Gαi1/3蛋白介導Akt-GSK3β-NFATc1信號通路調控破骨細胞分化機制研究
2024-01-04 02:43:54史冊高運楊光輝
實用骨科雜志 2023年12期
史冊,高運,楊光輝*
(1.徐州醫科大學附屬宿遷醫院,南京鼓樓醫院集團宿遷醫院骨科,江蘇 宿遷 223800;2.揚州大學附屬醫院骨科,江蘇 揚州 225100)
骨質疏松癥(osteoporosis,OP)是以骨量降低、骨質脆性增加而易于骨折為特征的常見骨骼代謝性疾病,對中國居民的健康產生了嚴重的威脅[1]。既往研究表明,破骨細胞的數量異常增多以及功能異常活化可能是導致骨質疏松癥的直接致病原因[2]。破骨細胞作為體內唯一行使骨吸收功能的細胞,由骨髓源性巨噬細胞(bone marrow-derived macrophages,BMMs)表面集落刺激因子-1受體(colony stimulating factor 1 receptor,CSF-1R)及核因子κB受體活化因子配體(receptor activator for nuclear factor-κB ligand,RANKL)受體分別與巨噬細胞集落刺激因子(macrophage colony stimulating factor,MCSF)及RANKL結合啟動下游信號通路,最終分化而成[3]。MCSF屬于受體酪氨酸激酶(receptor tyrosine kinases,RTKs),能促使BMMs轉化為破骨細胞前體細胞并促進其增殖,同時可誘導破骨細胞前體細胞表達RANK受體增加,進一步促進破骨細胞的分化成熟[4]。近期研究發現G蛋白抑制性α亞單位1/3(Gαi1/3蛋白)在腦源性神經營養因子(brain derived neurotrophic factor,BDNF)、血管內皮生長因子(vascular endothelial growth factor,VEGF)、干細胞因子(stem cellfactor,SCF)等RTKs介導的信號傳導中發揮重要的作用[5-7],而其在同為RTKs的MCSF介導的信號傳導過程中的作用尚不清楚。本研究著重探討Gαi1/3蛋白在調控破骨細胞生成的信號傳導過程中發揮的作用,為防治骨質疏松癥提供新的思路。
1 材料和方法
1.1 實驗動物 本實驗所用8~10周齡野生型(wild type,WT)雌性C57BL/6小鼠由上海斯萊克實驗動物有限公司提供[實驗動物生產許可證號(SCXK(滬)2017-0005];Gαi1/3雙敲除(Gαi1/3-double knock out,Gαi1/3-DKO)C57BL/6雌性小鼠由吉凱基因技術有限公司(上海)構建完成。
1.2 主要試劑與儀器 主要試劑:α-MEM培養基(Corning公司,美國);胎牛血清、0.25%胰蛋白酶(Gibco公司,美國);二甲亞砜(Abcam,中國);重組小鼠MCSF蛋白、RANKL蛋白(Abcam公司,英國);scr-shRNA、Gαi1-shRNA、Gαi3-shRNA(吉瑪制藥技術有限公司,中國);Gαi1、Gαi3一抗(Santa Cruz公司,美國);p-Akt473、Akt1/2/3、p-GSK3β、GSK3β、NFATc1一抗(Cell Signaling Technology公司,中國);抗酒石酸酸性磷酸酶染色試劑盒(tartrate resistant acid phosphatase,TRAP)(Sigma公司,英國)。
主要儀器:酶標儀(TECAN公司,瑞士);倒置顯微鏡(NIKON公司,日本),Micro CT掃描機(通用公司,美國);低溫高速離心機(Thermo Fisher Scientific公司,美國)。
1.3 實驗方法
1.3.1 小鼠BMMs提取及純化 將8~10周齡野生型及Gαi1/3-DKO雌性C57/BL6小鼠斷頸處死,切取下肢并剔除所附軟組織,剪除股骨及脛骨兩端軟骨沖取髓腔內細胞,沖洗液經70 μm濾網過濾后1 200 rpm離心5 min,吸棄上清液后加入2 mL紅細胞裂解液吹打重懸后靜置裂解2 min,加入4 mL磷酸鹽緩沖液(phosphate buffered solution,PBS)終止裂解予1 200 rpm離心5 min,棄上清即得小鼠BMMs,加入完全培養基培養24 h后吸取上清液,同法離心棄上清,加入預制培養基[α-MEM+10%胎牛血清(fetal bovine serum,FBS)+50 ng/mL MCSF],培養2~3 d所得貼壁細胞即為純化BMMs。
1.3.2 慢病毒轉染 將所得純化BMMs以5×104/孔密度鋪板于6孔培養板中,待24 h細胞數量增殖約為1×105加入依據感染復數(multiplicity of infection,MOI)=10配置的含有scr-shRNA、Gαi1-shRNA和Gαi3-shRNA的無雙抗培養基(α-MEM+10%FBS),24 h后更換為無病毒培養基培養,定期于熒光顯微鏡下觀察轉染情況。
1.3.3 破骨細胞誘導分化及TRAP染色 以5×104/mL密度于6孔培養板中接種先前成功轉染慢病毒BMMs,加入濃度50 ng/mL MCSF+50 ng/mL RANKL進行誘導,每2 d更換培養液至第4天破骨細胞分化成熟。依據TRAP操作說明書進行染色,使用倒置顯微鏡進行樣本拍照,觀察破骨細胞大小及數量。
1.3.4 蛋白免疫印跡(western blot,WB) 待檢測預處理BMMs及分化成熟破骨細胞,使用RIPA裂解液提取蛋白。使用十二烷基硫酸鈉聚丙烯酰胺凝膠電泳(sodium dodecyl sulfate polyacrylamide gel electrophoresis,SDS-PAGE)分離總蛋白,再轉印至聚偏二氟乙烯(polyvinylidene difluoride,PVDF)上,脫脂牛奶封閉1 h,4 ℃孵育一抗Gαi1、Gαi3、p-Akt473、Akt1/2/3、p-GSK3β、GSK3β、NFATc1搖床過夜,隨后二抗孵育2 h,完成后使用PBS加Tween20(PBS plus tween 20,PBST)清洗后加入電化學發光(electro-chemi luminescence,ECL)壓片顯影,使用Image J軟件定量分析目標條帶。
1.3.5 小鼠骨質疏松模型制備及給藥 隨機選擇12只6~7周齡雌性C57/BL6小鼠分為scr-shRNA及Gαi1/3-shRNA兩組各6只,喂養1周后行去勢手術,術后3 d每只小鼠予肌注青霉素40 000 U預防感染。兩組小鼠術后第4天腹腔麻醉下予右側股骨干骺端微量注射慢病毒scr-shRNA及Gαi1/3-shRNA(每側1 μL,滴度為3×108U/mL),持續7 d。
1.3.6 骨骼樣本收集及Micro CT檢測 4周后將兩組小鼠脫頸處死,剝取右側股骨予多聚甲醛固定,3 d后行Micro CT檢查。掃描分辨率為10 μm,自生長板底部0.5 mm處開始掃描,應用內置軟件對干骺端松、皮質骨進行圖像重建及骨量測量。
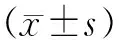
2 結 果
2.1 BMMs中轉染shRNA慢病毒抑制Gαi1/3表達 為探討Gαi1/3在破骨細胞分化過程中的調控作用,采用shRNA慢病毒干擾方法,通過scr-shRNA(GFP-RNA)、Gαi1-shRNA和Gαi3-shRNA轉導至WT BMMs中,24 h后在熒光顯微鏡下觀察病毒轉染情況(見圖1a)。轉染成功后WB證實BMMs中Gαi1/3蛋白表達受到顯著抑制(見圖1b)。

a scr-shRNA、Gαi1-shRNA、Gαi3-shRNA 慢病毒序列及轉染后熒光圖
2.2 敲減Gαi1/3抑制MCSF及MCSF+RANKL誘導Akt-GSK3β-NFATc1通路的活化 scr-shRNA、Gαi1/3-shRNA轉染WT BMMs成功后使用MCSF(50 ng/mL)、RANKL(50 ng/mL)、MCSF(50 ng/mL)+RANKL(50 ng/mL)處理30 min,WB檢測發現與對照組相比,敲減Gαi1/3顯著抑制MCSF及MCSF+RANKL誘導Akt-GSK3β-NFATc1通路活化(見圖2a~b),而對RANKL誘導通路活化影響無顯著抑制作用(見圖2c)。

a MCSF誘導敲減Gαi1/3的BMMs中Akt-GSK3β-NFATc1通路蛋白表達
2.3 敲除Gαi1/3抑制MCSF及MCSF+RANKL誘導Akt-GSK3β-NFATc1通路的活化 為進一步研究Gαi1/3對MCSF等因子活化作用影響,提取CRISPR/Cas9-Gαi1/3-DKO雌性C57BL/6小鼠的BMMs,同樣應用MCSF(50 ng/mL)、RANKL(50 ng/mL)、MCSF(50 ng/mL)+RANKL(50 ng/mL)處理30 min,WB檢測發現敲除Gαi1/3顯著抑制MCSF及MCSF+RANKL誘導Akt-GSK3β-NFATc1通路活化(見圖3a~b),而對RANKL誘導通路活化無顯著抑制作用(見圖3c)。

a MCSF誘導敲除Gαi1/3的BMMs中Akt-GSK3β-NFATc1通路蛋白表達
2.4 敲減Gαi1/3抑制破骨分化關鍵蛋白表達及破骨細胞分化 采用WB進行檢測發現敲減Gαi1/3顯著抑制MCSF(50 ng/mL)+RANKL(50 ng/mL)誘導4 d的破骨細胞分化關鍵蛋白NFATc1的表達(見圖4a),同時顯著抑制破骨細胞生成數量(見圖4b)。

a 敲減Gαi1/3破骨細胞分化關鍵蛋白NFATc1的表達
2.5 敲減Gαi1/3減輕小鼠骨質疏松模型中的骨質丟失 分別向WT小鼠骨質疏松模型股骨干骺端注射scr-shRNA、Gαi1/3-shRNA,4周后行Micro CT檢查敲減Gαi1/3可顯著減輕骨質疏松導致骨質丟失,進一步分析顯示敲減Gαi1/3對于松質骨丟失的挽救作用略高于皮質骨的丟失(見圖5)。

a 小鼠骨質疏松模型股骨干骺端注射scr-shRNA、Gαi1/3-shRNA后Gαi1/3表達
3 討 論
既往研究表明骨質疏松癥主因在于骨代謝失衡骨吸收增多所致,而破骨細胞數量增多及異常活化被證實是骨吸收異常增多的主因[2]。目前治療骨質疏松癥的藥物可導致下頜骨缺血性壞死、骨重建紊亂等不良反應[8],因此進一步了解骨質疏松癥的發病機制,針對性開發更加安全的新型治療藥物顯得愈加迫切[9-10]。
G蛋白由α、β、γ三個亞單位構成,其中Gα蛋白可分為Gi、Gs等6種類型亞基,既往研究顯示G蛋白抑制性α亞基(Gαi蛋白)可以與G蛋白耦聯受體(G protein-coupled receptors,GPCRs)相互結合從而發揮介導下游信號傳遞的作用[11]。筆者所在課題組前期研究發現G蛋白抑制性α亞單位1/3(Gαi1/3)能夠介導多個RTKs諸如BDNF、VEGF、SCF等下游信號轉導,通過敲減Gαi1/3可使其下游PI3K-Akt-mTOR、MAPK-Erk等通路的活化受到抑制[5-7]。
為進一步明確Gαi1/3在同屬于RTKs的MCSF的信號轉導過程中的作用,本研究采用MCSF和RANKL聯合誘導原代BMMs分化破骨細胞,該方法接近破骨細胞體內分化微環境,目前廣泛用于骨質疏松癥進展調控的研究[9]。本研究采用慢病毒敲減及基因敲除策略,獲取scr-shRNA、Gαi1/3-shRNA及Gαi1/3-DKO三種BMMs,研究中發現敲減、敲除Gαi1/3可顯著抑制經典破骨細胞分化信號通路Akt-GSK3β-NFATc1中關鍵蛋白的表達,尤其是終末分化關鍵蛋白NFATc1[12]。同時本實驗發現敲減、敲除Gαi1/3對于RANKL誘導的Akt-GSK3β通路活化無顯著抑制作用,考慮RANKL不屬于RTKs且Gαi1/3未參與核因子NF-κb等主要傳導通路所致,與前期既往研究結果一致[13-16]。本研究Gαi1/3-shRNA BMMs中加入MCSF 50 ng/mL+RANKL 50 ng/mL聯合誘導4 d進行染色顯示生成的破骨細胞的細胞核較少及體積明顯縮小,且生成數量顯著減少,提示敲減Gαi1/3對于BMMs向破骨細胞分化有顯著的抑制作用。最后體內實驗顯示,在小鼠骨質疏松模型中股骨干骺端注射Gαi1/3-shRNA可顯著抑制去勢雌鼠的骨質丟失,且對松質骨的丟失有更強的逆轉趨勢,進一步證實Gαi1/3蛋白在骨質疏松進程中扮演關鍵角色。
既往研究顯示MCSF對破骨細胞祖細胞的存活和增殖是必需的,同時MCSF含量與破骨細胞數量密切相關,MCSF基因缺陷可導致小鼠先天性骨質硬化癥,而外源性補充MCSF可促進骨硬化癥小鼠體內破骨細胞數量增多恢復骨吸收功能[17]。目前對于MCSF調控破骨細胞分化的研究主要集中在MCSF受體c-fms及相關信號通路[18],而本研究著重于探討Gαi1/3蛋白對于破骨細胞分化的調控作用,從而進一步明確MCSF調控破骨細胞分化的分子機制并對其進行干預,對于預防和治療骨質疏松具有重要的臨床意義。
綜上所述,本研究從分子、細胞及在體實驗初步解析Gαi1/3蛋白通過介導Akt-GSK3β-NFATc1信號通路調控破骨細胞分化機制,可為臨床治療骨質疏松提供新的思路和治療靶點。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
學苑創造·A版(2020年9期)2020-10-13 09:41:02
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
汽車工程學報(2017年2期)2017-07-05 08:13:02
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
