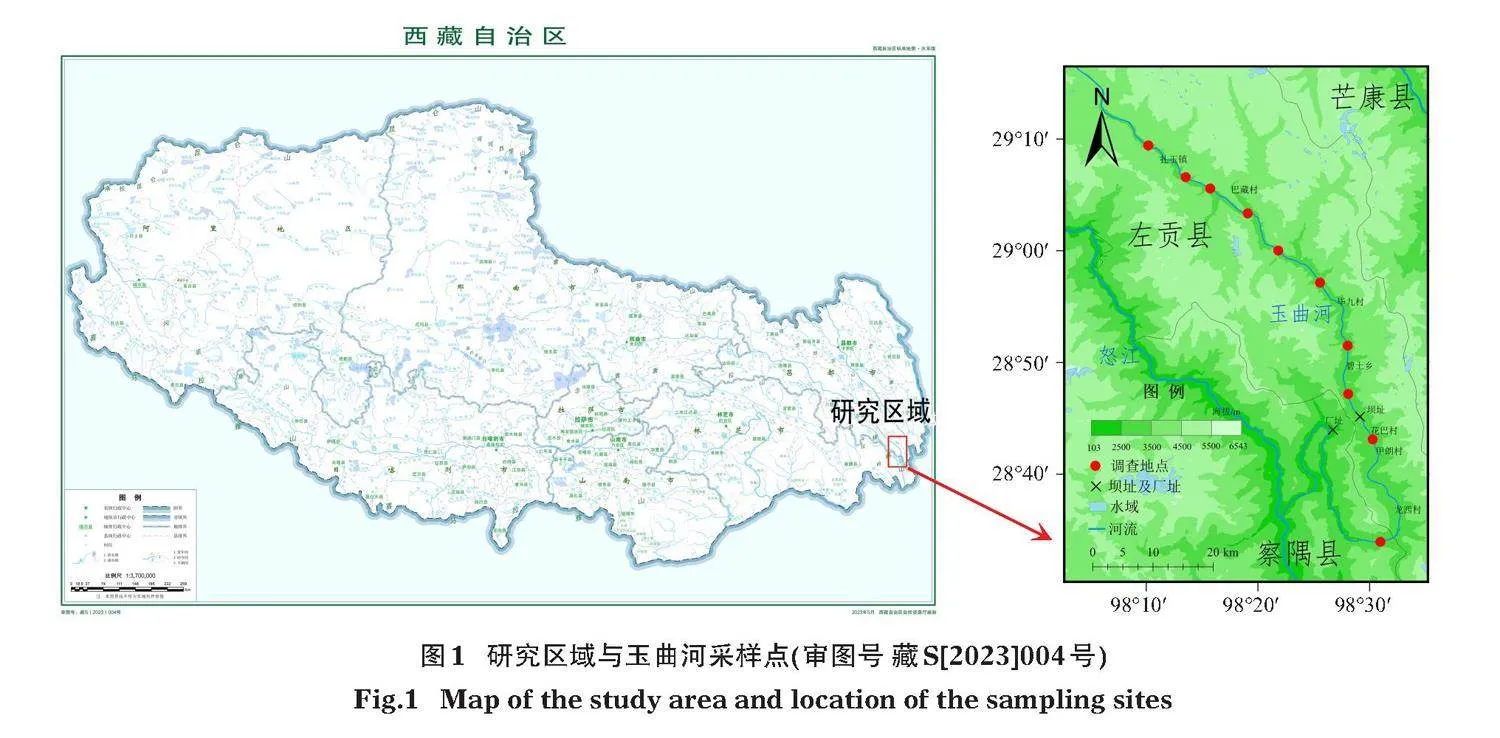
本次采樣地點位于我國西藏自治區芒康縣、左貢縣和察隅縣的玉曲河流域(圖1,國家自然資源部標準地圖服務網站HTTP://BZDT.CH.MNR.GOV.CN/,審圖號為藏S[2023]004,底圖無修改)。2021年6月中旬主要采用地籠捕獲怒江裂腹魚和貢山裂腹魚樣本,地點位于擬建大壩上下游80 km的區域。共捕獲怒江裂腹魚806尾,貢山裂腹魚282尾,從中挑選出健康個體作為測試對象進行試驗,其形態學參數見表3。
獲取的樣本怒江裂腹魚體長為7.9~22.5 cm,體重為5.3~145.0 g;貢山裂腹魚體長為8.1~21.4 cm,體重為9.9~135.6 g。將捕獲的試驗魚暫養于便攜式水槽中,每天換水1次,換水量約為總水量的30%,暫養期間持續充氧,使溶氧量維持在6 mg/L以上,試驗水槽與暫養池水溫維持在13.5~18℃。試驗結束后的樣本魚原地放生。
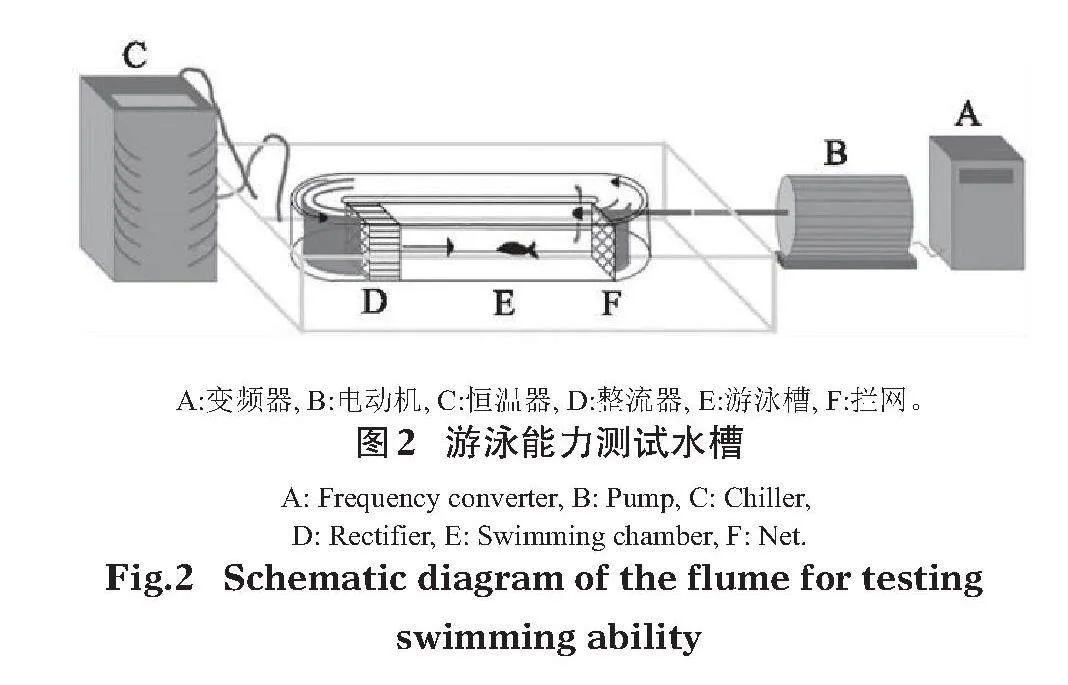
1.2"" 試驗裝置
試驗采用丹麥Loligo System公司生產的游泳水槽SW10150、SW10050進行魚類游泳能力測試(圖2)。測試區長×寬×高為55 cm×14 cm×14 cm、30 cm×7.5 cm×7.5 cm,測試水槽容積為30 L和5 L。根據試驗魚體長的不同,采用不同型號的試驗裝置,以保證數據的可靠性(王永猛等,2021)。電動機轉速在變頻器的控制下,產生不同的水流速度,位于測試區上游的蜂窩狀穩流裝置可維持測試區內的流場穩定。
1.3"" 游泳能力測試
1.3.1"" 感應流速"" 通常采用“遞增流速法”測定。將單尾魚置于測試水槽中,為減小試驗魚初入水槽的應激反應(徐革鋒等,2015),在1 BL/s的流速下適應1 h后,每隔5 s微調變頻器逐步增大流速,同時觀察試驗魚的游泳行為。當試驗魚隨著流速的緩慢增加出現游泳姿態擺正至頭部朝向來水方向且均勻擺尾時,此時的流速即為試驗魚的感應流速。感應流速分為絕對感應流速(Uind,m/s)和相對感應流速(U'ind,BL/s)。
1.3.2"" 臨界游泳速度"" 測試采用“遞增流速法”。先將單尾試驗魚置于流速為1 BL/s的水槽中以緩解魚的應激反應;適應1 h后,每隔15 min增加1 BL/s的流速,直至試驗魚疲勞,表現為試驗魚因不能抵抗水流速度維持游動,停靠在下游鐵網上,輕拍水槽下游壁面20 s仍不重新游動,隨即撈取試驗魚并測量其體重、體長、體寬等形態學參數。絕對臨界游泳速度(Ucrit,cm/s)按照下式計算。
Ucrit = Ut +[tΔt]ΔU ①
式中:Ut為魚能夠完成持續時間Δt的最大游泳速度,t為在最高流速下魚的實際游泳持續時間(tlt;Δt),Δt為改變流速的時間間隔(即15 min),ΔU為流速的增量(1.0 BL/s)。
相對臨界游泳速度(U'crit,BL/s)由絕對臨界游泳速度(Ucrit,cm/s)除以試驗魚體長求得:
U'crit =" [UcritBL]"""""""""""""""""""""""""" ②
式中:BL為試驗魚的體長(cm),試驗魚最大橫截面積小于水槽游泳區橫截面積的10%,不會產生阻塞效應,故不需對結果進行校正(Smit et al,1971)。
1.3.3"" 突進游泳速度"" 測試也采用“遞增流速法”,類似于臨界游泳速度的測試方法;不同之處在于將流速提升時間間隔Δt改變為20 s,流速增量依舊選擇1.0 BL/s,魚體疲勞時的速度即為突進游泳速度(于曉明等,2009)。突進游泳速度計算公式與臨界游泳速度計算公式一致。
通過爆發游泳速度預測魚道最大流速區域的間隔距離:
D = (Uburst - Vf) × 20nbsp;"""""""""""""""""""""""""""""""""""""" ③
式中:D為魚類游泳距離(cm),Uburst為持續游泳20 s能夠達到的最大游泳速度(cm/s),Vf為水流流速(cm/s)。
1.3.4"" 持續及耐久游泳速度"" 持續游泳速度和耐久游泳速度測試均采用“固定流速法”。試驗魚先在1.0 BL/s的流速下適應1 h,然后在1 min內將流速調至設定流速(Hammer, 1995)。設定流速的初值采用試驗魚種的平均臨界游泳速度,并記錄在該流速下的游泳時間;若持續游泳超過200 min,則停止試驗,每個流速下重復20尾。根據試驗結果,在該流速的基礎上調整下一組試驗魚的設定流速,改變值通常為0.1~0.2 m/s。若在某一流速下有50%試驗魚持續游泳時間t[≥]200 min,此時流速即為最大持續游泳速度。小于最大持續游泳速度的流速都稱為持續游泳速度(白艷勤等,2013)。
耐久游泳速度與持續游泳速度的測試步驟基本一致,其區別在于若在某一流速下有50%的魚持續游泳時間t≤20 s,則該流速為最大耐久游泳速度。最大可持續游泳速度至最大耐久游泳速度的范圍均為耐久游泳速度。根據可持續游泳速度,可計算出魚道內允許最大平均水流速度(Peake et al,1997):
Vf,max = max(Vs - d/EVs)""""""""""""""""""""""""""""""""" ④
式中:Vf,max為魚道內允許的最大水流速度(m/s),Vs為目標魚的游泳速度(m/s),d為魚道長度(m),EVs為目標魚在Vs下的游泳耐力(s)。
1.4"" 數據處理
試驗數據通過Excel 2019處理和統計,運用SPSS 19.0統計軟件,對試驗魚的感應流速、臨界游泳速度和突進游泳速度與體長之間的關系進行Pearson相關性檢驗,Origin 2018繪圖。試驗魚的趨流率與水流速度的關系通過非線性回歸進行擬合。統計顯著水平為Plt;0.05,結果均采用(平均值±標準差)表示。
2"" 結果與分析
2.1"" 感應流速
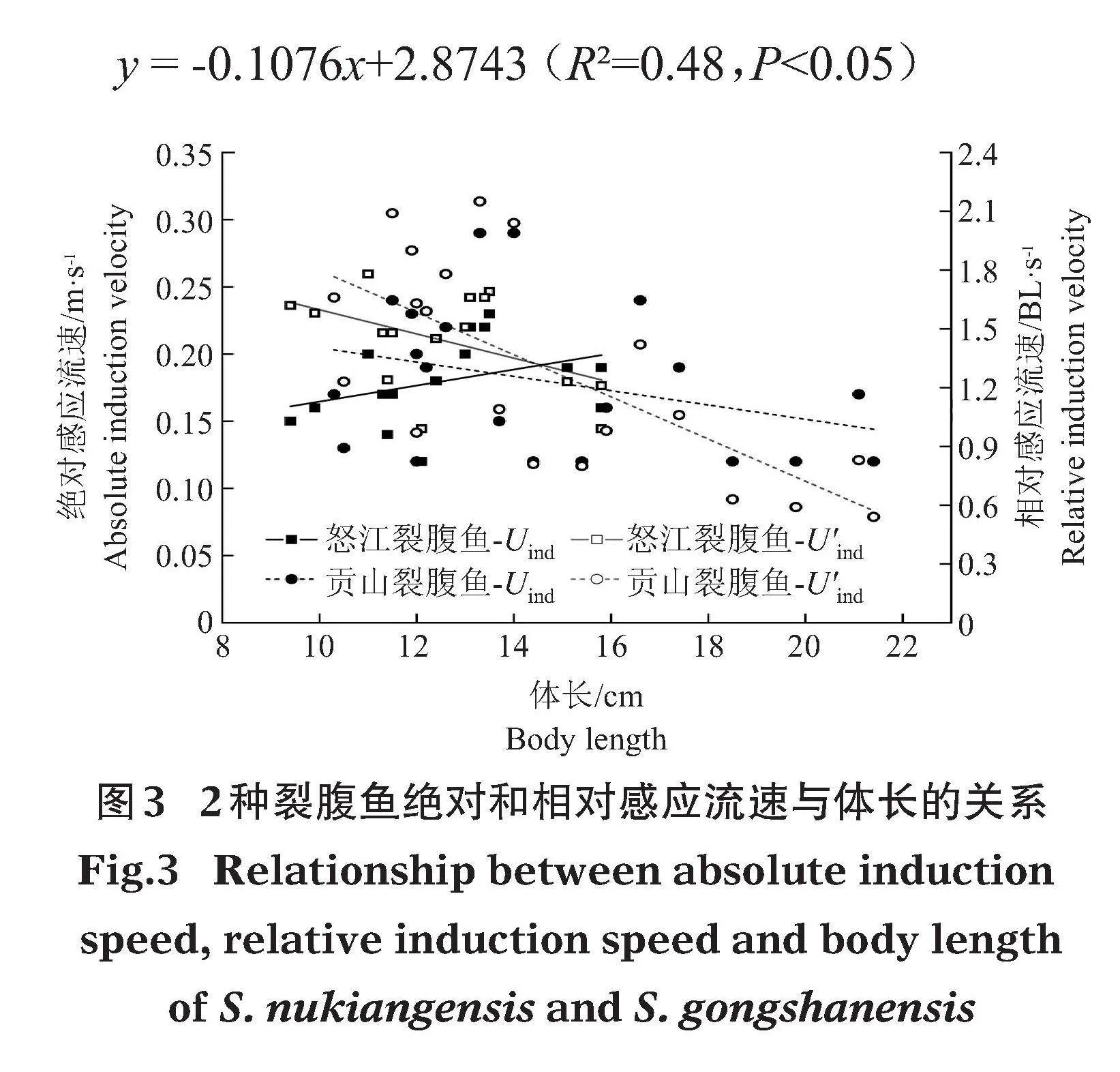
試驗測得怒江裂腹魚的絕對感應流速(Uind)為(0.18±0.01)m/s,相對感應流速(U'ind)為(1.44±0.24)BL/s;貢山裂腹魚Uind為(0.18±0.06)m/s,U'ind為(1.29±0.31)BL/s。對比發現,2種裂腹魚的Uind隨體長(BL)變化不明顯(Pgt;0.05),怒江裂腹魚的相對感應流速與體長無統計學意義。貢山裂腹魚相對感應流速隨體長增加有明顯降低的趨勢(Plt;0.05)(圖3),相對感應流速與體長的擬合關系式為:
y = -0.1076x+2.8743 (R2=0.48,Plt;0.05)
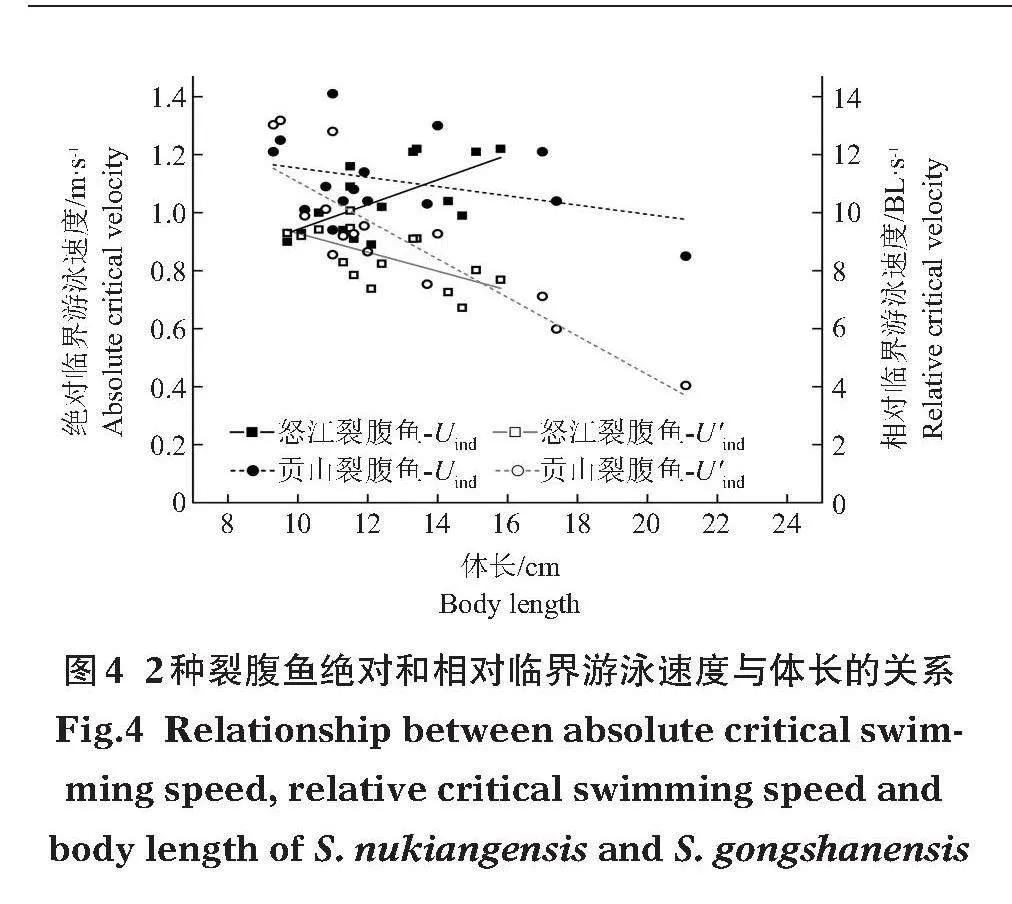
2.2"" 臨界游泳速度
怒江裂腹魚絕對臨界游泳速度(Ucrit)為(1.05±0.12)m/s,相對臨界游泳速度(U'crit)為(8.47±0.95)BL/s;貢山裂腹魚Ucrit為(1.11±0.14)m/s,U'crit為(9.21±2.45)BL/s。怒江裂腹魚絕對臨界游泳速度與體長關系不顯著(Pgt;0.05),但相對臨界游泳速度隨體長的增加而降低;貢山裂腹魚絕對和相對臨界游泳速度均與體長關系顯著(Plt;0.05)(圖4)。相對臨界游泳速度與體長的擬合關系式分別為:
怒江裂腹魚:
y = -0.3266x+12.5546 (R2 = 0.39,Plt;0.05)
貢山裂腹魚:
y = -0.6640x+17.7015 (R2 = 0.76,Plt;0.05)
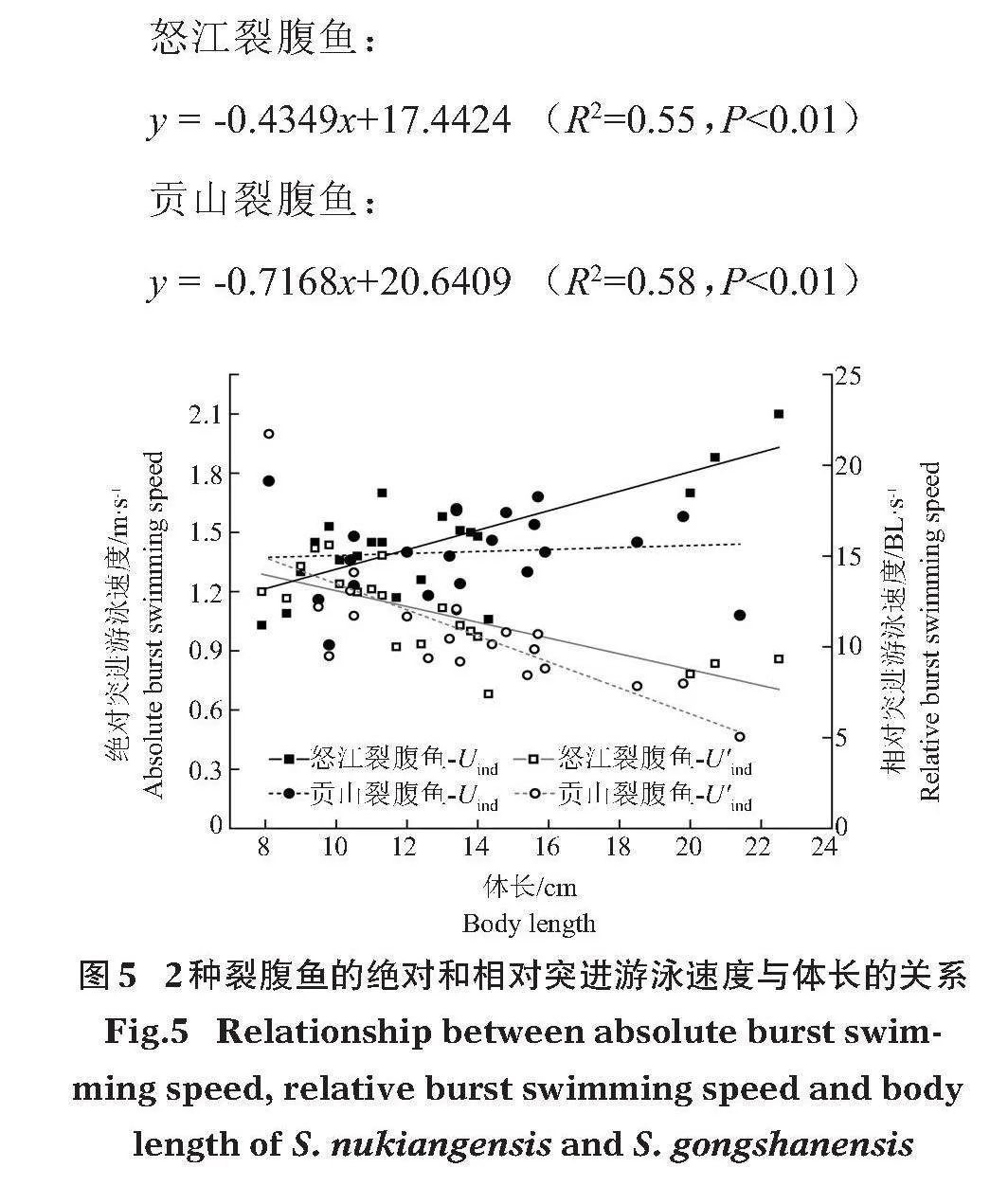
2.3"" 突進游泳速度
怒江裂腹魚絕對突進游泳速度(Ubrust)為(1.45±0.26)m/s,相對突進游泳速度(U'brust)為(11.90±2.31)BL/s;貢山裂腹魚Ubrust為(1.40±0.21)m/s,U'brust為(10.80±3.16)BL/s。怒江裂腹魚絕對和相對突進游泳速度與體長之間均存在極顯著關系(Plt;0.01),貢山裂腹魚的相對突進游泳速度與體長關系極顯著(Plt;0.01),相對突進游泳速度隨體長的增大而減小(圖5)。相對突進游泳速度與體長的擬合關系分別為:
怒江裂腹魚:
y = -0.4349x+17.4424"" (R2=0.55,Plt;0.01)""""
貢山裂腹魚:
y = -0.7168x+20.6409"" (R2=0.58,Plt;0.01)
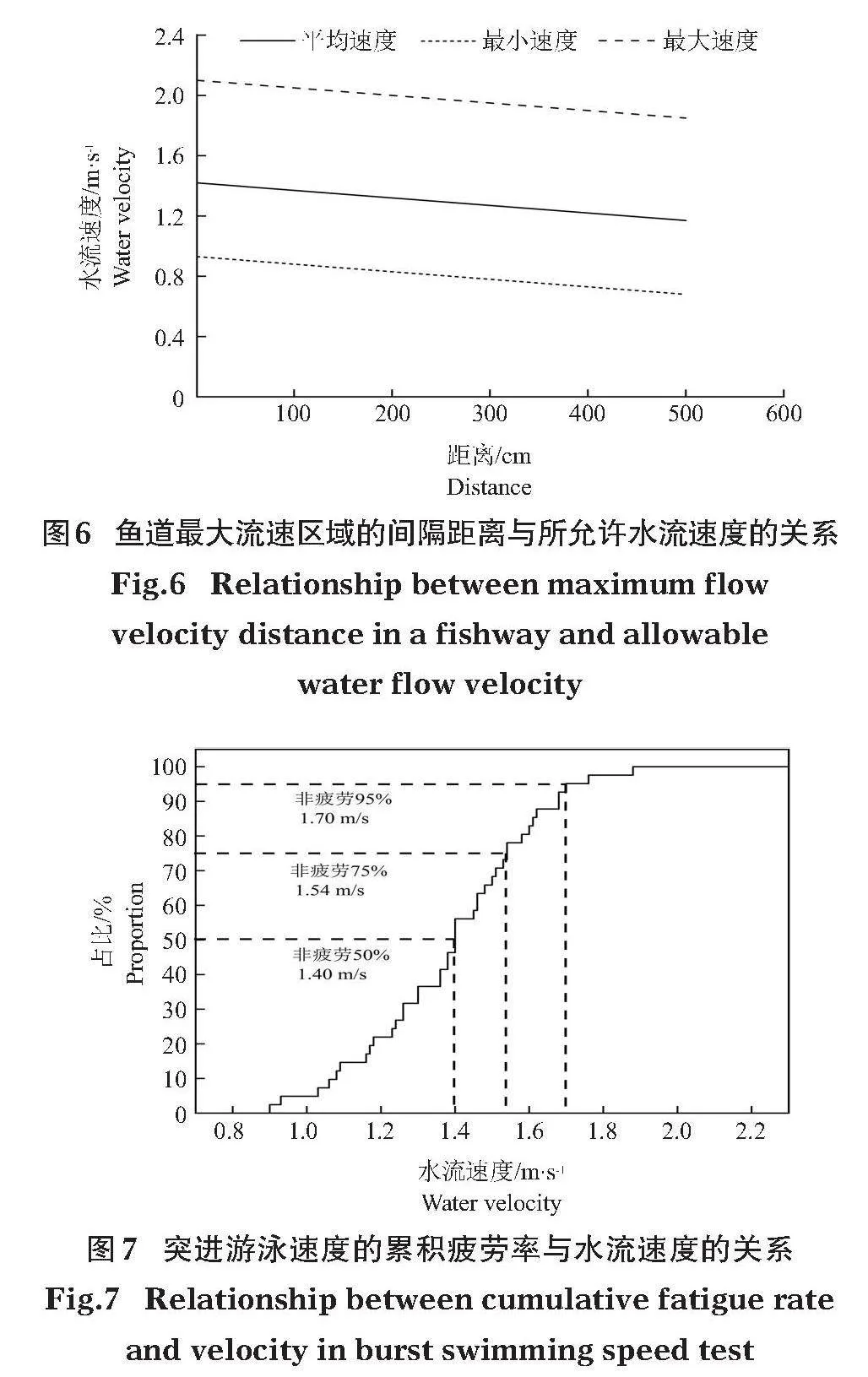
綜合考慮本研究的2種裂腹魚,最小突進游泳速度為0.93 m/s,平均突進游泳速度為1.42 m/s,最大突進游泳速度為2.10 m/s。據此可得最大流速區域的間隔距離與所允許的水流速度關系 (圖6)。
通過“生存分析”方法得出2種試驗魚突進游泳速度的累積疲勞率與水流速度的關系(圖7)。結果表明,在突進游泳速度的累積疲勞曲線中,95%、75%、50%的魚非疲勞突進游泳速度分別為1.70 m/s、1.54 m/s、1.40 m/s。
2.4"" 持續及耐久游泳速度
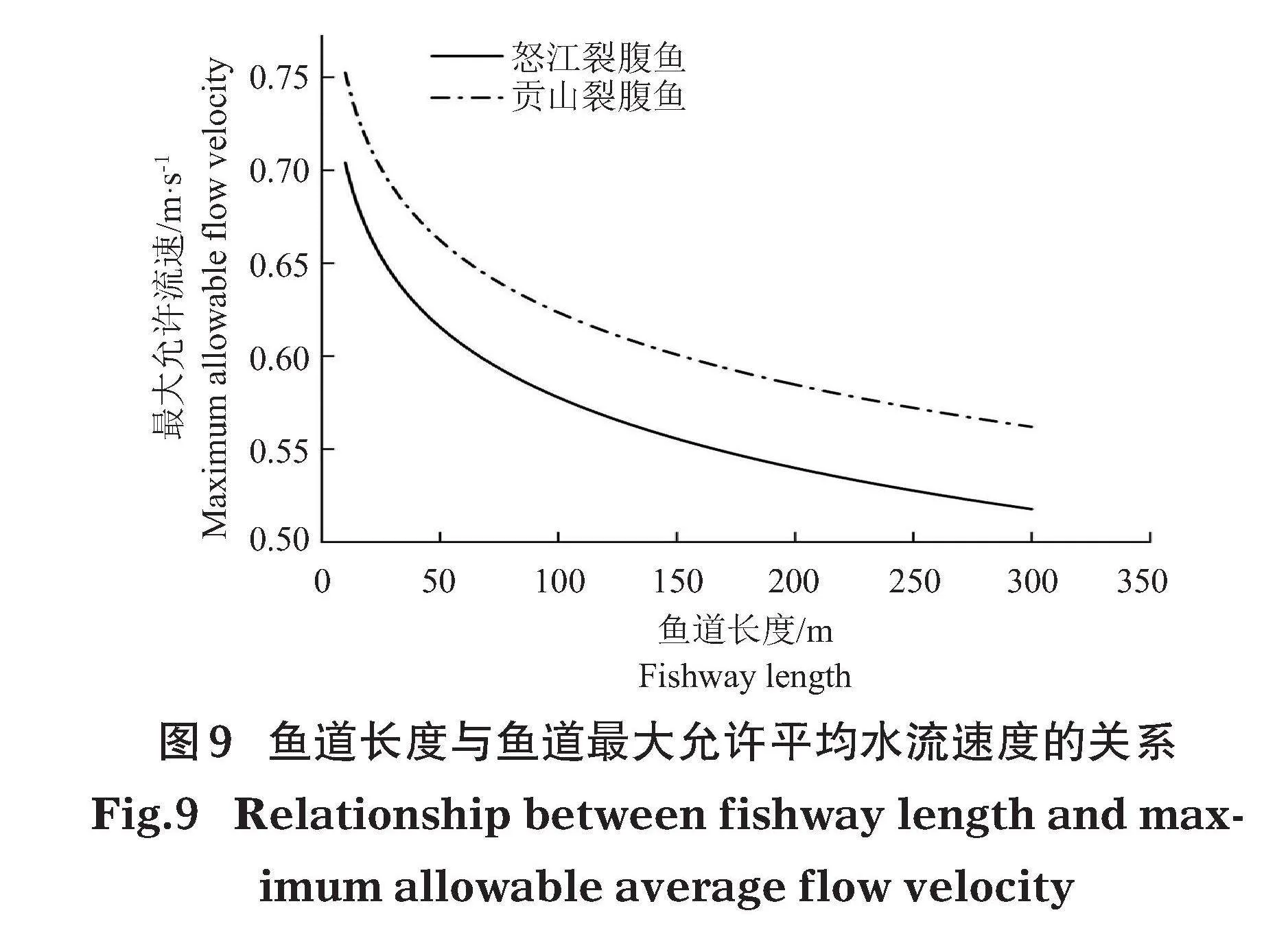
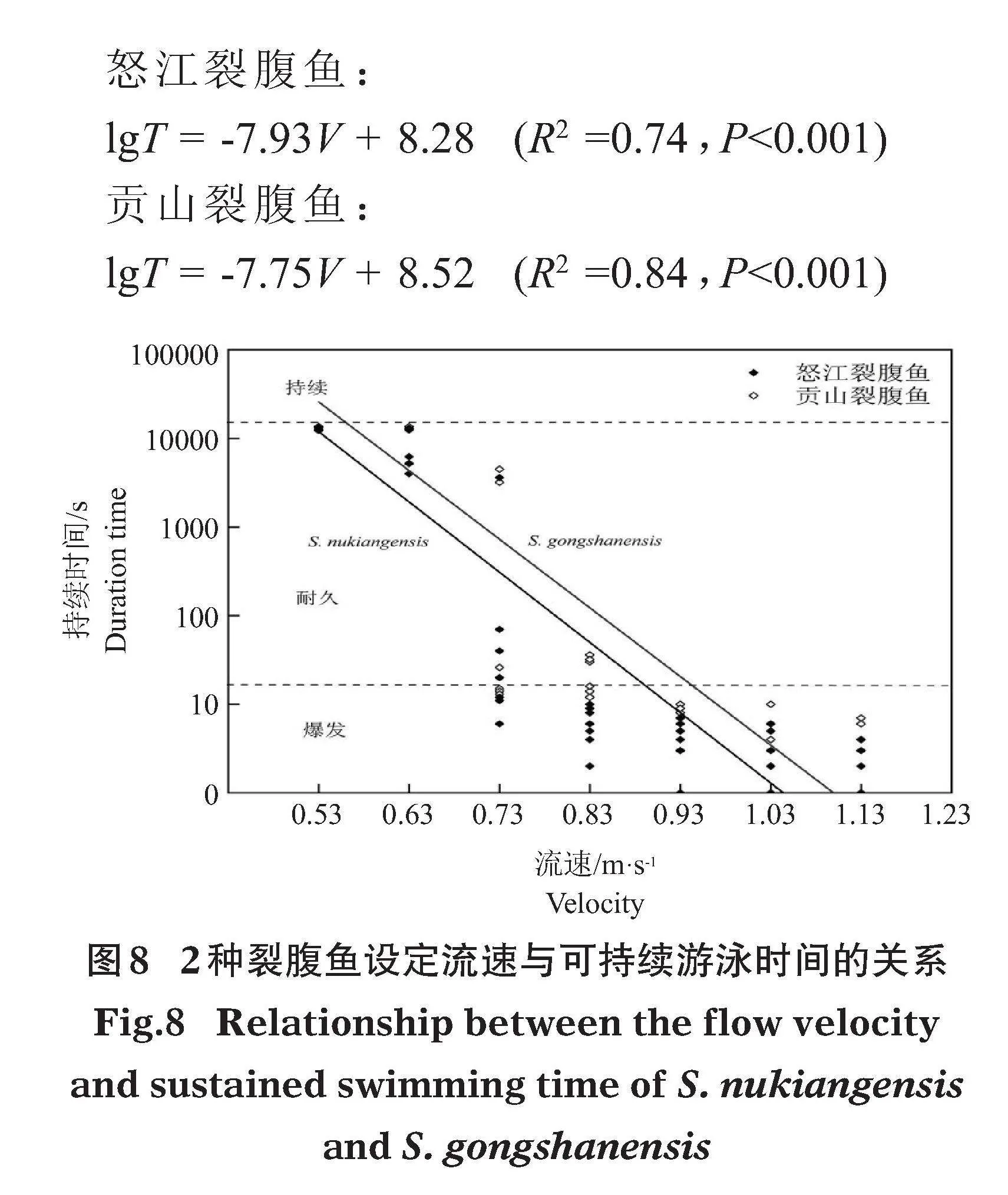
測試在設定流速0.53、0.63、0.73、0.83、0.93、1.03、1.13 m/s下進行。當設定流速調節至0.63 m/s時,有50%的試驗魚持續游泳時間大于200 min;當設定流速增至0.93 m/s時,50%的試驗魚持續游泳時間小于20 s。結果表明,怒江裂腹魚最大耐久游泳速度為0.93 m/s,最大持續游泳速度為0.63 m/s;貢山裂腹魚的最大耐久游泳速度為0.73 m/s,最大持續游泳速度為0.63 m/s。
分析2種裂腹魚設定流速與可持續游泳時間的關系(圖8),發現持續游泳時間與設定流速呈負相關關系,且持續游泳時間均呈現出明顯的兩極分化現象。持續游泳時間 (T,s) 和水流速度 (V,m/s) 的擬合關系分別為:
怒江裂腹魚:
lgT = -7.93V + 8.28"" (R2 =0.74,Plt;0.001)
貢山裂腹魚:
lgT = -7.75V + 8.52"" (R2 =0.84,Plt;0.001)
魚道長度與魚道內最大允許流速關系見圖 9。當魚道長度d為10~300 m時,根據2種裂腹魚在魚道內的最大允許流速與魚道長度關系曲線,得到魚道允許最大平均水流速度為0.52~0.70 m/s。
3"" 討論
3.1"" 不同魚類的游泳能力比較
本次試驗所用的2種裂腹魚棲息于急流險灘與回水灣沱密布的復雜生境中,對比發現二者的感應流速極為相近,該結果可能是由于2種裂腹魚親緣關系較近,且棲息于同一生境(丁少波等,2020)。將2種裂腹魚的絕對感應流速與河湖中生活的草魚絕對感應流速(8.21±0.68)cm/s、鰱感應流速(6.74±0.84)cm/s相比較(白艷勤等,2013),發現裂腹魚的感應流速更大。由于試驗裂腹魚大多生活在礫石、塊石形成的河流底質下,流場較為紊亂,且該調查河段上下游落差較大,水流湍急,因而2種裂腹魚的感應流速較大,而來自于養殖場的鰱與草魚,由于長期生活在相對靜止的水環境中,對水流的感知則較為敏銳;另外,雷青松等(2020)和李志敏等(2018)測試魚類感應流速前將其在靜水條件下適應,而本試驗在1.0 BL/s的條件下適應,可能導致了測得的感應流速偏大。本研究中,2種裂腹魚與新疆木扎提河流域同為裂腹魚屬的斑重唇魚(Diptychus maculatus)的絕對感應流速(0.18±0.02)m/s基本一致(雷青松等,2020);相似的水流環境及種屬是其原因之一(丁少波等,2020)。
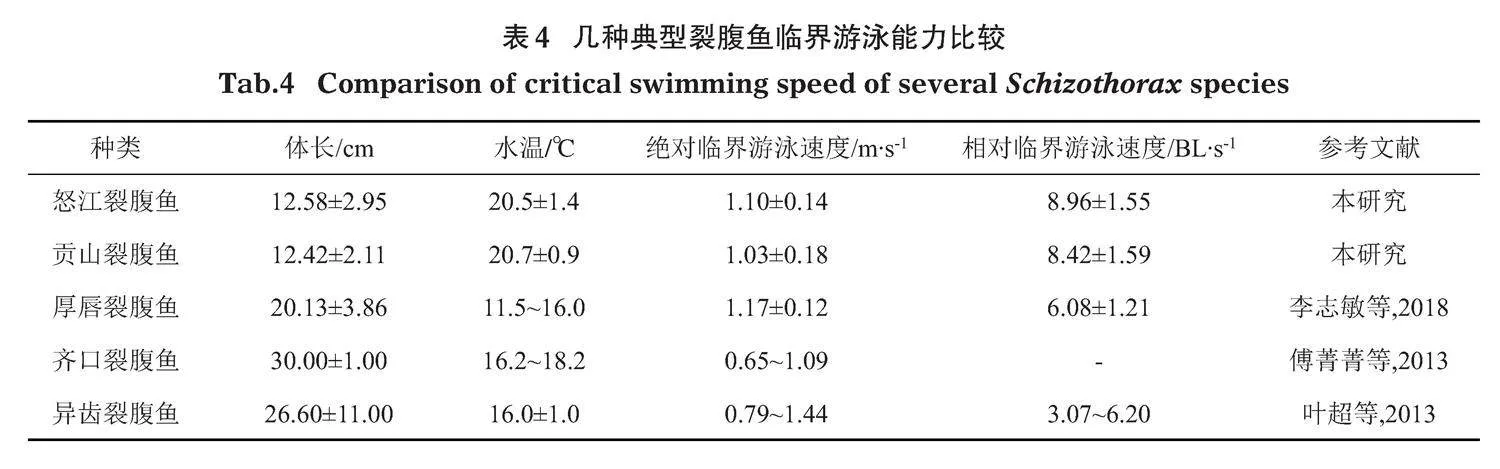
臨界游泳速度是魚類有氧運動評價的重要評價指標(Brett,1964)。目前,國內有很多學者研究過魚類的臨界游泳速度(表4)。本試驗測得的2種裂腹魚與厚唇裂腹魚(S. irregularis)的絕對臨界游泳速度較為接近,略微大于齊口裂腹魚(S. prenanti)。這可能與幾種魚的棲息環境以及試驗溫度及魚體規格等有關(田凱等,2010;李志敏等,2018;Tudorache et al,2007;Kern et al,2018)。
3.2"" 魚類游泳能力指標在魚道設計中的應用
魚道設計通常需考慮多種過魚對象的游泳能力。感應流速是指魚類剛剛能夠產生趨流反應的流速值,魚類的感應流速是魚道設計的重要參數之一,若魚道內設計流速過低,會導致魚類難以找到魚道入口,影響魚道的過魚效率。因此,魚道設計流速應大于主要過魚對象的感應流速(李志敏等,2018;崔忠波,2011);此外,對于其他類型的過魚設施,如升魚機及集運魚系統的上游放流點選擇,應參考過魚對象的感應流速(蔡露等,2018)。當以上述2種裂腹魚為主要過魚對象時,建議魚道內的最小控制流速不低于0.24 m/s。
一般情況下,魚類通過魚道依靠耐久游泳速度,但當穿越魚道進口或豎縫時采用突進游泳速度(李志敏等,2018)。根據魚類的趨流性,魚道進口或豎縫處的流速設計值應較大,但應小于過魚對象的突進游泳速度(曾海釗等,2017)。根據本次試驗結果,建議以2種裂腹魚為過魚對象時,魚道進口及豎縫處的流速應介于臨界游泳速度與突進游泳速度之間,即1.05~1.40 m/s;若要保證80%的魚可通過魚道成功上溯,則魚道豎縫處流速最大不宜超過1.23 m/s。魚類進入魚道內部后,若魚道內設計流速過小,將迷失方向,建議魚道內流速應在感應流速和臨界游泳速度之間,即魚道內整體平均流速為0.24~1.05 m/s。值得注意的是,若要保證更大規格的魚通過魚道,魚道內的最低設計流速可適當提高。
突進游泳是魚類穿越高速水流時的游泳行為(葉超等,2013;Watkins,1996;Plaut,2001)。突進游泳速度是魚類進行無氧運動的重要指標(Balley,2003)。對該試驗突進游泳能力測試結果表明,怒江裂腹魚和貢山裂腹魚的絕對突進游泳速度均有隨體長的增大而呈現出正相關的趨勢,怒江裂腹魚比貢山裂腹魚的趨勢更加明顯,二者相對突進游泳速度均隨體長的增大而呈現出負相關趨勢,這與其他學者的測試結果類似(葉超等,2013;王永猛等,2020)。在魚道設計方面,高速流區(主要有魚道進口處、內孔口及豎縫處)的設計流速應小于過魚對象的突進游泳速度(鄭金秀等,2010)。綜合本試驗測得的臨界游泳速度及突進游泳速度,當以怒江裂腹魚和貢山裂腹魚為主要過魚對象時,建議魚道高流速區最大流速應小于1.40 m/s。
自然狀態下,魚類多采用持續式游泳以維持較低的代謝水平(Reidy et al,2000)。本試驗測得怒江裂腹魚和貢山裂腹魚的最大持續游泳速度均為0.63 m/s。耐久游泳是魚類游至疲勞的游泳類型,該狀態下魚類兼具無氧代謝和有氧代謝,直至魚體內乳酸積累至疲勞(何大仁和蔡厚才,1998)。本試驗測得怒江裂腹魚和貢山裂腹魚最大耐久游泳速度均為0.73 m/s。
在魚道設計中,通常會設置休息池以恢復魚類體能,池內的主流流速過大過小都會對過魚效率產生不利影響,即主流水流速度介于過魚對象感應流速和臨界游泳速度之間(于曉明等,2017)。依據試驗結果綜合考慮,建議以怒江裂腹魚和貢山裂腹魚為主要過魚對象時,休息池內的主流流速應設為0.18~1.05 m/s。
參考文獻
白艷勤,路波,羅佳,等,2013. 草魚、鰱和瓦氏黃顙魚幼魚感應流速的比較[J]. 生態學雜志, 32(8):2085-2089.
大唐玉曲河水電開發有限公司, 2018. 西藏玉曲河扎拉水電站環境影響報告書[R]. 武漢:長江水資源保護科學研究所.
蔡露,侯軼群,金瑤,等,2021. 魚游泳能力對體長的響應及其在魚道設計中的應用[J]. 農業工程學報, 37(5):209-215.
曹亮,張鶚,臧春鑫,等,2016. 通過紅色名錄評估研究中國內陸魚類受威脅現狀及其成因[J]. 生物多樣性, 24(5):598-610.
曾海釗,周小波,陳子海,2017. 藏木水電站魚道設計[J]. 水電站設計, 33(1):68-71.
陳鋒,陳毅鋒,2010. 拉薩河魚類調查及保護[J]. 水生生物學報, 34(2):278-283.
崔忠波,2011. 《水利水電工程魚道設計導則》(送審稿) 審查會在北京召開[J]. 水利技術監督,19(6):29.
丁少波,施家月,黃濱,等,2020. 大渡河下游典型魚類的游泳能力測試[J]. 水生態學雜志, 41(1):46-52.
馮克義,2019. 大渡河水電站開發中水土流失環境問題分析[J]. 水利水電技術, 50(S2):229-231.
傅菁菁,李嘉,安瑞冬,等,2013. 基于齊口裂腹魚游泳能力的豎縫式魚道流態塑造研究[J]. 四川大學學報(工程科學版), 45(3):12-17.
何大仁,蔡厚才,1998. 魚類行為學[M]. 廈門:廈門大學出版社.
侯軼群,蔡露,陳小娟,等,2020. 過魚設施設計要點及有效性評價[J]. 環境影響評價, 42(3):19-23.
雷青松,涂志英,石迅雷,等,2020. 應用于魚道設計的新疆木扎提河斑重唇魚的游泳能力測試[J]. 水產學報, 44(10):1718-1727.
李雷,馬波,金星,等,2019. 西藏雅魯藏布江中游裂腹魚類優先保護等級定量評價[J]. 中國水產科學, 26(5):914-924.
李志敏,陳明曦,金志軍,等,2018. 葉爾羌河厚唇裂腹魚的游泳能力[J]. 生態學雜志, 37(6):1897-1902.
劉潔雅,2016. 西藏巨須裂腹魚個體生物學和種群動態研究[D]. 阿拉爾:塔里木大學.
田凱,曹振東,付世建,2010. 速度增量及持續時間對瓦氏黃顙魚幼魚臨界游泳速度的影響[J]. 生態學雜志, 29(3):534-538.
王龍濤,2015. 怒江上游水電開發對魚類棲息環境影響分析及保護[D]. 武漢:華中農業大學.
王永猛,柯森繁,林晨宇,等,2021. 紅河(元江)流域的典型魚類游泳能力探究及在過魚設施流速設計中的應用[J]. 湖泊科學, 33(6):1820-1831.
王永猛,李志敏,涂志英,等,2020. 基于雅礱江兩種裂腹魚游泳能力的魚道設計[J]. 應用生態學報, 31(8):2785-2792.
西藏自治區水產局,1995. 西藏魚類及其資源[M]. 北京:中國農業出版社.
徐革鋒,尹家勝,韓英,等,2015. 不同適應條件對細鱗鮭幼魚游泳能力的影響[J]. 生態學報, 35(6):1938-1946.
葉超,王珂,黃福江,等,2013. 異齒裂腹魚游泳能力初探[J]. 淡水漁業, 43(3):33-37.
于曉明,崔聞達,陳雷,等,2017. 水溫、鹽度和溶氧對紅鰭東方鲀幼魚游泳能力的影響[J]. 中國水產科學, 24(3):543-549.
于曉明,張秀梅,張沛東,2009. 凡納濱對蝦的游泳能力及游泳疲勞后的生理反應[J]. 中國水產科學, 16(4):533-540.
鄭金秀,韓德舉,胡望斌,等,2010. 與魚道設計相關的魚類游泳行為研究[J]. 水生態學雜志, 3(5):104-110.
周小波,陳靜,張連明,2017. 淺論魚道設計方法與過程——以藏木水電站魚道設計為例[J]. 水電站設計, 33(3):29-32.
Balley D M, Bagley P M, Jamieson A J, et al, 2003. In situ investigation of burst swimming and muscle performance in the deepsea fish Antimora rostrata[J]. J Exp Mar Biol Ecol, 285:295-311.
Brett J R, 1964. The respiratory metabolism and swimming performance of young sockeye salmon[J]. Journal of the Fisheries Research Board of Canada, 21(5):1183-1226.
Hammer C, 1995. Fatigue and exercise tests with fish[J]. Comparative Biochemistry and Physiology-Part A: Physiology, 112(1):1-20.
Katopodis C, 2005. Developing a toolkit for fish passage, ecological flow management and fish habitat works[J]. Journal of Hydraulic Research, 43(5):451-467.
Kern P, Cramp R L, Gordos M A, et al, 2018. Measuring Ucrit and endurance: equipment choice influences estimates of fish swimming performance[J]. Journal of Fish Biology, 92(1):237-247.
Peake S, Beamish F W H, Mckinley R S, et al, 1997. Relating swimming performance of lake sturgeon, Acipenser fulvescens, to fishway design[J]. Canadian Journal of Fisheries and Aquatic Sciences, 54(6):1361-1366.
Plaut I, 2001. Critical swimming speed: its ecological relevance[J]. Comparative Biochemistry and Physiology-Part A: Molecular amp; Integrative Physiology, 131(1):41-50.
Reidy S P, Kerr S R, Nelson J A, 2000. Aerobic and anaerobic swimming performance of individual Atlantic cod[J]. Journal of Experimental Biology, 203:347-357.
Smit H, Amelink-Koutstaal J M, Vijverberg J, et al, 1971. Oxygen consumption and efficiency of swimming goldfish[J]. Comparative Biochemistry and Physiology-Part A: Physiology, 39(1):1-28.
Tan J J, Chen L, Tan H L, et al, 2022. Identifying optimal position for a fish collection system for endemic fishes in Hong River, China[J]. Ecological Engineering, 176:106524.
Tudorache C, Viaenen P, Blust R, et al, 2007. Longer flumes increase critical swimming speeds by increasing burstglide swimming duration in carp Cyprinus carpio, L.[J]. Journal of Fish Biology, 71(6):1630-1638.
Watkins T B, 1996. Predator-mediated selection on burst swimming performance in tadpoles of the Pacific tree frog, Pseudacris regilla[J]. Physiol Zool, 69(1):154-167.
(責任編輯"" 萬月華)
Swimming Ability of Schizothorax in the Yuqu River Basin and Its Application
in the Design of Fish Passage Facilities
LIU Han‐wen1,2, TAN Jun‐jun1,2, WANG Yong‐meng3, LI Yang‐xi1,2, WANG Meng3, KE Sen‐fan1,2,
SHI Xiao‐tao1,2, TAN Hong‐lin1,2, WANG Yuan‐yang1,2, LI Wei‐dong1,2, PU Jin4, JIN Zhi‐jun3
(1. College of Hydraulic and Environmental Engineering, China Three Gorges University,
Yichang, Hubei"" 443002, P. R. China;
2. Hubei International Science and Technology Cooperation Base of Fish Passage,
Three Gorges University, Yichang"" 443002, P. R. China;
3. Power China Guiyang Engineering Corporation Ltd., Guiyang"" 550081, P. R. China;
4. The Branch of China Datang Corporation Ltd in Tibet, Tibet, Changdu"" 854000, P. R. China)
Abstract:In this study, the dominant species in the Yuqu River basin, Schizothorax nukiangensis and Schizothorax gongshanensis, were selected for research. Swimming capability was assessed by measuring induction flow velocity, critical swimming speed, burst swimming speed, sustainable swimming speed and endurance swimming speed. The objective of the study was to provide basic parameters for fishway design. In June 2021, the test fish were collected in Yuqu River, with a body length range of 7.9-22.5 cm and body weight range of 5.3-145.0 g for S. nukiangensis and of 8.1-21.4 cm and 9.9-135.6 g for S. gongshanensis. After acclimation, the swimming ability of the fish were tested in an open flume using the stepped velocity method. The absolute induction velocities of S. nukiangensis and S. gongshanensis were similar, with values of (0.18±0.01) m/s and (0.18±0.06) m/s, respectively. The absolute critical swimming speeds of S. nukiangensis and S. gongshanensis were (1.05±0.12) m/s and (1.11±0.14) m/s, respectively, and their respective absolute burst swimming speeds were (1.45±0.26) m/s and (1.40±0.21) m/s. The respective relative critical swimming speeds were (8.47 ±0.95) BL/s and (9.21±2.45) BL/s and the respective relative burst swimming speeds were (11.90±2.31) BL/s and (10.80±3.16) BL/s, both exceeding 10 times their body length. The measured maximum sustainable swimming speeds of S. nukiangensis and S. gongshanensis were both 0.63 m/s, and the maximum endurance swimming speeds of S. nukiangensis and S. gongshanensis were 0.93 m/s and 0.73 m/s, respectively. When S. nukiangensis and S. gongshanensis are selected as target fish species for a fish passage, the velocities at the entrance and vertical slots should be 1.05-1.40 m/s, the average velocity inside the fish passage should be 0.24-1.05 m/s, and the velocity in the resting pools should be 0.18-1.05 m/s. Our results provide a reference for designing fish passages in the Yuqu River basin of Tibet, and is important for fish resource protection.
Key words:Schizothorax nukiangensis; Schizothorax gongshanensis; induction flow velocity; critical swimming speed; burst swimming speed; fishway design; Yuqu River
