原兒茶酸與卵白蛋白的相互作用
2024-01-03 05:45:16王晨雨付子航陳成誠陳姝彤張平平甘宇鑫張軍兵張愛琳江慎華
食品研究與開發 2023年24期
王晨雨,付子航,陳成誠,陳姝彤,張平平,甘宇鑫,張軍兵,3,張愛琳,江慎華,3*
(1.天津農學院 食品科學與生物工程學院,天津 300392;2.九江學院藥學與生命科學學院,江西 九江 332000;3.江西丹霞生物科技股份有限公司,江西 鷹潭 335000)
原兒茶酸(protocatechuic acid,PCA)是一種天然酚酸,廣泛存在于日常飲食和中草藥中(如姜黃)[1],也是人體腸道微生物群降解多酚(特別是花青素或原花青素等類黃酮類)的代謝產物[2],具有抗氧化[3]、抗過敏[4]、神經保護[5]和抗骨質疏松[6]等多種活性。
卵白蛋白(ovalbumin,OVA)是一種磷糖球蛋白,分子量為42~47 kDa,是蛋清蛋白的主要成分,由385個氨基酸組成,50%氨基酸疏水、33%氨基酸帶電,顯示出其作為親脂成分高效載體的潛力[7]。OVA 作為一種載體,已被廣泛用于研究小分子與OVA 之間的結合機制,包括結合方式和作用力。已有研究結果表明,OVA能夠有效改善小分子水溶性、穩定性和生物利用度[8]。
多酚與蛋白質的相互作用可能會使蛋白質的相關特性發生改變。Seczyk 等[9]指出,酚類化合物與蛋白質的不可逆結合可能會影響蛋白質的消化吸收及氨基酸的生物利用度。同時多酚與蛋白質發生相互作用,可能會對食品營養和功能特性產生影響。由此可見,生物活性成分與載體絡合具有較多優勢,會發生很多變化,了解兩者之間的相互作用和結合機制至關重要[10]。龍梅等[11]研究發現,PCA 能夠猝滅魚精DNA的本征熒光,且猝滅機理為靜態猝滅。但是,PCA 與OVA 相互作用及其作用機制目前尚不清楚。
綜上,本文采用多光譜技術對PCA 與OVA 的相互作用及其機制進行研究,并測定相互作用前后對ABTS+自由基清除能力的影響,以期為拓寬PCA 和OVA 在食品工業中的應用提供參考。
1 材料與方法
1.1 材料與試劑
PCA(純度≥97%)、OVA 凍干粉(純度>98%)、2,2-聯氮-二(3-乙基-苯并噻唑-6-磺酸)二銨鹽[2,2'-azinobis-(3-ethylbenzthiazoline-6-sulphonate,ABTS]、磷酸氫二鈉、磷酸二氫鉀(均為分析純):上海阿拉丁生化科技股份有限公司。
1.2 儀器與設備
熒光分光光度計(F-7000):日本日立公司;酶標儀(SpectraMax 190):美谷分子儀器(上海)有限公司;pH計(PB-10):Sartorius 儀器設備有限公司;雙光束紫外-可見分光光度計(TU-1901):北京普析通用儀器有限責任公司。
1.3 試驗方法
1.3.1 溶液配制
OVA 采用0.05 mol/L、pH7.4 磷酸鹽緩沖液(phosphate-buffered saline,PBS)溶解、得到1×10-5mol/L 儲備液,PCA 采用去離子水溶解。
1.3.2 熒光光譜測定
參照Wang 等[12]方法,稍加修改。取3.0 mL 濃度為1×10-5mol/L 的OVA 溶液于熒光比色皿中,連續滴加10 μL 濃度1×10-2mol/L 的PCA 溶液、充分混勻,分別于284、291、298 K 條件下水浴靜置5 min 使其平衡。熒光分光光度計狹縫寬度設置為5 nm,激發波長(λex)分別設置為280 nm[色氨酸(Trp)和酪氨酸(Tyr)均被激發]和295 nm(僅Trp 被激發),發射光譜(λem)在300~450 nm 處以1 200 nm/min 的掃描速率讀取。在284、291、298 K 記錄OVA 的熒光發射光譜,以評價溫度對PCA 與OVA 相互作用的影響。
1.3.3 紫外-可見光譜測定
于298 K 條件下掃描樣液在190~500 nm 的紫外-可見光譜,以PBS 緩沖液作為空白對照。
1.3.4 同步熒光光譜測定
同時改變激發和發射單色器獲得284 K 同步熒光光譜,狹縫寬度均為5 nm。激發和發射波長的差值(△λ=λem-λex) 分別固定在15 nm 和60 nm。當△λ=15 nm 時(λem=275 nm,λex=260~320 nm) 僅顯示Tyr殘基的特征光譜;當△λ=60 nm 時(λem=310 nm,λex=250~320 nm)僅顯示Trp 殘基的特征光譜[13]。所有樣品恒溫孵育5 min 后進行測量。
1.3.5 三維熒光光譜測定
狹縫寬度為5 nm,掃描速度60 000 nm/min,測定激發波長200~400 nm、發射波長200~600 nm 的三維熒光光譜,增量為5 nm。
1.3.6 ABTS+自由基清除能力測定
采用Moghadam 等[14]方法稍作修改,測定樣品對ABTS+自由基清除活性。采用5 mmol/L 磷酸緩沖液(pH7.4)溶解得到ABTS 原液,將7 mmol/L ABTS 原液與2.45 mmol/L 過硫酸鉀等體積混合,室溫避光靜置12~16 h,使其生成ABTS+自由基。再將ABTS+溶液在734 nm 波長監測下稀釋至吸光值為0.70±0.02。分別將50 μL 濃度0.012 5~0.400 0 μg/mL 的樣品溶液與100 μL 稀釋后的ABTS+溶液混合,室溫避光反應10 min,測734 nm 處吸光值。用磷酸緩沖液代替樣品為對照。采用以下公式計算樣品對ABTS+自由基清除活性。
式中:X 為ABTS+自由基清除率,%;A0和A 分別為對照和樣品的吸光值。
1.4 數據處理
每組試驗均獨立重復3 次,所有數據用平均值±標準差表示,采用單因素方差分析與最小顯著性差異法檢驗比較各組差異,p<0.05 為具有顯著性差異。
2 結果與分析
2.1 PCA 對OVA 熒光猝滅效果的測定
對于大分子,熒光測量可以在分子水平上給出小分子物質與蛋白質大分子結合的信息,如結合常數、結合位點數、熱力學參數、相互作用力和作用機理等[12]。熒光發射光譜可用來獲得OVA 三級結構構象變化的信息,氨基酸殘基是主要的熒光團[15]。OVA 的熒光來源于Trp、Tyr 和苯丙氨酸(Phe)殘基。同時,OVA 的本征熒光特性對其微環境非常敏感,一些因素(如蛋白質構象轉變、亞單位結合、底物結合和變性)會導致蛋白質的本征熒光變化。因此,蛋白質的本征熒光可以提供關于其結構和動力學的大量信息[16]。圖1 為在pH7.4 條件下,加入PCA 的OVA 在λex=280 nm 處的熒光發射光譜。
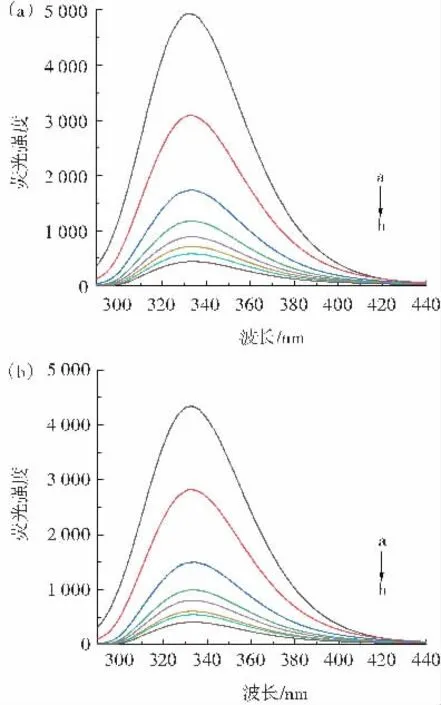

圖1 不同溫度下PCA 與OVA 相互作用熒光光譜Fig.1 Fluorescence spectra of interaction between PCA and OVA at different temperatures
由圖1 可知,在不同溫度下,OVA 的熒光強度隨PCA 濃度增加逐漸降低,最大發射波長沒有明顯變化,表明PCA 與OVA 之間存在相互作用,PCA 與OVA 的結合對熒光團周圍的微環境沒有太大影響。Li 等[17]對原花青素與OVA 相互作用的研究結果與本文結論一致。
2.2 PCA 對OVA 熒光猝滅機理分析
熒光猝滅表示大分子與猝滅劑分子的各種相互作用引起熒光量子產率的降低,即熒光強度的降低。OVA 的本征熒光對其微環境非常敏感,當OVA 周圍微環境稍有改變時,其本征熒光會明顯減弱,蛋白質構象轉變、生物分子結合與變性等因素是導致熒光強度減弱的原因[18]。該過程可由多種機制引起,通常包括動態猝滅和靜態猝滅[10]。溫度效應會導致靜態猝滅不同于動態猝滅。在動態猝滅過程中,溫度越高、擴散系數越大,熒光團和猝滅劑之間的碰撞次數越多,猝滅常數隨著溫度的升高而增大;而在靜態猝滅過程中,熒光團和猝滅劑之間形成基態絡合物,猝滅常數隨溫度增加而減小[19]。使用Stern-Volmer 方程分析不同溫度下的熒光數據可闡明熒光猝滅機理[10],公式如下。
F0/F=1+Ksv[Q]=1+Kqτ0[Q]
式中:F0和F 分別為不存在和存在PCA 時OVA 的穩態熒光強度;Ksv 為Stern-Volmer 猝滅常數,L/mol;[Q] 為PCA 濃度,mol/L;Kq 為雙分子擴散猝滅速率常數,2.0×10-10L/(mol·s),τ0為沒有與PCA 結合時熒光團的平均壽命,10-8s。
在2.1 中發現PCA 對OVA 熒光會產生猝滅效果。采用Stern-Volmer 方程對猝滅作用機理進行分析。圖2 為在不同溫度(284、291、298 K) 下的Stern-Volmer線性圖,相關參數如表1 所示。

表1 不同溫度下PCA 對OVA 熒光猝滅的Stern-Volmer 猝滅常數Table 1 Stern-Volmer quenching constants of OVA by PCA at different temperatures
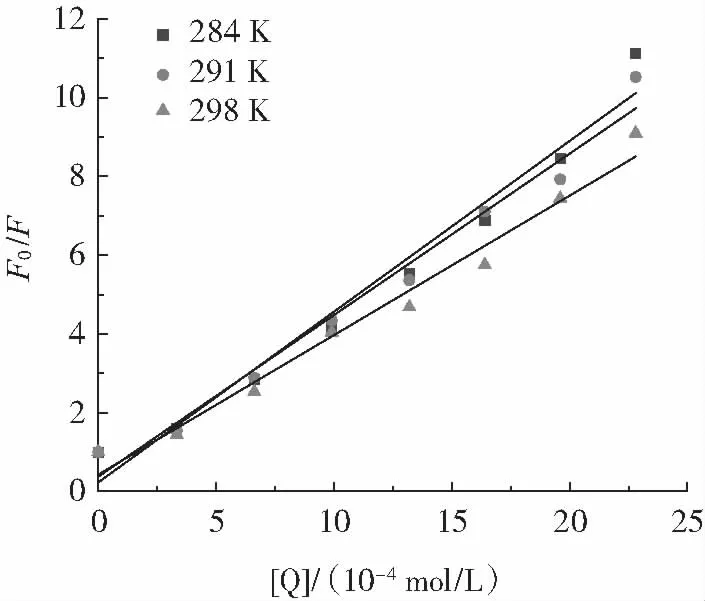
圖2 不同溫度下PCA 對OVA 熒光猝滅的Stern-Volmer 曲線Fig.2 Stern-Volmer curve for the fluorescence quenching of OVA by PCA at different temperatures
Ksv 反映熒光團對猝滅劑(PCA)的敏感性,有助于確定PCA-OVA 體系中的猝滅機制。由圖2 和表1可以發現,在284~298 K 內,Ksv 值隨溫度升高而降低,表明PCA 與OVA 的相互作用為靜態猝滅機制。Kq是衡量相互作用猝滅能力和效率的關鍵參數[20]。3 種溫度下計算得到的雙分子擴散猝滅速率常數[4.332×1011、4.098×1011、3.540×1011L/(mol·s)]均遠大于最大擴散碰撞猝滅速率常數[2.0×1010L/(mol·s)],進一步說明PCA 與OVA 相互作用機理為靜態猝滅。此外,Kq 值隨著溫度升高而降低,表明PCA 對OVA 的猝滅過程是一個放熱反應。
2.3 結合常數及結合位點數測定
在靜態猝滅機制下,結合常數(Ka)和結合位點數(n)可以使用雙對數方程計算[21],公式如下。
log[(F0-F)/F]=logKa+nlog[Q]
式中:Ka 為表觀結合常數,L/mol,表示OVA 與PCA 結合的親和力;n 為結合位點數。
在2.2 中確定猝滅機制后,對PCA 與OVA 相互作用的表觀結合常數(Ka)和結合位點數(n)進行測定。圖3 為不同溫度下PCA 與OVA 相互作用的雙對數曲線,結合常數及結合位點數如表2 所示。

表2 不同溫度下PCA 與OVA 相互作用的結合常數Table 2 Binding constants of interaction between PCA and OVA at different temperatures

圖3 不同溫度下PCA 與OVA 相互作用的雙對數曲線Fig.3 Double logarithmic curve of interaction between PCA and OVA at different temperatures
由表2 可知,Ka 值均大于1×104L/mol,表明PCA與OVA 具有較強的結合能力。Ka 值隨溫度升高而升高,表明PCA 與OVA 的絡合穩定性與溫度成正比。此外,在284~298 K 溫度范圍內,n 值均接近于1,顯示PCA與OVA 絡合物的化學計量比為1∶1,表明不同溫度下PCA 與OVA 只有一個結合位點。
2.4 熱力學參數及相互作用力判斷
通常,反應過程中焓變(ΔH)和熵變(ΔS)是確定小分子與蛋白質結合方式的主要熱力學參數,由van’t Hoff 方程(Eq)計算得到[22]。
ln Ka=-ΔH/RT+ΔS/R
ΔG=ΔH-TΔS
式中:T 為熱力學溫度,K;R 為氣體常數,8.314 J/(mol·K);ΔG 為吉布斯自由能變,kJ/mol;ΔH 為焓變,kJ/mol;ΔS 為熵變,J/(mol·K)。
通過對熱力學參數的研究,可以驗證作用過程的自發性,并探討蛋白質與配位體相互作用的機理。判斷主要結合驅動力的熱力學定律包括:1)ΔH>0、ΔS>0 或ΔH>0、ΔS<0 時,表示疏水相互作用為主要作用力;2)ΔH<0、ΔS<0 時,范德華力和氫鍵占主導;3)ΔH<0、ΔS>0時,主要作用力為靜電相互作用[23]。在2.3 計算出不同溫度下結合常數Ka 后確定的熱力學參數如表3 所示。

表3 不同溫度下PCA 與OVA 相互作用的熱力學參數Table 3 Thermodynamic parameters of interaction between PCA and OVA at different temperatures
由表3 可知,ΔG 為負值,表明PCA 與OVA 的結合是自發過程;ΔH 和ΔS 均大于0,表明自發過程是由熵驅動的吸熱反應,主導作用力是疏水相互作用。Ge 等[24]研究發現,疏水相互作用也是姜黃素及其衍生物與牛血清白蛋白相互作用的主要驅動力。
2.5 同步熒光光譜檢測PCA 對OVA 微環境變化
同步熒光光譜靈敏度高、選擇性好、干擾少,可以提供關于配體影響熒光團周圍微環境變化的重要信息[25]。因此,在2.1~2.4 基本確定PCA 與OVA 相互作用后,進一步通過同步熒光光譜檢測PCA 對OVA 構象變化的影響,結果如圖4 所示。
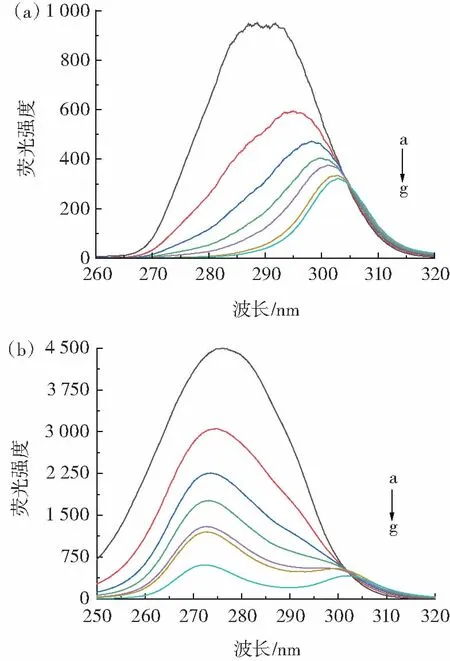
圖4 284 K 條件下PCA 與OVA 相互作用的同步熒光光譜Fig.4 Synchronous fluorescence spectra of interaction between PCA and OVA at 284 K
配體引起的蛋白質熒光猝滅意味著蛋白質氨基酸殘基周圍的極性改變[26]。由圖4 可知,隨著混合體系中PCA 濃度逐漸增加,兩個波長間隔下的熒光強度均呈現規律性下降,同時觀察到Tyr 殘基最大發射波長發生紅移(從288.6 nm 到303.0 nm),而Trp 殘基的最大發射波長沒有發生明顯變化,表明PCA 與OVA 相互作用使Tyr 殘基周圍微環境極性增加、疏水性降低,而Trp殘基所處微環境幾乎沒有發生變化。此外,Δλ=60 nm時的猝滅間距比Δλ=15 nm 處的猝滅間距更明顯,意味著結合位置更接近Trp 殘基[27]。
2.6 三維熒光光譜檢測OVA 微環境變化
三維熒光光譜可以同時改變激發和發射波長,進而提供更科學、更可靠的蛋白質結構和微環境變化的細節,能夠證明蛋白質在與配體結合時構象發生了變化[25]。圖5 為PCA 不存在和存在時OVA 的三維熒光光譜,相關特征參數如表4 所示。

表4 PCA 不存在和存在下OVA 的三維熒光光譜特征參數Table 4 Characteristic parameters in three-dimensional fluorescence spectra of OVA in the absence and presence of PCA

圖5 284 K 條件下PCA 與OVA 相互作用的三維熒光光譜Fig.5 Three-dimensional fluorescence spectra of interaction between PCA and OVA at 284 K
由圖5 可知,主要有4 個特征峰,峰A 和峰B 分別表示利散射峰和二階瑞利散射峰,對應波長分別為λex=λem 和2λex=λem,其特征是化合物溶解的溶劑進行輻射再發射,并且一小部分吸收的輻射在同一波長向各個方向散射[25]。峰1 提供了由于π-π* 躍遷而與二級結構相關的多肽骨架結構的光譜行為信息,峰2(λex=280.0 nm)反映的主要是Trp 和Tyr 殘基的本征熒光[28]。與OVA 相比,加入PCA 后峰1(λex 235.0 nm/λem 330.0 nm)的熒光強度從1 273.0(圖5a)明顯下降至181.7(圖5b),峰2 的熒光強度從4406.0(圖5a)明顯下降至1 748.0(圖5b),且從280.0 nm 明顯紅移至325.0 nm。結果表明PCA 與OVA 之間存在相互作用,且蛋白質的二級結構可能發生了變化、氨基酸殘基所處微環境的極性增加,該結果與同步熒光結果一致。
2.7 紫外-可見光譜檢測PCA 與OVA 相互作用
通過檢查熒光團的吸收光譜不僅可驗證猝滅類型,還可用于研究蛋白質發色團周圍微環境的變化和蛋白質配體復合體的形成。動態猝滅只影響熒光團的激發態,因此吸收光譜不會發生改變。然而,由于基態絡合物的形成,靜態猝滅經常導致熒光團吸收光譜的擾動[29]。為了進一步驗證PCA 與OVA 相互作用的機理及對蛋白質產生的影響,在298 K 條件下測定OVA及PCA-OVA 復合物的紫外-可見光譜,結果如圖6所示。

圖6 298 K 條件下PCA 與OVA 相互作用的紫外-可見光譜Fig.6 Ultraviolet-visible spectrum of interaction between PCA and OVA at 298 K
由圖6 可知,280 nm 附近顯示出吸收峰是由于蛋白質的生色團(芳香族氨基酸)對紫外線的吸收,常被用來反映蛋白質構象的變化[30]。隨著PCA-OVA 體系中PCA 濃度的增加,280 nm 處吸收峰略有增加,且在相互作用后發生了輕微紅移(從280 nm 到287 nm),表明PCA 和OVA 之間存在相互作用,并形成了基態復合物,再次證明PCA 對OVA 的熒光猝滅機制是靜態猝滅;同時表明PCA 與OVA 的相互作用影響了蛋白質構象,并改變了OVA 芳香族氨基酸殘基周圍的微環境,與同步熒光及三維熒光光譜結論一致。
2.8 PCA 與OVA 及其復合物對ABTS+自由基清除能力的影響
ABTS+自由基清除法是評價抗氧化能力最常見方法之一,本研究采用該方法對PCA、OVA 及PCA-OVA 復合物對ABTS+自由基清除能力進行測定,結果見圖7。
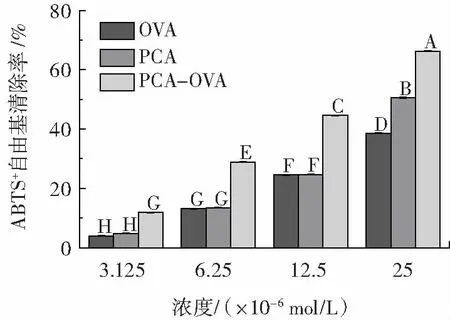
圖7 PCA、OVA 及PCA-OVA 復合物對ABTS+自由基的清除能力Fig.7 Scavenging ability of PCA,OVA and PCA-OVA complexes against ABTS+ radical
由圖7 可知,游離PCA、OVA 和PCA-OVA 復合物均表現出ABTS+自由基清除活性,且清除能力均隨濃度增加而增強。在相同濃度下,PCA-OVA 復合物的抗氧化活性較PCA 有顯著提高(p<0.05)。Liu 等[8]在研究姜黃素與OVA 結合前后清除DPPH 自由基活性變化時也發現了類似的研究結果。
3 結論
本文運用多光譜技術對PCA 與OVA 相互作用的機理進行研究,并通過ABTS+自由基清除試驗考察PCA 與OVA 相互作用對PCA 抗氧化性能的影響。結果顯示,在284~298 K 溫度范圍內,PCA 與OVA 具有一個較強親和力的結合位點;PCA 與OVA 的結合過程通過疏水作用力自發進行,并使Tyr 殘基周圍微環境極性增加,疏水性降低。PCA 與OVA 結合改變了對ABTS+自由基的清除能力。本研究為拓寬PCA 和OVA在食品工業中的應用提供依據。
