肉源凝固酶陰性葡萄球菌對多環(huán)芳烴的消減作用
2024-01-03 05:41:22劉航航潘瓊羅慧婷蔡克周徐寶才李沛軍
食品研究與開發(fā) 2023年24期
劉航航,潘瓊,羅慧婷,蔡克周,徐寶才,李沛軍
(合肥工業(yè)大學 食品與生物工程學院,安徽 合肥 230009)
多環(huán)芳烴(polycyclic aromatic hydrocarbons,PAHs)是一類含有兩個或多個由碳原子和氫原子組成的強疏水性有機化合物,具有至少兩個凝聚或融合的芳香環(huán)結(jié)構(gòu)[1]。PAHs 源于有機物的不完全燃燒和熱解,可以通過食物鏈進入人體內(nèi)造成危害[2]。由于雙環(huán)氧化合物的代謝激活會導致DNA 復制和突變錯誤從而導致癌變,PAHs 被認為對人體具有潛在的遺傳毒性和致癌性,易使人體罹患乳腺癌、肺癌和結(jié)腸癌等癌癥[3]。國際癌癥研究機構(gòu)(International Agency for Research on Cancer,IARC)已經(jīng)將PAHs 分類為1 類致癌物、2A 類致癌物和2B 類致癌物[4],其中研究較多的是1 類致癌物苯并[a]芘(benzo [a] pyrene,BaP)和2B 類致癌物苯并[a]蒽(benzo [a] anthracene,BaA)、(chrysene,CHR)和苯并[b]熒蒽(benzo [b]fluoranthene,BbFA。歐盟委員會(European Commission,EC)規(guī)定了煙熏肉制品中BaP和PAH4(BaA、CHR、BbFA 和BaP)的最大限量,分別為2.0 μg/kg 和12.0 μg/kg[5]。我國GB 2762—2022《食品安全國家標準食品污染物限量》規(guī)定,肉及肉制品中BaP 的含量不得超過5 μg/kg[6]。為減少食品中PAHs危害物對人體健康的潛在危害,降低其含量具有重大意義。
目前,對食品中PAHs 的減少措施主要集中于物理和化學方法。比較常見的物理方法有改變煙熏方式和食品原料的預處理[7]等;在化學方法方面,通過向肉制品中添加醋和啤酒等物質(zhì),利用其中酚類物質(zhì)和抗氧化成分降低肉制品中PAHs[8]。然而,這些物理和化學方法大多昂貴、復雜和低效,甚至會產(chǎn)生有毒的次生代謝物,使這些方法往往難以推廣和大規(guī)模實踐[9]。與此同時,隨著消費者對良好生活質(zhì)量的要求不斷提高,消費者更愿意接受綠色安全的方法。
近年來,通過生物學方法降解PAHs 受到廣泛關(guān)注。許多細菌已被證實通過代謝或共代謝降解PAHs,并且PAHs 降解酶和生物降解途徑是多樣的。在環(huán)境領(lǐng)域,Hadibarata 等[10]研究表明白腐真菌(Armillaria sp.)F022 可以將BaP 降解為苯并[a]芘-1,6-醌,1-羥基-2-苯甲酸和苯甲酸。在食品領(lǐng)域,乳酸菌和凝固酶陰性葡萄球菌(coagulase-negative staphylococci,CNS)是傳統(tǒng)發(fā)酵肉制品中最主要的細菌群,它們決定了產(chǎn)品的感官和安全特性[11]。Bartkiene 等[12]研究了乳酸菌對冷熏豬肉香腸中PAHs 的降低效果,結(jié)果表明乳酸片球菌(Pediococcus acidilactici)KTU05-7、戊糖片球菌(Pediococcus pentosaceus)KTU05-9和清酒乳桿菌(Lactobacillus sakei)KTU05-6 均能夠降低熏腸內(nèi)部PAH4 含量,其中L.sakei KTU05-6 對PAH4 具有較好的去除效果。CNS 可以通過蛋白質(zhì)水解和脂肪分解來促進肉制品風味的形成,還可改善肉制品色澤[13]。然而,關(guān)于其消減食品中危害物的相關(guān)報道鮮見。
本研究通過分析肉源CNS 菌株在模擬體系中對PAHs 的降低作用,篩選出消減PAHs 能力最強的菌株,并通過明確菌株消減方式、消減位置和消減關(guān)鍵成分,探究其消減作用機理。
1 材料與方法
1.1 材料與試劑
PAHs 標準品(BaA、CHR、BbFA 和BaP)(純度>99%):北京北方偉業(yè)計量技術(shù)研究院;鏈霉蛋白酶(EC 3.4.24.31)、結(jié)晶紫、阿拉伯樹膠粉;上海源葉生物科技有限公司;酸水解酪蛋白、辛基苯基聚氧乙烯醚:北京索萊寶科技有限公司;三羥甲基氨基甲烷鹽酸鹽、布拉德福德蛋白濃度測定試劑盒:上海碧云天生物技術(shù)有限公司;磷酸二氫鈉(NaH2PO4)、磷酸氫二鈉(Na2HPO4)、乙腈、無水乙醇(均為色譜級):國藥集團化學試劑有限公司;除特殊標記外其他試劑均為分析純。
受試菌株:木糖葡萄球菌(Staphylococcus xylosus)A2、肉葡萄球菌(Staphylococcus carnosus)C2 均分離于自然發(fā)酵風干腸,馬胃葡萄球菌(Staphylococcus equorum)E1 分離于風鴨,小牛葡萄球菌(Staphylococcus vitulinus)CICC 10850:中國工業(yè)微生物菌種保藏管理中心。
7.5%氯化鈉肉湯(sodium chloride broth,SCB)培養(yǎng)基:廣東環(huán)凱微生物科技有限公司;甘露醇氯化鈉瓊脂(mannitol salt agar,MSA)培養(yǎng)基:廣東環(huán)凱微生物科技有限公司。
1.2 儀器與設備
潔凈工作臺(AlphaClean 1300):力康精密科技(上海)有限公司;振蕩培養(yǎng)箱(BSD-250):上海博迅實業(yè)有限公司;臺式低速離心機(TDZ5-WS):湖南湘儀實驗室儀器開發(fā)有限公司;酶標儀(SynergH1):美國Bio-Tek 儀器公司;液相色譜儀(S-6000):華譜科儀(北京)科技有限公司;水浴振蕩器(WE-3):天津歐諾儀器股份有限公司;分光光度計(UV-6000PC):上海元析儀器有限公司;靜音液晶超聲波清洗器(KS-7200XDS):昆山潔力美超聲儀器有限公司。
1.3 試驗方法
1.3.1 菌株培養(yǎng)
在無菌環(huán)境下,用無菌接種環(huán)挑取葡萄球菌斜面上的一環(huán)菌泥,將其接入10 mL 無菌SCB 培養(yǎng)基中,隨后漩渦振蕩,將接過菌的SCB 培養(yǎng)基置于恒溫培養(yǎng)箱,在恒溫條件(37 ℃,150 r/min)下培養(yǎng)18 h 進行活化。在無菌環(huán)境下,將活化后的SCB 培養(yǎng)基漩渦振蕩,通過無菌移液槍吸取0.2 mL 菌液接入10 mL 無菌SCB培養(yǎng)基中進行傳代,連續(xù)傳代培養(yǎng)2 次(37 ℃,18 h),第3 代菌液4 ℃冷藏備用。取10 mL 受試菌株第3 代菌液(1×108CFU/mL),離心(12 000×g,15 min,4 ℃)去上清液得全細胞。全細胞經(jīng)無菌磷酸鹽緩沖溶液(phosphate buffer solution,PBS,50 mmol/L,pH7.0)洗滌2 次,并溶解在10 mL PBS 中以獲得全細胞懸液(whole cell suspension,WCS)。
1.3.2 不同菌株對PAHs 消減能力的測定
不同菌株對PAHs 消減能力的評定參照Yousefi等[14]的方法并稍作修改。在PBS 添加10 ng/mL PAHs標準品,制備得到模擬體系。即向WCS 和PBS 中添加PAHs 標準品,使其最終濃度為10 ng/mL,其中PBS 作為對照組。培養(yǎng)(37 ℃,150 r/min,18 h)結(jié)束后檢測PAHs含量。
PAHs 含量測定參照Li 等[15]的方法并稍做修改,采用高效液相色譜法(high performance liquid chromatography,HPLC)進行。待測樣品經(jīng)0.22 μm 濾膜過濾后,將其添加到2 mL 進樣瓶中用于HPLC 檢測。色譜條件:在裝配熒光檢測器的高效液相色譜儀上進行分離采用雙流動相,A 相乙腈,B 相水。流動相洗脫梯度:0~17.5 min,40%~100%乙腈;17.5~24.0 min,100%乙腈;24.0~25.5 min,40%乙腈;25.5~27.5 min,40%乙腈。熒光檢測器的激發(fā)與發(fā)射波長為BaA:274/382 nm;CHR:260/360 nm;BbFA:283/430 nm;BaP:285/410 nm。
1.3.3 S.equorum 不同細胞組分消減PAHs 能力的測定
參照Xiao 等[16]的方法并稍作修改。S.equorum WCS經(jīng)超聲破碎(600 W,工作2 s,間歇3 s,180 次,0 ℃)得全細胞提取液(whole cell extracts,WCE),將全細胞提取液離心(12 000×g,15 min,4 ℃)得胞內(nèi)提取液(intracellular extracts,IE) 和細胞碎片。將細胞碎片溶解在10 mL PBS 中,獲得細胞碎片懸液(cell debris suspension,CDS)。分別向WCS、WCE、CDS、IE 和10 mL PBS中添加PAHs 標準品,使其最終濃度為10 ng/mL,培養(yǎng)(37 ℃,150 r/min,18 h)結(jié)束后檢測PAHs 含量。
1.3.4 蛋白酶處理對S.equorum WCE 消減PAHs 效果的影響
參照O'Brien 等[17]的方法并稍作修改。將0.1 mL鏈霉蛋白酶溶液添加到10 mL S.equorum WCE 中,以不添加鏈霉蛋白酶作為對照,水浴(37 ℃,1 h)冷卻后,添加PAHs 標準品,使其最終濃度為10 ng/mL,培養(yǎng)(37 ℃,150 r/min,18 h)結(jié)束后檢測PAHs 含量。
1.3.5 S.equorum 中PAHs 降解酶活力的測定取上述制備的S.equorum IE 樣品,用于鄰苯二酚2,3-雙加氧酶、脂肪酶和α-淀粉酶活力的測定。
1.3.5.1 鄰苯二酚2,3-雙加氧酶活力
參照Xi 等[18]的方法并稍做修改。酶活性反應體系:0.2 mL 鄰苯二酚(5 mmol/L),3 mL PBS(50 mmol/L,pH 8.0)和0.2 mL IE,測定其在3 min 內(nèi)、375 nm 處吸光度的變化。定義每分鐘1 μmoL 鄰苯二酚裂解產(chǎn)生2-羥粘糠酸半醛所需要的酶量為1 個酶活力單位U。鄰苯二酚2,3-雙加氧酶的酶活力按公式(1)計算。
式中:U 為酶活力,U/mL;A 為樣品吸光值;Vt為反應總體積,mL;ε 為摩爾吸光系數(shù);t 為反應時間,min;d 為比色皿內(nèi)腔厚度,cm;Vs為粗酶液體積,mL。
通過布拉德福德蛋白濃度測定試劑盒檢測蛋白濃度后,用公式(1)所得結(jié)果再除以每毫升粗酶液中所含蛋白含量,即可得酶的比活力。
1.3.5.2 脂肪酶活力
參照Wang 等[19]的方法并稍作修改。底物溶液的制備方法如下:取90 mg 對硝基苯酚棕櫚酸酯溶于30 mL乙腈后,將100 μL 對硝基苯酚棕櫚酸酯溶液加入到900 μL 含有0.1%阿拉伯樹膠和0.4% Triton X-100的50 mmol/L Tris-HCl 緩沖液中。在37 ℃和150 r/min條件下孵育5 min 后,向樣品中加入50 μL IE,并在150 r/min 和37 ℃條件下再孵育5 min。用300 μL 的10% Na2CO3終止反應。混合物經(jīng)離心后測量410 nm處的吸光度。定義在37 ℃條件下,每分鐘釋放1 μmol對硝基苯酚(p-nitrophenol)所需的酶量為1 個酶活力單位U。脂肪的酶活力按公式(2)計算。
式中:U 為酶活力,U/mL;A1為樣品吸光值;A0為樣品空白吸光值;K 為對硝基苯酚標準曲線的斜率;C0為對硝基苯酚標準曲線的截距;n 為稀釋倍數(shù);V1為反應液體積,mL;V2為粗酶液體積,mL;t 為反應時間,min。
通過布拉德福德蛋白濃度測定試劑盒檢測蛋白濃度后,用公式(2)所得結(jié)果再除以每毫升粗酶液中所含蛋白含量,即可得酶的比活力。
1.3.5.3 α-淀粉酶酶活力
參照Bo?ic等[20]的方法并稍作修改。采用3,5-二硝基水楊酸(3,5-dinitrosalicylic acid,DNS)法測定α-淀粉酶活力。將含有100 μL IE 和400 μL 1.0%可溶性淀粉的反應混合物在PBS(50 mmol/L,pH7.0)中孵育(40 ℃,30 min)。通過加入1 mL DNS 試劑停止反應,然后將混合物煮沸5 min,冷卻至室溫并用蒸餾水稀釋。淀粉水解過程中釋放的還原糖量通過葡萄糖標準曲線測定。定義在40 ℃、pH7 條件下,每分鐘淀粉水解產(chǎn)生1 mg 葡萄糖所需要的酶量為1 個酶活力單位U。α-淀粉酶的酶活力按公式(3)計算。
式中:U 為酶活力,U/mL;m 為葡萄糖質(zhì)量,mg;n為稀釋倍數(shù);V 為反應所用淀粉酶原液體積,mL;t 為反應時間,min。
通過布拉德福德蛋白濃度測定試劑盒檢測蛋白濃度后,用公式(3)所得結(jié)果再除以每毫升粗酶液中所含蛋白含量,即可得酶的比活力。
1.3.6 熱失活S.equorum 對PAHs 消減作用的影響
參考Yousefi 等[14]的方法對S.equorum 進行熱處理。WCS 經(jīng)熱處理(100 ℃,15 min)冷卻后,添加PAHs標準品,使其最終濃度為10 ng/mL,對照組不經(jīng)熱處理直接添加PAHs 標準品。培養(yǎng)(37 ℃,150 r/min,18 h)結(jié)束后通過離心(12 000×g,15 min,4 ℃)去除細胞,收集上清液用于檢測PAHs 含量。
1.3.7 S.equorum WCE 的傅里葉變換紅外光譜分析
參照Peng 等[21]的方法并稍做修改。S.equorum 的WCE 經(jīng)凍干機凍干后,采用傅里葉變換紅外光譜儀進行光譜掃描,掃描范圍為4 000~400 cm-1。
1.4 數(shù)據(jù)處理
試驗重復3 次,結(jié)果以平均值±標準差表示。使用SPSS 25.0 軟件對數(shù)據(jù)進行分析,各數(shù)據(jù)之間的顯著性差異(P<0.05)使用Duncan's 多重分析法進行檢驗,利用Origin 2017 作圖。
2 結(jié)果與分析
2.1 不同CNS 菌株對PAH4 的消減效果
食品中將包括BaP 在內(nèi)的16 種多環(huán)芳烴列為食品系統(tǒng)的主要污染物,歐洲食品安全委員會(EFSA)建議用PAH4(BaA、CHR、BbFA 和BaP)代表食品中多環(huán)芳烴的產(chǎn)生和毒性水平,因此本研究將PAH4 作為篩選菌株的依據(jù)。在模擬體系中接種不同菌株對PAHs的消減效果見圖1。
圖1 不同菌株對PAH4 降低量的影響Fig.1 Effects of different bacteria on PAH4 reduction
由圖1 可知,接種組中PAH4 含量均顯著低于對照組(P<0.05),即4 種葡萄球菌菌株均能顯著降低模擬體系中的PAH4(P<0.05)。S.xylosus A2、S.carnosus C2、S.equorum E1 和S.vitulinus CICC 10850 對BaA、CHR、BbFA 和BaP 的降低率分別為3.63%~12.14%、3.86%~8.69%、12.71%~17.11%和6.21%~15.12%。在所有待測菌株中,S.xylosus A2 和S.carnosus C2 降低PAH4 能力最弱,S.equorum E1 降低PAH4 能力最強,S.equorum E1 對BaA、CHR、BbFA 和BaP 的降低率分別達到15.79%、17.11%、13.54%和12.71%。不同的葡萄球菌菌株對PAHs 表現(xiàn)出不同的消減效果,這與Zhao等[22]的結(jié)果類似,他們的研究表明,15 株乳酸菌均可去除無菌蒸餾水中的BaP,去除率為11%~67%。這主要由菌株特異性造成。后續(xù)試驗以降低PAH4 能力最強的S.equorum E1 為研究對象,探究其對PAHs 的消減機理。
2.2 不同細胞組分對消減PAH4 的影響
為了明確S.equorum 各細胞組分對PAHs 的消減效果,比較了WCS、WCE、IE 和CDS 對PAHs 的消減效果。S.equorum 的WCS、WCE、CDS 和IE 對PAH4 降低率見圖2。
圖2 S.equorum 的WCS、WCE、CDS 和IE 對PAH4 降低率Fig.2 The reduction rate of PAH4 treated with the WCS,WCE,CDS and IE of S.equorum
如圖2 所示,WCS 組對BaA、CHR、BbFA 和BaP的降低率分別為15.79%、17.11%、13.54%和12.71%。WCE 組對BaA、CHR、BbFA 和BaP 的降低率分別為20.00%、19.98%、13.65%和14.00%,WCE 組對BaA 和CHR 的降低率顯著高于WCS 組(P<0.05),這表明超聲破碎處理提高了S.equorum 對PAHs 的降低效果,可能是超聲處理使細胞中更多的消減物質(zhì)暴露,PAHs更容易與消減物質(zhì)接觸作用的結(jié)果。Yousefi 等[14]研究發(fā)現(xiàn)超聲處理可以改變細胞壁結(jié)構(gòu)、增加其通透性,從而增加肽聚糖的可利用性,使細胞壁上產(chǎn)生新的結(jié)合位點。可見,S.equorum 對PAHs 的消減可能與物理吸附途徑同時存在。WCE 組對PAH4 的降低率和CDS組不存在顯著差異(P>0.05),但顯著高于IE 組(P<0.05),這表明細胞碎片和細胞內(nèi)物質(zhì)都具有消減作用,從IE組的消減效果可得,S.equorum 對PAHs 的消減可能存在生物降解途徑。與本研究結(jié)果類似,Zhao 等[22]研究表明植物乳桿菌(L.plantarum)CICC 22135 和戊糖乳桿菌(L.pentosus)CICC 23163 肽聚糖在去除BaP 中起重要作用,并且細胞結(jié)構(gòu)完整程度影響B(tài)aP 去除效率。
2.3 S.equorum 生物降解PAH4 關(guān)鍵消減成分的確定
在生物降解PAHs 時,細菌中有多種酶及其蛋白質(zhì)產(chǎn)物參與細菌對PAHs 的降解。為了確認S.equorum中蛋白質(zhì)類物質(zhì)(酶和蛋白質(zhì)產(chǎn)物)是否在消減PAHs中起關(guān)鍵作用,探究蛋白酶處理對S.equorum 消減PAHs 的影響。蛋白酶處理前后S.equorum 對PAH4 的消減效果如表1 所示。
表1 蛋白酶處理對S.equorum 消減PAH4 的影響Table 1 Effects of protease treatment on PAH4-decreasing ability of S.equorum
對照組(未經(jīng)蛋白酶處理)對BaA、CHR、BbFA 和BaP 的降低率分別為20.00% 、19.98% 、13.65% 和14.00%,蛋白酶處理組對BaA、CHR、BbFA 和BaP 的降低率分別為11.53%、13.77%、7.55%和1.61%。經(jīng)蛋白酶處理后,S.equorum 對PAH4 消減能力顯著下降(P<0.05),由此表明S.equorum 細胞片段上蛋白質(zhì)類物質(zhì)(酶和蛋白質(zhì)產(chǎn)物)在消減PAHs 過程中起重要作用。Xiao 等[16]在探究戊糖乳桿菌R3 對N-亞硝胺降解時,發(fā)現(xiàn)其表層蛋白能顯著降低N-亞硝胺含量(P<0.05)。
2.4 S.equorum 生物降解PAH4 關(guān)鍵酶的探究
生物降解是在高度污染環(huán)境中去除PAHs 的主要途徑,Chen 等[23]研究表明枯草芽孢桿菌(Bacillus subtilis)和伯克霍爾德菌(Burkholderia cepacia)降低PAHs能力主要取決于它們是否具有參與芘降解的關(guān)鍵酶。已有研究表明,鄰苯二酚2,3-雙加氧酶(C23O)、脂肪酶和α-淀粉酶都與PAHs 和烴類化合物的代謝有關(guān)[23-25]。C23O 是超二醇雙加氧酶家族的成員,催化鄰苯二酚和取代鄰苯二酚環(huán)的裂解效應。Karimi 等[24]研究發(fā)現(xiàn)細菌產(chǎn)生的淀粉酶和商業(yè)淀粉酶均能降解烴類化合物。脂肪酶在油性烴類的生物降解中也起重要作用[25]。因此,通過檢測S.equorum 中關(guān)鍵酶(C23O、脂肪酶和α-淀粉酶)的活性可以初步判斷PAHs 降解代謝途徑。
在S.equorum 中檢測到3 種酶活性如圖3 所示。
圖3 S.equorum 中C23O、脂肪酶和α-淀粉酶的比酶活Fig.3 Specific activity of C23O,Lipase and α-amylase in S.equorum
由圖3 可知,C23O、脂肪酶和α-淀粉酶的比酶活分別為56.65、266.92、3.31 U/mg,其中脂肪酶活性最高(P<0.05),這表明S.equorum 可能通過產(chǎn)生這些關(guān)鍵降解酶來降解PAHs。
2.5 熱失活S.equorum 對PAH4 的物理吸附作用
為了明確S.equorum 對PAHs 的去除是否存在物理吸附途徑,研究了細胞活性對消減PAHs 的影響,結(jié)果見表2。

表2 細胞活性對S.equorum 消減PAH4 的影響Table 2 Effect of cell viability on PAH4-decreasing ability of S.equorum
由表2 可以看出,對照組對BaA、CHR、BbFA 和BaP的降低率分別為15.79%、17.11%、13.54%和12.71%,熱處理組對BaA、CHR、BbFA 和BaP 的降低率分別為28.77%、30.52%、25.41%和27.38%,S.equorum 經(jīng)熱處理后對PAH4 的降低能力顯著提升(P<0.05),這與Yousefi 等[14]的結(jié)果一致。Zhu 等[26]研究指出熱處理提高了乳酸菌對酚甲烷的結(jié)合去除能力。結(jié)合本研究結(jié)果可知,S.equorum 在熱處理后可能改變了細菌細胞壁結(jié)構(gòu),從而產(chǎn)生新的結(jié)合位點,增加了細胞對多環(huán)芳烴的吸附。Zhao 等[22]認為乳酸菌對BaP 的去除不存在生物代謝,在PAHs 去除過程中細胞活性是不必要的。
2.6 S.equorum 對PAH4 的吸附官能團分析
通過FTIR 分析S.equorum 的WCE 以識別與PAHs結(jié)合相關(guān)的潛在官能團和可能的結(jié)合位點,獲取PAHs 消減過程中涉及官能團的更多信息結(jié)果見圖4。
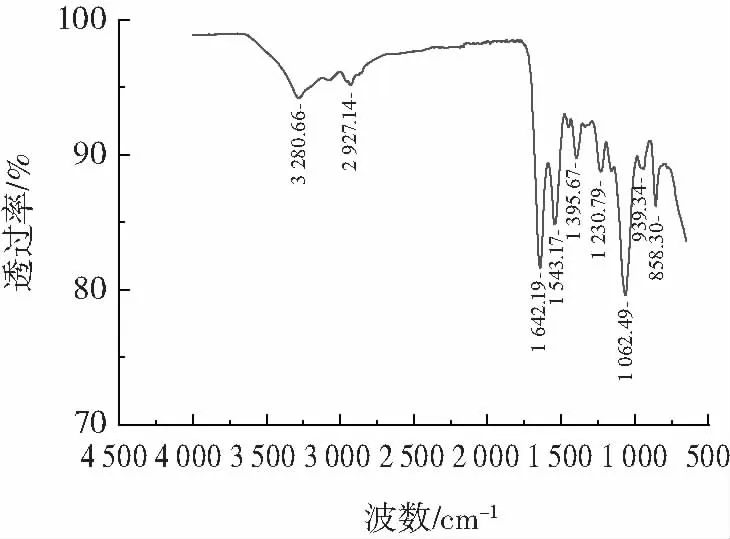
圖4 S.equorum 全細胞提取液的傅里葉紅外分析Fig.4 Fourier transform infrared spectrum of the whole cell extracts of S.equorum
如圖4 所示,根據(jù)文獻[27]報道對L.pentosus R3主要吸收譜帶進行歸屬:3 280.66 cm-1(—OH 的伸展振動吸收);2 927.14 cm-1(C—H 鍵的不對稱伸縮振動吸收);1 642.19 cm-1(酰胺Ⅰ帶中的C O 伸縮振動吸收);1 543.17 cm-1(酰胺Ⅱ帶中的N—H 鍵的彎曲振動吸收和C—N 鍵的拉伸);1 395.67 cm-1(羧基的對稱伸縮振動);1 230.79 cm-1(脂質(zhì)和多糖P O 的伸縮振動);1 062.49 cm-1(肽聚糖C—O 的伸縮振動)。939.34 cm-1(羧酸面外彎曲振動吸收);858.30 cm-1(C—H 面外彎曲振動吸收)。Xiao 等[28]表明官能團(羥基、胺基等)除了引發(fā)范德華力,還可以引發(fā)各種分子水平的相互作用,這些基團在白腐真菌吸附PAHs 過程中也發(fā)揮重要作用。此外,這些化學基團存在于多糖、蛋白質(zhì)和核酸等物質(zhì)中,一些脂肪族和芳香族成分可能有助于S.equorum 對PAHs 物理吸附。
3 結(jié)論
在模擬體系中,受試4 株CNS 菌株對PAHs 都具有消減作用,其中S.equorum 效果最好。進一步分析發(fā)現(xiàn),S.equorum 的WCS、WCE、IE 和CDS 對PAHs 均具有消減效果,且WCE 和CDS 對PAHs 消減效果最強(P<0.05)。蛋白酶處理使S.equorum WCE 對PAH4 消減能力顯著降低(P<0.05),說明S.equorum 關(guān)鍵消減物質(zhì)為蛋白類物質(zhì);同時,在WCE 中檢測到PAHs 關(guān)鍵降解酶活性,表明S.equorum 消減PAHs 存在生物降解途徑。熱滅活處理使S.equorum 對PAH4 消減能力顯著提升(P<0.05),說明S.equorum 消減PAHs 存在物理吸附途徑,并且—OH、C O、N—H、C—N 等官能團參與了物理吸附過程。因此,S.equorum 可以通過物理吸附和生物降解兩種作用途徑消減PAHs。
