miR-125b對肝臟血管新生調控作用的實驗研究
2024-01-08 03:26:56王佳慧黃艷青段雪琳劉艷芳趙鐵建梁天堅
安徽醫科大學學報 2023年12期
王佳慧,鄭 洋,汪 磊,黃艷青,段雪琳,劉艷芳,趙鐵建,梁天堅
肝纖維化(hepatic fibrosis, HF)是肝臟對慢性損傷的應答反應,以細胞外基質(extracellular matrix, ECM)大量沉積為特征[1]。當肝臟受到損傷,肝竇內皮細胞(hepatic sinusoid endothelial cell, HSEC)表型會改變,導致分化抗原簇31(cluster of differentiation 31,CD31)、血管性血友病因子(von Willebrand factor, vWF)以及層粘連蛋白(laminin, LN)和Ⅳ型膠原(type IV collagen, Collagen IV)表達增多,從而介導肝竇毛細血管化發生[2]。肝竇毛細血管化可導致肝細胞損傷和肝星狀細胞(hepatic stellate cell, HSC)活化[3]。活化的HSC可導致大量ECM生成促進肝纖維化的發生發展,進一步表明肝竇毛細血管化是肝纖維化發生的重要病理事件。
miR-125b通過調控HSC的凋亡,減少ECM的沉積發揮抗肝纖維化作用[4]。血管內皮生長因子(vascular endothelial growth factor, VEGF)可通過調節一氧化氮(nitric oxide, NO)介導血管新生[5]。miR-125b作為VEGF的上游分子,可以影響其下游信號傳導[6]。目前圍繞血管新生防治肝纖維化的藥物研究,因為缺少明確靶點而進展緩慢。該研究旨在探究miR-125b對肝臟血管新生的作用,為肝纖維化提供新的作用靶點。
1 材料與方法
1.1 材料與儀器人肝竇內皮細胞(HHSEC)(美國ScienCellTM公司,貨號:5000),Lip2000(上海賽默飛,貨號:12566014),miR-125b相關片段(上海吉瑪制藥技術有限公司),TRIzol(美國Ambion公司,貨號:15596-026),VEGF抗體(貨號:ab52917),CD31抗體(貨號:ab222783),vWF抗體(貨號:ab174290),Collagen IV抗體(貨號:ab6586),LN抗體(貨號:ab154848),雙熒光素酶報告基因檢測試劑盒(貨號:ab228530)購自Abcam公司,無血清培養基(上海賽默飛,貨號:12753018),PBS(北京索萊寶科技有限公司,貨號:P1003),DAF-FMDA(上海碧云天公司,貨號:S0019),多聚甲醛(國藥集團,貨號:80096618),Matrigel(北京索萊寶科技有限公司,貨號:354277),NLRP3野生型(wild type,wt)/突變型(mutant,mut)的熒光素酶報告基因質粒載體(上海Genepharm公司),Easy Pure miRNA Kit(北京全式金生物技術有限公司,貨號:ER601-01-V2),ELISA試劑(上海酶聯生物科技有限公司,貨號:ml064281;ml060878;ml023069;ml062947;ml028571)。RT-PCR儀(美國賽默飛,型號:ABI QuantStudio6),掃描電鏡(日本HITACHI,型號:SU-8010),倒置顯微鏡(日本OLYMPUS,型號:IX51),培養箱(上海賽默飛,型號:311)、臨界點干燥器(美國 tousimis 公司,型號:Autosamdri-931)。
1.2 方法
1.2.1細胞培養 用含5%FBS的ECM培養基進行HHSEC細胞培養,置于5% CO2的37 ℃培養箱中培養,待細胞長至單層后用于實驗。
1.2.2細胞轉染以及分組處理 取生長至單層的HHSEC細胞將其分為空白組、過表達組(mimics組)、過表達對照組(mimics NC組)、抑制組(inhibitor組)、抑制對照組(inhibitor NC組),隨后進行轉染。將細胞接種于24孔板上,第2天當細胞密度達到約30%時,進行miRNA-125b mimics和陰性對照轉染。用無血清培養基將miRNA-125b mimics和陰性對照配成miRNA-DMEM,終濃度為20 nmol/L,輕輕震蕩搖勻后在室溫下孵育5 min,同樣配制濃度為5 μl/ml的Lipofec-tamineTM2000試劑,輕輕震蕩搖勻后在室溫下孵育5 min,隨后將配制好的miRNA-125b mimics和陰性對照分別與Lipofec-tamineTM2000混勻后在室溫下孵育20 s,使之形成mimics-Lipo2000混合物,將100 μl mimics-Lipo2000混合物加入到24板孔中,置于37 ℃、5%CO2的培養箱中培養4~6 h,更換新鮮培養液,繼續培養24 h,以供后續實驗使用。miRNA-125b inhibitor和陰性對照轉染方法與mimics相同,空白組全程使用ECM培養基進行培養,培養條件與其它四組相同。各組細胞用100 μg/L廋素活化,培養48 h,用于各項后續實驗。
1.2.3qRT-PCR檢測miR-125b和U6、VEGF、CD31、vWF、CollagenⅣ、LN和β-actinmRNA表達 將各組細胞按上述處理后,借助TRIzol試劑和Easy Pure miRNA Kit提取RNA,用mRNA逆轉錄試劑盒得到cDNA,進行PCR擴增反應,反應條件:預變性95 ℃ 10 min,變性95 ℃ 15 s,退火/延伸60 ℃ 1 min,共40個循環。用miRNA逆轉錄試劑盒操作完成高效加尾和合成第一鏈cDNA,然后以cDNA為模板,進行擴增,反應條件為:95 ℃ 30 s,1個循環;95 ℃ 5 s,60 ℃ 35 s,40個循環。通過各目的基因片段的擴增曲線確定Ct值;以β-actin、U6為內參基因,采用2-ΔΔCt法分析目標基因mRNA的相對表達量;用qRT-PCR檢測相應分子的表達,miR-125b、U6、VEGF、CD31、vWF、CollagenⅣ、LN和β-actin上下游引物及長度見表1。實驗重復3次。

表1 引物序列表
1.2.4ELISA檢測VEGF、CD31、vWF、CollagenⅣ、LN蛋白表達 將各組細胞按上述處理后,以5×103/ml細胞密度接種至96孔板中,以離心半徑15 cm、1 000 r/min離心20 min,取上清液檢測。按試劑盒說明書操作,在酶標儀450 nm波長下測量吸光度值,計算細胞上清液中VEGF、CD31、vWF、Collagen Ⅳ及LN的含量。實驗重復3次。定量的ELISA實驗檢測蛋白表達更能反映miR-125b的靶向作用。
1.2.5熒光探針檢測NO的表達 將各組細胞按上述處理后,接入12孔板爬片,于37 ℃、5%CO2培養箱中培養48 h,去除細胞培養液,加入1 ml稀釋好的DAF-FMDA,37 ℃細胞培養箱內孵育20 min,用PBS洗滌細胞3次,以去除未進入細胞內的DAF-FMDA,將爬好細胞的玻片浸入4%的多聚甲醛溶液中室溫固定25 min,隨后用PBS洗滌3次,每次5 min,滴加DAPI避光孵育5 min,PBST 5 min×4次洗去多余的DAPI,用含抗熒光淬滅劑的封片液封片,然后在熒光顯微鏡下觀察采集圖像,實驗重復3次。
1.2.6掃描電鏡觀察肝竇內皮細胞的結構 各組孵育的細胞爬片,2.5%的戊二醛固定2 h,0.1 mol/L磷酸緩沖液沖洗3次,1%鞣酸處理1 h,1%四氧化鋨后固定1 h,磷酸鹽緩沖液再次沖洗,梯度乙醇溶液脫水,之后用臨界點干燥器對細胞標本進行臨界點干燥,掃描電鏡觀察樣品,每組選取隨機選取3個視野。
1.2.7血管形成實驗檢測血管新生數量 4 ℃融化Matrigel膠,按每100 μl均勻加至96孔細胞培養板底部,37 ℃孵育60 min,在40 min時取HHSEC,按照5×104/ml密度,接種于96孔細胞培養板,各組經過1.2.2項轉染處理后,在倒置顯微鏡下觀察管腔形成情況,每組選取隨機選取3個視野。
1.2.8雙熒光素酶報告基因實驗驗證miR-125b與VEGF的靶向關系 按照雙熒光素酶報告基因檢測試劑盒說明書,將VEGF野生型(wild type, wt)/突變型載體(mutant, mut)和短鏈非編碼RNA-125b 模擬物(miR-125b mimic)轉染入HHSEC細胞中,培養48 h后,3 500 r/min離心2 min, 小心吸取培養液棄去。PBS洗滌細胞后,再加入細胞裂解液,在室溫下震蕩 5~10 min,收集并在3 000 r/min條件下離心5 min,取上清液進行檢測,實驗重復3次。
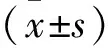
2 結果
2.1 轉染后miR-125b的表達情況細胞轉染結束后,用qRT-PCR檢測轉染各組miR-125b的表達量,結果如圖1所示。轉染miR-125b mimics及inhibitor后,過表達組miR-125b的表達明顯高于空白對照組(F=4.617,P=0.001),同時明顯高于mimics NC組(F=3.321,P=0.002),而 inhibitor 組則沒有檢測到miR-125b表達,表明inhibitor對miR-125b表達具有較好的抑制作用。

圖1 轉染后各組miR-125b表達情況
2.2 miR-125b對VEGF、CD31、vWF、CollagenⅣ、LNmRNA表達的作用與陰性對照組相比,miR-125b mimics轉染后VEGF、CD31、vWF、CollagenⅣ、LNmRNA表達明顯下降(均P<0.05);而miR-125binhibitor 轉染后VEGF、CD31、vWF、CollagenⅣ、LNmRNA表達明顯增加(均P<0.05)。見表2。

表2 miR-125b模擬物和抑制物對VEGF、CD31、vWF、CollagenⅣ、LN mRNA表達的比較
2.3 miR-125b對VEGF、CD31、vWF、CollagenⅣ、LN蛋白表達的作用ELISA檢測發現與陰性對照組相比,miR-125b mimics轉染后VEGF、CD31、vWF、Collagen Ⅳ、LN蛋白表達明顯降低(均P<0.05);而miR-125b inhibitor 轉染后VEGF、CD31、vWF、Collagen Ⅳ、LN蛋白表達明顯增加(均P<0.05)。見表3。

表3 miR-125b模擬物和抑制物對VEGF、CD31、vWF、Collagen Ⅳ、LN蛋白表達的比較
2.4 miR-125b對NO表達的作用與陰性對照組相比,miR-125b mimics轉染后NO平均熒光強度表達下降(t=5.176,P=0.007);而miR-125b inhibitor轉染后NO平均熒光強度表達增加(t=3.752,P=0.020)。見圖2。

圖2 miR-125b模擬物和抑制物對肝竇內皮細胞NO表達的影響
2.5 miR-125b對肝竇內皮細胞表面窗孔數量的作用與陰性對照組相比,miR-125b mimics轉染后肝竇內皮細胞表面窗孔的數增加(t=6.897,P=0.002);而miR-125b inhibitor 轉染后肝竇內皮細胞表面窗孔的數量下降(t=3.001,P=0.046)。見圖3。

圖3 miR-125b模擬物和抑制物對肝竇內皮細胞窗孔數量的影響
2.6 miR-125b對血管新生的作用與陰性對照組相比,miR-125b mimics轉染后血管新生的數量下降(t=3.223,P=0.032);而miR-125b inhibitor 轉染后血管新生的數量增加(t=3.874,P=0.018)。見圖4。

圖4 miR-125b模擬物和抑制物對血管新生數量的影響
2.7 miR-125b對VEGF的靶向作用相比于NC mimics組,VEGF-wt和miR-125b mimics共轉染明顯降低VEGF的3′UTR區熒光酶活性(t=7.148,P=0.002),相比于NC mimics組,VEGF-mut和miR-125b mimics共轉染熒光酶活性差異不顯著(t=1.981,P=0.119)。表明miR-125b mimics通過直接靶向作用于VEGF的3′UTR區域,抑制肝竇內皮細胞中VEGF的表達。見圖5。

圖5 雙熒光素酶報告基因檢測相對熒光強度
3 討論
HF是慢性肝病的中間病理過程,因此探究HF的分子機制具有重要意義[7]。劉聰 等[8]認為肝臟血管新生與肝纖維化的關系密切,實驗證明肝纖維化中病理性血管新生以肝竇毛細血管化為特點,并且發現其早于肝纖維化的形成。有研究[9]表明抑制肝竇毛細血管化的形成,可以抑制肝纖維化。同時發現失窗孔化的HSEC 即肝竇毛細血管化的發生會加重HSC活化[10]。
本研究成功構建miR-125b高表達和低表達的細胞模型。后續的qRT-PCR檢測發現,miR-125b mimics可以抑制VEGF、CD31、vWF、CollagenⅣ和LN的mRNA表達,miR-125b inhibitor可以增加VEGF、CD31、vWF、CollagenⅣ和LN的mRNA表達。ELISA檢測發現,miR-125b mimics可以抑制VEGF、CD31、vWF、Collagen Ⅳ和LN的蛋白表達,miR-125b inhibitor可以增加VEGF、CD31、vWF、Collagen Ⅳ和LN的蛋白表達。本研究雙熒光素酶報告基因實驗發現,miR-125b可以結合VEGF mRNA的3′UTR區域,調控其表達,表明miR-125b可以通過靶向VEGF,從而影響內皮細胞標志物CD31和vWF以及連續性基底膜標志物Collagen Ⅳ和LN的表達。本研究熒光探針實驗檢測發現,miR-125b mimics可以抑制NO的表達,miR-125b inhibitor可以增加NO的表達。研究[11]表明NO可以通過血管舒張來調節血管生成因子,從而調節血管生成。血管新生與肝纖維化相促相伴,是肝纖維化重要的早期病變。本研究血管新生實驗也發現,miR-125b mimics可以抑制血管新生,miR-125b inhibitor可以促進血管新生。表明miR-125b調控NO表達,可能是其介導血管新生的關鍵策略。本研究發現miR-125b mimics可以增加肝竇內皮細胞窗孔的數量,miR-125b inhibitor可以減少肝竇內皮細胞窗孔的數量。研究[12]顯示肝細胞損傷可導致肝臟缺氧,使肝竇內皮細胞線粒體功能衰竭,細胞骨架塌陷,導致其失窗孔化發生。并且失窗孔化也促進了肝竇連續基底膜的形成,破壞了肝細胞和肝血竇之間營養物質的交換[13]。有研究[14]表明以血管生成與重構為靶點進行藥物干預,可能成為抑制或逆轉肝纖維化的有效策略。
猜你喜歡
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
中學生數理化·中考版(2022年11期)2022-02-16 07:01:20
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
發明與創新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
