氮添加對(duì)稀土尾砂地猴樟幼苗根系生長、生物量分配及非結(jié)構(gòu)性碳水化合物的影響
2024-01-02 11:21:56陳志琪張海娜劉佳麗魯向暉楊寶城
植物研究 2024年1期
陳志琪 張海娜 劉佳麗 魯向暉 楊寶城
(南昌工程學(xué)院江西省樟樹繁育與開發(fā)利用工程研究中心,南昌 330099)
我國南方贛南、閩西、粵北等地區(qū)擁有大量的稀土資源。自20 世紀(jì)80 年代以來,由于工業(yè)產(chǎn)品的大量需求,稀土資源被大規(guī)模開發(fā),經(jīng)歷了池浸、堆浸、原位浸礦等無序開采工藝,導(dǎo)致當(dāng)?shù)刂参镌獾酱竺娣e破壞,遺留大面積的稀土尾礦廢棄地[1]。這些廢棄尾砂土壤結(jié)構(gòu)遭到嚴(yán)重破壞,保水保肥能力差,且土壤酸化,營養(yǎng)元素缺乏[2],導(dǎo)致尾礦地植被恢復(fù)受限。已有研究表明,樟樹(Cinnamomum camphora)可以在稀土尾砂上種植[3]。樟樹作為江西省的特色經(jīng)濟(jì)植物,其作為香料的應(yīng)用價(jià)值得到廣泛關(guān)注。猴樟(C.bodinier)作為樟科(Lauraceae)樟屬(Cinnamomum)的常綠喬木,其枝葉富含精油,目前以矮林栽培方式種植,每年地上部分被平茬提油。為了滿足新一輪枝葉的連續(xù)生長,每年需補(bǔ)充大量氮肥,而尾砂保肥能力差,多余氮肥會(huì)造成更為嚴(yán)重的水土流失和面源污染問題。因此,在稀土尾砂植被恢復(fù)中,需要更為科學(xué)的施肥管理。
氮素作為植物的生命元素,影響著植物生長、營養(yǎng)元素的吸收利用和能量物質(zhì)的貯藏等[4]代謝過程。通常情況,植物生長受氮限制時(shí),氮添加有益于植物生長[5];而在氮相對(duì)豐富的區(qū)域,氮添加反而會(huì)抑制植物生長,降低植物生產(chǎn)力[6]。例如:高鈿惠等[7]研究發(fā)現(xiàn)不同氮水平對(duì)中金楊(Populus×euramericana‘Zhongjin7’)幼苗的生長促進(jìn)效果不同,其中高氮水平會(huì)抑制植物根系生長,影響生物量的積累;高晉麗等[8]的研究顯示,高濃度氮素添加能夠顯著促進(jìn)大興安嶺凍土區(qū)表層土壤(0~ 20 cm)中植物細(xì)根的生長;董姣姣等[9]和裴昊斐等[10]的研究發(fā)現(xiàn)適量添加氮會(huì)促進(jìn)植物的生長和生物量的積累,并顯著提高植物的生產(chǎn)能力,但更高水平的氮添加可能對(duì)植物生長產(chǎn)生一定抑制作用。由此可見,不同環(huán)境中植物對(duì)氮元素的需求量不同。稀土尾砂地土壤環(huán)境相對(duì)缺氮,在稀土尾砂地進(jìn)行氮添加是否會(huì)影響植物體內(nèi)營養(yǎng)物質(zhì)的儲(chǔ)存與轉(zhuǎn)化目前尚不清楚。
非結(jié)構(gòu)性碳水化合物(Non-structural carbohydrates,NSC)以可溶性糖和淀粉的形式為植物生長提供所需要的能量[11],其中可溶性糖(Soluble sugars,SS)是植物體內(nèi)碳水化合物運(yùn)輸?shù)闹饕问剑矸郏⊿tarch,St)是植物碳的主要儲(chǔ)存物質(zhì),兩者在一定條件下可以相互轉(zhuǎn)換以維持正常的生理活動(dòng)[12]。植物葉片光合作用新合成的碳和同化物,通過維管束運(yùn)輸,轉(zhuǎn)移至莖稈或根中,當(dāng)環(huán)境變化時(shí),非結(jié)構(gòu)性碳水化合物在植物不同組織的分配可能會(huì)發(fā)生變化,進(jìn)而影響植物的生長和對(duì)環(huán)境變化的響應(yīng)策略[13]。以往研究表明,缺氮時(shí)植物會(huì)增加分配到根系的碳水化合物比例,以促進(jìn)氮的吸收[14],而氮有效性的增加則會(huì)導(dǎo)致更多的碳水化合物分配到地上組織[15],對(duì)杉木(Cunninghamia lanceolate)來說,氮添加則會(huì)降低葉片可溶性糖和NSC 含量[16];而史增順等[17]的研究表明,氮添加會(huì)使細(xì)根比根長、比表面積增大,但NSC 含量降低,尤其是淀粉含量。造成這些影響的原因可能與植物對(duì)氮素的敏感性和具體的施氮水平有關(guān)。
基于此,本研究以尋烏縣某典型稀土礦尾砂為基質(zhì),通過1 年生猴樟扦插幼苗的盆栽試驗(yàn),研究氮添加對(duì)稀土尾砂基質(zhì)中猴樟幼苗生長及非結(jié)構(gòu)性碳水化合物含量的影響,為離子型稀土礦尾砂地生態(tài)修復(fù)提供理論參考。
1 材料與方法
1.1 研究區(qū)概況
試驗(yàn)在南昌工程學(xué)院生物技術(shù)試驗(yàn)基地進(jìn)行(28°41′40.85″N,116°1′41.18″E)。該基地位于南昌市,屬亞熱帶濕潤季風(fēng)氣候,年降雨量1 600~1 700 mm,4—6 月為雨季,7—9 月為季節(jié)性干旱期,年平均相對(duì)濕度為78.5%,年日照時(shí)間為1 723~1 820 h。
供試植物為長勢(shì)良好且均一的1 年生猴樟扦插幼苗,供試尾砂取自江西省贛州市尋烏縣某廢棄稀土礦。稀土尾砂的理化性質(zhì)如下:土壤pH=5.08,屬于強(qiáng)酸性,電導(dǎo)率高達(dá)79.12 dS·m-1,尾砂中土壤有機(jī)碳、有機(jī)質(zhì)、全氮、全磷質(zhì)量分?jǐn)?shù)分別為6.29、10.85、0.20、1.21 g·kg-1,速效磷和速效鉀質(zhì)量分?jǐn)?shù)分別為13.78、225.97 mg·kg-1。
1.2 試驗(yàn)設(shè)計(jì)
本研究采用盆栽控制試驗(yàn)。2022 年4 月初將30株長勢(shì)均一的1年生猴樟扦插幼苗(株高28 cm,地徑0.5 mm)移栽至底部開孔的塑料盆中(盆口直徑29 cm,盆高16 cm),每盆1 株,盆中裝入供試尾砂土8 kg,定期澆水,緩苗1 個(gè)月。為了促進(jìn)香料用樟樹的生長,當(dāng)?shù)剞r(nóng)民會(huì)在樟樹平茬之后進(jìn)行施肥。本研究以硝酸銨鈣為N源(含N 15%),共設(shè)置3個(gè)氮肥添加處理,分別為CK(0)、N1(1.8 g·株-1)、N2(3.6 g·株-1)。試驗(yàn)設(shè)計(jì)中,N1 施氮量根據(jù)當(dāng)?shù)剞r(nóng)民常用的施肥量來定,并換算成單一元素的化肥施用量,為1.8 g·株-1;N2 施氮量根據(jù)馮瑜等[18]的研究來確定,發(fā)現(xiàn)氮、磷、鉀施用量為2.5 g·株-1時(shí)對(duì)樟樹生長的促進(jìn)作用明顯,產(chǎn)生的水土流失量較小,經(jīng)換算后N2 的施肥量定為3.6 g·株-1。試驗(yàn)采用單因子三水平隨機(jī)試驗(yàn),每個(gè)處理10 盆,于當(dāng)年5—8 月進(jìn)行氮添加試驗(yàn)。施肥時(shí),在樟樹幼苗周圍開挖環(huán)狀施肥溝,將硝酸銨鈣溶于200 mL 水均勻施入溝中,每月定期1 次。試驗(yàn)期間不同處理的植株隨機(jī)擺放。生長期末,即10月15 日對(duì)所有植株進(jìn)行破壞性采樣,然后將每株植物分為根、莖、葉,分別裝入自封袋中,帶回實(shí)驗(yàn)室進(jìn)行分析。
1.3 測(cè)定方法
根性狀測(cè)定:根系采用“全根法”進(jìn)行挖掘。首先將地上部分齊地剪斷,然后沿著每條根的走向,用鑷子和小鏟子挖出根系并用清水沖洗干凈,最后將粗根(直徑>2 mm)和細(xì)根(直徑≤2 mm)分開。根性狀指標(biāo)通過根系掃描儀(Epson Perfection V850 Pro,中國)和根系分析系統(tǒng)WinRHIZO獲得,包括根長、根表面積、根直徑。
生物量測(cè)定:將植物不同組織和掃描后的根系分別裝入信封標(biāo)記,105 ℃殺青30 min,之后80 ℃烘至恒質(zhì)量,獲得植株不同組織的生物量。其中,總生物量為各組織生物量之和;冠層生物量為葉、莖生物量之和;不同組織生物量占比為各組織生物量與總生物量之比。由于樟樹葉片中的精油含量最高,而葉莖比反映了植物枝葉間光合產(chǎn)物的分配和積累狀況,是樟樹精油含量的重要指標(biāo),因此本試驗(yàn)增加了葉莖比指標(biāo)。比根長(SRL)用根長/根干質(zhì)量表示;比表面積(SRA)用根表面積/根干質(zhì)量表示。
NSC 測(cè)定:NSC 包括可溶性糖和淀粉[19]。將烘干后的植物組織粉碎研磨、過0.25 mm 篩,用于NSC 含量測(cè)定。NSC 含量采用硫酸蒽酮法[20]測(cè)定。
1.4 數(shù)據(jù)處理
數(shù)據(jù)的統(tǒng)計(jì)分析用SPSS 26.0 進(jìn)行,利用單因素方差分析檢驗(yàn)氮添加對(duì)植株根系性狀、總生物量和不同組織的生物量及NSC 含量的差異顯著性。作圖通過Origin 2018完成。
2 結(jié)果與分析
2.1 氮添加對(duì)猴樟幼苗生物量及分配的影響
不同水平的氮添加均能促進(jìn)猴樟幼苗各組織及總生物量的增加,其中葉、莖、粗根、細(xì)根、冠層和總生物量均在N1 處理下最高,分別比CK 高392.78%、86.95%、116.17%、99.39%、170.99%、149.99%,也顯著高于N2(P<0.05);N2 處理下的莖、根和總生物量與CK 無顯著差異(P>0.05)。根冠比在N1 和N2 水平下分別降低了8.33%、31.67%,葉莖比在N1 和N2 處理下分別比CK 顯著增加了163.16%、213.16%(P<0.05)(見表1)。
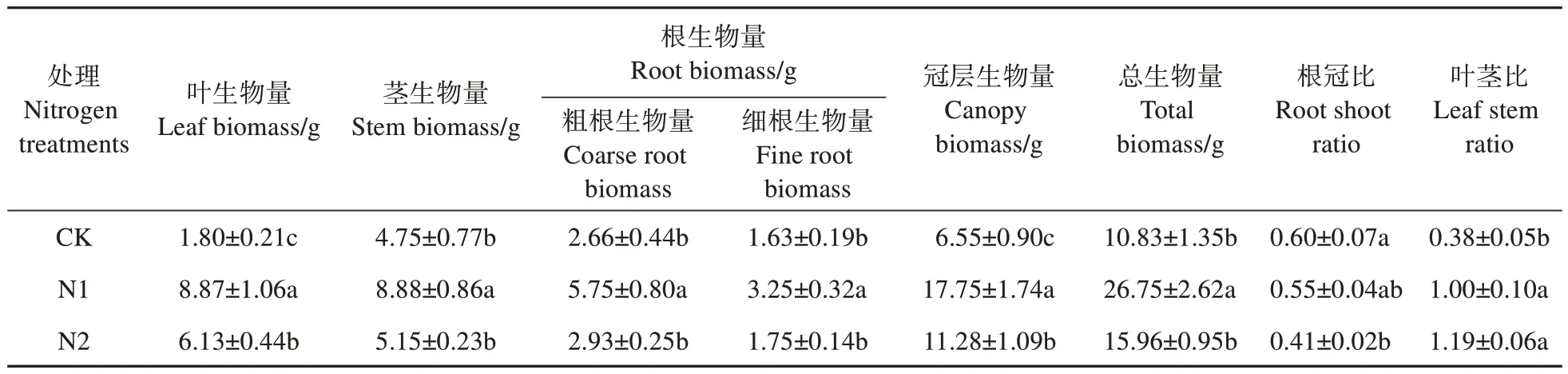
表1 不同氮水平對(duì)猴樟幼苗生物量的影響Table 1 Effects of different nitrogen levels on biomass of C. bodinieri seedlings
與CK 相比,葉比重在N1 和N2 水平下顯著增加了123.53%、92.94%(P<0.05);N1 和N2 水平下,莖比重分別降低了9.46%、19.26%,而兩者之間無顯著差異(P>0.05),粗根比重較CK 分別顯著降低了17.90%、27.63%(P<0.05),細(xì)根比重分別顯著降低了27.16%、32.10%(P<0.05)(見表1、圖1)。

圖1 不同氮水平下猴樟幼苗生物量的分配不同處理間,同一種生物量上的不同小寫字母表示差異顯著;下同。Fig.1 Biomass allocation of C. bodinieri seedlings under different nitrogen levels Different lowercase letters indicate significant differences among treatments(P<0.05);the same as below.
2.2 氮添加對(duì)猴樟幼苗根系生長的影響
與CK 相比,N1 和N2 水平下猴樟幼苗的粗根根長分別顯著增加了85.66%、57.49%(P<0.05),不同施氮水平間無顯著性差異(P>0.05)(見圖2A);粗根表面積在N1 處理時(shí)顯著高于CK 與N2(P<0.05)(見圖2B);粗根直徑在施氮條件下均低于CK,不同施氮水平間差異不顯著(P>0.05)(見圖2C);粗根比根長在N2 水平下分別比CK、N1增加了81.19%、23.27%(見圖2D);粗根比表面積在N2 水平下顯著高于CK 和N1(P<0.05)(見圖2E)。
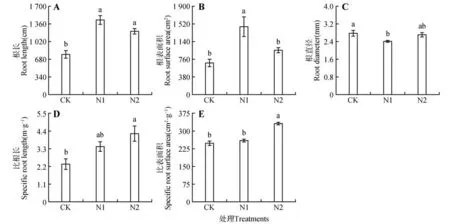
圖2 不同氮水平下猴樟幼苗粗根性狀Fig.2 Coarse root traits of C. bodinieri seedlings under different nitrogen levels
與CK 相比,細(xì)根根長在N1 和N2 水平下分別顯著增加了123.81%、73.27%(P<0.05)(見圖3A);細(xì)根表面積在N1 和N2 水平下分別比CK 顯著增加了69.66%、46.93%(P<0.05)(見圖3B);細(xì)根直徑在N1條件下均低于CK和N2,且與CK有明顯差異(P<0.05)(見圖3C);細(xì)根比根長在N2 處理下最大,顯著高于CK(P<0.05),不同施氮水平間差異不顯著(P>0.05)(見圖3D);細(xì)根比表面積在N2處理時(shí)最大,顯著高于CK(P<0.05)(見圖3E)。
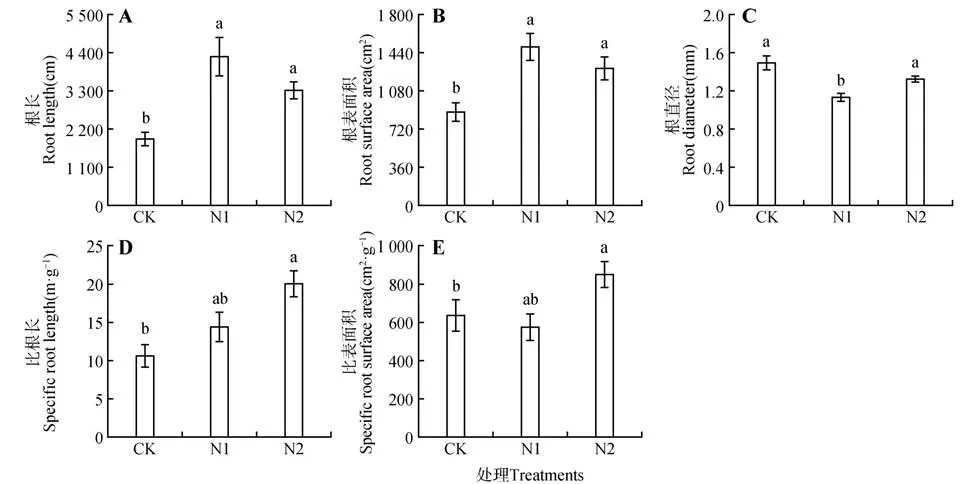
圖3 不同氮水平下猴樟幼苗細(xì)根性狀Fig.3 Fine root traits of C. bodinieri seedlings under different nitrogen levels
2.3 氮添加對(duì)猴樟幼苗非結(jié)構(gòu)性碳水化合物的影響
不同氮添加處理對(duì)猴樟幼苗的葉、莖、粗根、細(xì)根4個(gè)組織的可溶性糖質(zhì)量分?jǐn)?shù)均有影響(見圖4:A~D)。葉、莖可溶性糖質(zhì)量分?jǐn)?shù)均在N1 處理下質(zhì)量分?jǐn)?shù)最高,N2處理最低,N1處理下葉、莖中可溶性糖質(zhì)量分?jǐn)?shù)分別比CK、N2 提升了9.14%、26.59%、1.13%、13.56%(見圖4:A~B)。此外,氮添加也會(huì)影響猴樟幼苗各組織的淀粉質(zhì)量分?jǐn)?shù)(見圖4:E~H)。N1 處理下的葉淀粉質(zhì)量分?jǐn)?shù)最低,分別比CK、N2 降低了43.14%、29.17%(見圖4E),并且CK 與N1 間差異顯著(P<0.05);莖中的淀粉質(zhì)量分?jǐn)?shù)在N2處理下最低,分別比CK、N1降低了39.40%、6.01%(見圖4F);粗根中的淀粉質(zhì)量分?jǐn)?shù)以N2 最低,比N1 顯著降低37.93%(P<0.05)(見圖4G)。N1、N2中的葉NSC質(zhì)量分?jǐn)?shù)分別比CK降低6.43%、12.28%(見圖4I)。莖中的NSC質(zhì)量分?jǐn)?shù)以N2 處理最低,比CK 顯著降低了14.29%(P<0.05)(見圖4J)。N2 處理下粗根、細(xì)根中的NSC 較CK顯著降低了53.41%、46.49%(P<0.05)(見圖4:K~L)。葉的NSC 質(zhì)量分?jǐn)?shù)要明顯高于莖、粗根與細(xì)根(見圖4:I~L)。猴樟幼苗葉片的可溶性糖質(zhì)量分?jǐn)?shù)占NSC 的77.41%~93.21%,淀粉質(zhì)量分?jǐn)?shù)為13.18%~22.59%,莖、粗根與細(xì)根的表現(xiàn)相同。
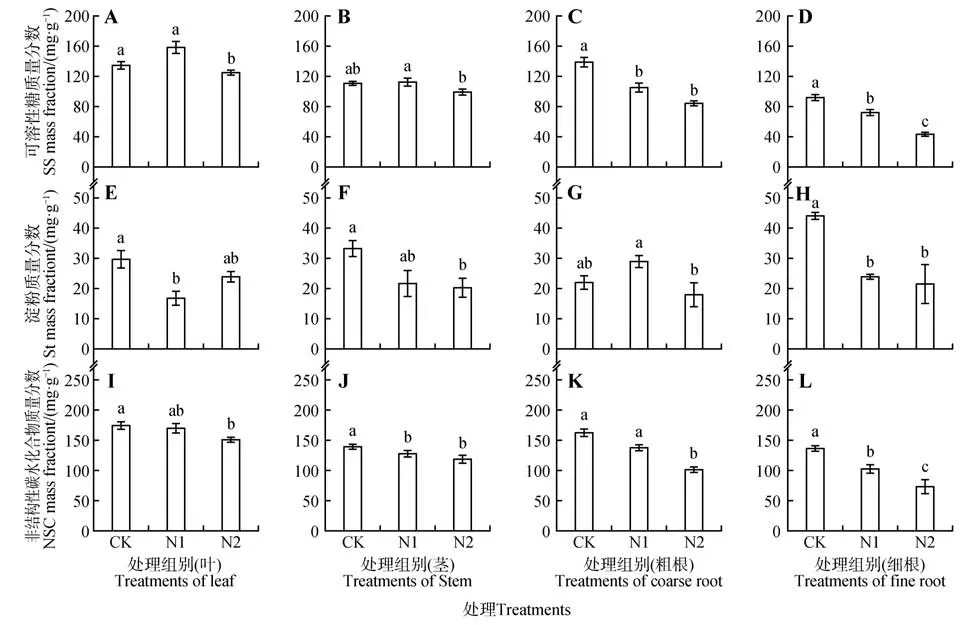
圖4 不同氮水平對(duì)猴樟幼苗非結(jié)構(gòu)性碳水化合物的影響Fig.4 Effects of different nitrogen levels on non-structural carbohydrates of C. bodinieri seedlings
3 討 論
3.1 氮添加對(duì)猴樟幼苗生物量及分配的影響
生物量是光合產(chǎn)物積累的結(jié)果,氮的供應(yīng)狀況會(huì)影響植物對(duì)碳同化物質(zhì)的分配格局和生物量的積累[21]。本研究中,猴樟幼苗葉、莖、粗根、細(xì)根生物量均在N1 處理下最高,顯著高于N2 處理(P<0.05),說明低氮條件有利于促進(jìn)猴樟幼苗的生物量積累,這與李樹萍等[22]對(duì)樹番茄(Cyphomandra betacea)的研究結(jié)果相似,樹番茄的生物量在低氮條件下最高。齊成媚等[23]的研究發(fā)現(xiàn),高氮水平對(duì)假臭草(Praxelis clematidea)生物量累積的效果最佳,這與本研究結(jié)果不同,原因可能是假臭草屬于草本植物,對(duì)環(huán)境要求不高,且對(duì)土壤肥力的吸收能力較強(qiáng),猴樟是木本植物,兩者對(duì)氮素的響應(yīng)不同。氮添加有利于提升植物光合作用產(chǎn)生有機(jī)物的能力,使植株生物量的積累效率得到提高[24]。猴樟幼苗的葉、莖生物量在N1、N2 處理時(shí)均高于CK,并且N1 處理的提升效果顯著高于N2(P<0.05),說明N1條件下的氮添加量更能促進(jìn)猴樟幼苗葉、莖生物量的積累,表明不同濃度氮添加處理下植物對(duì)養(yǎng)分吸收效率的提升效果不同。毛晉花等[25]的研究表明也說明了這一點(diǎn)。本研究還發(fā)現(xiàn),N1 處理下猴樟幼苗的粗根、細(xì)根生物量均顯著高于CK(P<0.05),而N2 處理時(shí)與CK 無明顯差異(P>0.05),且N1處理的根生物量均顯著高于N2(P<0.05)。這可能是因?yàn)榈蜐舛鹊砑樱∟1)條件下猴樟幼苗根的生長不會(huì)受限,而高濃度氮添加(N2)時(shí),尾砂中的可利用氮源大幅增加,植物會(huì)減少葉片向根系分配的碳量,導(dǎo)致根發(fā)育所需的養(yǎng)分缺失,從而抑制根的生長。
氮素營養(yǎng)的改變還影響植物體內(nèi)的碳分配格局[26]。植物不同組織中的生物量占比反映了植株分配給各組織生物量的比例,是判斷植物生長狀況的重要指標(biāo)。本研究中,猴樟幼苗的葉比重在N2 處理時(shí)最高,說明相比于N1 添加水平,氮濃度更高的N2 處理更有利于葉生物量積累。這與王立冬等[27]對(duì)木荷(Schima superba)的研究結(jié)果相似,其研究結(jié)果表明,較高銨態(tài)氮濃度處理顯著提升了木荷的葉生物量。根冠比反映了生物量在地上和地下部分之間分配的情況。本研究結(jié)果顯示,猴樟幼苗根比重、根冠比在施氮條件下均低于CK,且N2處理差異最顯著(P<0.05)。而高歌等[28]的研究發(fā)現(xiàn),與不施氮相比,施氮后杉木(Cunninghamia lanceolate)、青岡(Quercus glauca)的根生物量比、根冠比均顯著降低(P<0.05)。氮添加可能會(huì)增加土壤中N 的有效性[29],隨著氮的施入,植物生長的資源限制由氮變成碳。因此,植物需要更多的葉片來獲取更多的二氧化碳,從而導(dǎo)致植物的根系發(fā)育受到抑制,使植物根生物量下降。吳茜等[30]的研究顯示氮添加后楓香(Liquidambar formosana)、木荷(Schima superba)、禿瓣杜英(Elaeocarpus glabripetalus)和青岡的根冠比與根比重均顯著低于CK(P<0.05);而王曉榮等[31]發(fā)現(xiàn)氮沉降能夠顯著提升枹櫟(Quercus glandulifera)的根冠比(P<0.05)。這些研究結(jié)果表明了不同植物間的根冠比對(duì)于氮添加的響應(yīng)具有一定的差異。本研究發(fā)現(xiàn),猴樟幼苗的莖、粗根、細(xì)根占比、以及根冠比在N2 水平下最低,而葉莖比在N2 時(shí)最高,說明高氮水平下,尾砂中氮元素含量較充足,植物會(huì)減少向根系分配碳元素的比例,從而改變植物地面部分與地下部分的碳分配格局[32]。
3.2 氮添加對(duì)猴樟幼苗根系生長的影響
根長、根表面積、根直徑等根系形態(tài)指標(biāo)與植物的生長發(fā)育有著密切聯(lián)系,因其具有對(duì)環(huán)境變化敏感且可塑性強(qiáng)等特點(diǎn),常被用于分析植物對(duì)環(huán)境的適應(yīng)能力[33]。在逆境環(huán)境中,植物可以提高根系的碳投入量(如提高根冠比或總根長)、減小根系直徑來降低根系的構(gòu)建成本,以擴(kuò)大根吸收面積來適應(yīng)環(huán)境[34]。本研究中,無論粗根還是細(xì)根,其根長和根表面積在不同氮濃度處理下均有所增加,而根直徑存在降低的情況,且N1 處理下的根直徑均低于CK 與N2,這與高晉麗等[8]的研究結(jié)果相似。植物對(duì)營養(yǎng)元素的吸收主要通過細(xì)根來實(shí)現(xiàn),細(xì)根比根長與比表面積體現(xiàn)了植物對(duì)營養(yǎng)資源的吸收能力[35]。本研究中細(xì)根的比根長、比表面積均以N2 處理下最佳,說明高濃度氮添加(N2)更有利于提高植物對(duì)養(yǎng)分的吸收效率,這與史順增等[17]的研究結(jié)果相似。稀土尾砂的氮含量較低,植物可利用的氮元素較少,氮添加使土壤可利用氮增加,提高植物的凈光合速率,從而提升植物的生產(chǎn)力,促進(jìn)植物的生長[36]。本研究結(jié)果顯示,與CK 相比,猴樟幼苗根長、根表面積在施氮條件下均有顯著增加(P<0.05),其中N1 處理的提升效果高于N2,表明低氮(N1)條件更適合猴樟幼苗根系生長。這與郝龍飛等[37]對(duì)樟子松(Pinus sylvestrisvar.mongolica)的研究結(jié)果相似,其研究結(jié)果表明,高濃度氮添加會(huì)使樟子松根長、根表面積等根系形態(tài)指標(biāo)下降。這可能是由于氮添加對(duì)植物生長的促進(jìn)作用是有一定范圍的,超過這個(gè)范圍,植物的生長就不再受到氮輸入的正面影響,甚至?xí)霈F(xiàn)抑制生長的現(xiàn)象[38]。
3.3 氮添加對(duì)猴樟幼苗NSC的影響
不同氮添加下植物各組織間的NSC 及其組分的分配格局體現(xiàn)了植物對(duì)其生活環(huán)境條件的適應(yīng)[39]。本研究中,猴樟幼苗各組織NSC 含量由高到低依次為:葉、粗根、莖、細(xì)根。這可能是由于葉是植物NSC 的主要合成部位[40],根是NSC 的主要儲(chǔ)存部位[41],植物完成NSC 合成后首先需要滿足葉的需求,其次多余的NSC 再通過莖運(yùn)輸至根儲(chǔ)存,同時(shí)一部分NSC 也會(huì)儲(chǔ)存于莖中,而細(xì)根是養(yǎng)分吸收部位,NSC 含量低,這與呂茹冰等[42]的研究結(jié)果相似。本研究還發(fā)現(xiàn),不同氮添加對(duì)猴樟幼苗葉、莖、粗根、細(xì)根等不同組織的可溶性糖、淀粉、NSC含量的影響不同,N1處理下葉中的可溶性糖高于CK,而N2 條件下卻顯著低于CK 與N1(P<0.05)。這可能是因?yàn)榈蜐舛鹊砑訒?huì)提高植物葉片的光合速率,促進(jìn)植物光合作用合成和積累有機(jī)物的能力[43],而氮濃度過高則會(huì)引起葉片的氣孔導(dǎo)度下降,減弱葉片的凈光合速率,使幼苗光合生產(chǎn)力受到抑制,導(dǎo)致植物積累有機(jī)物的能力下降[44]。葉、莖、細(xì)根中淀粉含量在施氮條件下均低于CK,不同施氮水平間差異不顯著(P>0.05),其中,莖、細(xì)根在N2處理下顯著低于CK(P<0.05),說明高氮條件下植物莖、細(xì)根中淀粉的積累受到抑制,導(dǎo)致根系生長受到影響,閆道良等[45]的研究也說明了這一點(diǎn)。而且,植物不同組織的NSC 含量對(duì)N 添加表現(xiàn)出不同的響應(yīng)特點(diǎn),相比于CK,施氮均降低了植物各組織NSC 含量,其中猴樟幼苗葉、莖、粗根、細(xì)根NSC 含量在N2 水平下差異最明顯,表明高氮條件不利于植物NSC的積累。
4 結(jié) 論
稀土尾砂中氮輸入對(duì)猴樟幼苗的生物量積累、根系生長與非結(jié)構(gòu)性碳水化合物含量具有重要影響。氮添加顯著增加了猴樟幼苗的生物量積累(P<0.05),其中N1 處理下猴樟幼苗通過增加根長和根表面積來加強(qiáng)根系構(gòu)建,使植株對(duì)養(yǎng)分的獲取能力增強(qiáng),葉、莖、根和冠層生物量積累增加;N2 處理下猴樟幼苗主要通過增加細(xì)根的比根長與比表面積提升植物對(duì)養(yǎng)分的吸收能力。綜合考慮植物生物量的積累和肥料的經(jīng)濟(jì)投入,稀土尾砂中添加N1 水平(即1.8 g·株-1)氮素可以滿足猴樟幼苗生長所需。
猜你喜歡
體育科技文獻(xiàn)通報(bào)(2022年3期)2022-05-23 13:46:54
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
小讀者(2021年2期)2021-03-29 05:03:48
科技傳播(2019年22期)2020-01-14 03:06:54
華人時(shí)刊(2019年13期)2019-11-17 14:59:54
民用飛機(jī)設(shè)計(jì)與研究(2019年4期)2019-05-21 07:21:24
文苑(2018年22期)2018-11-19 02:54:14
紅領(lǐng)巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
紅領(lǐng)巾·萌芽(2016年1期)2016-09-10 07:22:44
