姜黃素可能通過上調(diào)Prdx6蛋白表達(dá)抑制軟骨細(xì)胞鐵死亡
2024-01-08 03:11:44周富麗付萬進(jìn)周仁鵬
安徽醫(yī)科大學(xué)學(xué)報(bào) 2023年12期
陳 凡,周富麗,陳 勇,付萬進(jìn),周仁鵬,胡 偉,魯 超
骨關(guān)節(jié)炎(osteoarthritis,OA)是一種以軟骨損傷為主的常見關(guān)節(jié)疾病,其特征是關(guān)節(jié)軟骨細(xì)胞外基質(zhì)(extracellular matrix, ECM)降解、關(guān)節(jié)軟骨退化和骨贅的形成[1-2]。鐵死亡是一種鐵離子依賴的,以脂質(zhì)活性氧(reactive oxygen species, ROS)累積,線粒體皺縮為主要標(biāo)志的新型程序性細(xì)胞死亡方式,許多抑制或緩解脂質(zhì)過氧化的藥物均可抑制鐵死亡[3],從而能夠有效干預(yù)疾病的發(fā)展,因此成為當(dāng)前研究的熱點(diǎn)。姜黃素(curcumin,Cur)具有抗氧化、抗菌以及與非甾體類抗炎藥物類似的抗炎活性,已經(jīng)廣泛應(yīng)用于OA的治療中[4]。Cur的抗氧化作用與多種信號(hào)通路有關(guān)[5],研究報(bào)道[6]Cur在調(diào)控過氧化物酶6(peroxiredoxin6,Prdx6)表達(dá)在抑制內(nèi)質(zhì)網(wǎng)應(yīng)激和線粒體凋亡中發(fā)揮重要作用,但其在軟骨細(xì)胞鐵死亡中的作用及機(jī)制目前尚不清楚。Prdx6作為過氧化物還原酶家族的一員,是發(fā)揮抗氧化應(yīng)激與維持細(xì)胞磷脂平衡的關(guān)鍵蛋白[7]。因此,本研究旨在探討Cur調(diào)控Prdx6蛋白表達(dá)在抑制軟骨細(xì)胞鐵死亡過程中的作用及可能的機(jī)制,為OA的防治提供新的靶點(diǎn)與策略。
1 材料與方法
1.1 材料
1.1.1細(xì)胞株及實(shí)驗(yàn)動(dòng)物 C28/I2人正常軟骨細(xì)胞系,由ATCC公司提供;SD大鼠20只,雄性,體質(zhì)量160~180 g,鼠齡6~8周,由濟(jì)南朋悅實(shí)驗(yàn)動(dòng)物繁育有限公司提供,合格證號(hào):SCXK(魯)20190003。于溫度(22±3)℃,濕度40%~70%條件下12 h光照12 h黑暗交替飼養(yǎng)。
1.1.2人膝關(guān)節(jié)軟骨組織樣本 2019年—2021年臨床收集正常人(5例)和OA患者(5例)置換下來的膝關(guān)節(jié)軟骨組織標(biāo)本,排除因有膝關(guān)節(jié)感染性疾病、類風(fēng)濕性關(guān)節(jié)炎等其他疾病進(jìn)行膝關(guān)節(jié)置換的軟骨標(biāo)本。本研究通過安徽理工大學(xué)生物醫(yī)學(xué)研究倫理委員會(huì)審核(倫理批號(hào):2021020)。
1.1.3藥品和試劑 Cur(美國Sigma-Aldrich公司,貨號(hào):08511);Erastin(美國Selleck生物科技有限公司,貨號(hào):S7242);胎牛血清(fetal bovine serum,FBS)(美國Invitrogen生命技術(shù)有限公司,貨號(hào):C0235);DMEM 高糖培養(yǎng)基(以色列BI生物科技公司,貨號(hào):06-1055-57-1ACS);乳酸脫氫酶(lactate dehydrogenase,LDH)檢測試劑盒(瑞士Hoffmann-La Roche有限責(zé)任公司,貨號(hào):11644793001);噻唑藍(lán)(thiazolyl blue tetrazolium bromide,MTT)(上海碧云天生物技術(shù)有限公司,貨號(hào):ST1537);BODIPY 581/591 C11熒光探針(美國Invitrogen生命技術(shù)有限公司,貨號(hào):D3861);總谷胱甘肽(glutathione,GSH)檢測試劑盒(上海碧云天生物技術(shù)有限公司,貨號(hào):S0052);抗SLC7A11抗體(英國abcam公司,貨號(hào):ab307601);抗GPX4抗體(英國abcam公司,貨號(hào):ab125066);抗Prdx6抗體(江蘇親科生物研究中心有限公司,貨號(hào):DF6765);抗β-actin抗體(北京中杉金橋生物技術(shù)有限公司,貨號(hào):TA-09);抗ACSL4抗體(美國Santa Cruze公司,貨號(hào):sc-365230);抗FTH抗體(美國Santa Cruze公司,貨號(hào):sc-376594)。
1.1.4儀器 電泳儀(型號(hào):EPS 600,上海天能電子有限公司);CO2培養(yǎng)箱(型號(hào):371Steri-Cycle,美國Thermo Fisher科技公司);酶標(biāo)儀(型號(hào):800TS,美國ThermoFisher科技公司);流式細(xì)胞儀(型號(hào):CytoFLEX,美國Thermo Fisher科技公司);倒置熒光顯微鏡(型號(hào):AxioVert.A1,德國Carl Zeiss股份公司);超低溫冰箱(型號(hào):DW-86L626,青島海爾股份有限公司);化學(xué)發(fā)光分析儀(型號(hào):FINEDO X6,上海天能電子有限公司);離心機(jī)(型號(hào):No. 5417R,德國Eppendorf股份公司)。
1.2 方法
1.2.1膝關(guān)節(jié)軟骨組織染色
1.2.1.1OA大鼠模型的建立及組織切片 20只SD大鼠隨機(jī)分為2組:正常大鼠組、OA大鼠模型組。采用左膝前交叉韌帶切除術(shù)+內(nèi)側(cè)半月板摘除術(shù)建立左膝 OA 模型。取材后膝關(guān)節(jié)組織樣本經(jīng)4%多聚甲醛固定,10%EDTA溶液脫鈣后進(jìn)行組織浸蠟、包埋、切片。人膝關(guān)節(jié)軟骨組織切片同大鼠。
1.2.1.2Safranin O/Fast Green染色 大鼠膝關(guān)節(jié)切片脫蠟水化后,按常規(guī)方法進(jìn)行Safranin O/Fast Green染色,倒置熒光顯微鏡拍照保存。
1.2.1.3HE染色 大鼠膝關(guān)節(jié)切片脫蠟水化后,按常規(guī)方法進(jìn)行HE染色,倒置熒光顯微鏡拍照保存。
1.2.1.4免疫組織化學(xué)染色 大鼠和人軟骨組織切片脫蠟水化后孵育Prdx6和GPX4蛋白一抗(1 ∶50)過夜,37 ℃孵育二抗(1 ∶150) 30 min,DBA顯色3 min,蘇木精溶液復(fù)染2 min,脫水封片后顯微鏡下觀察。
1.2.2C28/I2軟骨細(xì)胞系培養(yǎng)和傳代 C28/I2軟骨細(xì)胞系用含有10%胎牛血清和1%雙抗的DMEM培養(yǎng)基,在含有5% CO2,溫度為37 ℃的條件下培養(yǎng),觀察細(xì)胞貼壁情況至達(dá)到對(duì)數(shù)增長期,用0.25%胰酶進(jìn)行消化傳代分組,分為:Control組、Erastin組、Cur組、Erastin+Cur組。
1.2.3MTT法檢測細(xì)胞活力 按1.2.2項(xiàng)中分組情況,在96孔板中用2.5 μmol/L Erastin和不同濃度Cur(0、0.312、0.625、1.25、2.5、5、10、20和40 μmol/L)分別處理Erastin組、Cur組和Erastin+Cur組的C28/I2軟骨細(xì)胞系8 h,顯微鏡下觀察細(xì)胞死亡數(shù)量至50%時(shí),每孔加入MTT檢測液,孵育4 h后加入二甲基亞砜溶解,酶標(biāo)儀選定波長490 nm和570 nm,測定吸光度值計(jì)算細(xì)胞活力,確定最宜Cur加藥濃度為20 μmol/L。
1.2.4LDH法檢測細(xì)胞毒性 按1.2.2項(xiàng)中分組情況,在24孔板中用2.5 μmol/L Erastin和20 μmol/L Cur處理Erastin組、Cur組和Erastin+ Cur組C28/I2軟骨細(xì)胞系8 h,依據(jù)羅氏LDH試劑盒使用說明書,使用酶標(biāo)儀測定波長在492 nm和620 nm處的吸光度,確定LDH的釋放值。
1.2.5流式細(xì)胞術(shù)檢測脂質(zhì)ROS 按1.2.2項(xiàng)中分組情況,在6孔板中用2.5 μmol/L Erastin和20 μmol/L Cur處理Erastin組、Cur組和Erastin+Cur組C28/I2軟骨細(xì)胞系8 h。用2 μmol/L BODIPY 581/591C11探針染色30 min,胰酶消化并于1 500 r/min,4 ℃離心5 min收集細(xì)胞沉淀,通過流式細(xì)胞儀進(jìn)行分析,數(shù)據(jù)用FlowJo軟件分析。
1.2.6總GSH試劑盒檢測GSH含量 按1.2.2項(xiàng)中分組情況,在6孔板中用2.5 μmol/L Erastin和20 μmol/L Cur處理Erastin組、Cur組和Erastin+Cur組C28/I2軟骨細(xì)胞系8 h。胰酶消化并1 500 r/min,4 ℃,離心5 min收集細(xì)胞沉淀,實(shí)驗(yàn)按碧云天生物的試劑盒使用說明進(jìn)行。
1.2.7Western blot檢測ACSL4、SLC7A11、Prdx6、FTH、GPX4蛋白表達(dá) 按1.2.2項(xiàng)中分組情況,在6孔板中用2.5 μmol/L Erastin和20 μmol/L Cur處理Erastin組、Cur組和Erastin+Cur組C28/I2軟骨細(xì)胞系8 h。用RIPA蛋白裂解液提取軟骨細(xì)胞總蛋白,根據(jù)不同的蛋白分子量在合適的SDS-PAGE凝膠中進(jìn)行電泳(分離膠80 V 30 min,濃縮膠120 V 60 min)。再轉(zhuǎn)移到聚偏二氟乙烯(PVDF)膜上用5%脫脂牛奶封閉,置于抗ACSL4(1 ∶500)、抗SLC7A11(1 ∶1 000)、抗FTH(1 ∶500)、抗GPX4(1 ∶500)、抗Prdx6(1 ∶1 000)一抗孵育過夜,用辣根過氧化物酶(HRP)標(biāo)記的山羊抗兔/鼠IgG二抗(1 ∶500)孵育1h,配制ECL混合顯影液平鋪在膜表面,化學(xué)發(fā)光分析儀中顯影。使用ImageJ軟件對(duì)Western blot條帶進(jìn)行量化。
1.2.8Cur分子與Prdx6蛋白的分子對(duì)接 利用分子對(duì)接技術(shù)Biovia Discovery Studio2016評(píng)價(jià)Cur分子與Prdx6蛋白的對(duì)接,并展示Cur分子與Prdx6蛋白相互作用的二維結(jié)構(gòu)。
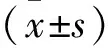
2 結(jié)果
2.1 OA大鼠關(guān)節(jié)軟骨病理染色和Western blot法檢測結(jié)果正常大鼠和OA大鼠Safranin O/Fast Green病理染色結(jié)果顯示,正常大鼠關(guān)節(jié)軟骨表面完整、平滑,軟骨細(xì)胞排列正常、數(shù)量正常、核染正常,軟骨基質(zhì)著色鮮紅;OA大鼠模型關(guān)節(jié)面糜爛、缺損、變形,軟骨結(jié)構(gòu)消失,軟骨層進(jìn)行性喪失或者損壞,軟骨細(xì)胞數(shù)量減少、排列紊亂、有簇集現(xiàn)象,軟骨基質(zhì)著色變淺。HE病理染色結(jié)果顯示正常大鼠膝關(guān)節(jié)軟骨表面光滑連續(xù)、無損傷,而OA大鼠模型關(guān)節(jié)軟骨的形態(tài)學(xué)出現(xiàn)明顯的改變,關(guān)節(jié)軟骨表面不連續(xù),軟骨基質(zhì)損傷、斷裂,軟骨組織被侵蝕,關(guān)節(jié)表面輪廓發(fā)生典型的病理改變(圖1A)。免疫組織化學(xué)染色觀察Prdx6和鐵死亡標(biāo)志物GPX4蛋白在OA大鼠模型中的表達(dá),結(jié)果顯示OA大鼠模型關(guān)節(jié)軟骨中Prdx6和GPX4蛋白黃染變淡(圖1B)。以上結(jié)果說明OA發(fā)病進(jìn)程中存在鐵死亡。
2.2 Prdx6和鐵死亡標(biāo)志物GPX4蛋白在OA患者關(guān)節(jié)軟骨中的表達(dá)情況Western blot檢測結(jié)果顯示與正常人相比,OA患者中Prdx6(t=3.38,P<0.01)和GPX4(t=3.66,P<0.01)蛋白表達(dá)明顯降低(圖2A、B)。免疫組織化學(xué)染色觀察Prdx6和鐵死亡標(biāo)志物GPX4蛋白在OA患者中的表達(dá),結(jié)果顯示OA患者關(guān)節(jié)軟骨中Prdx6和GPX4蛋白黃染變淡(圖2B)。以上結(jié)果進(jìn)一步表明OA發(fā)病進(jìn)程中存在鐵死亡,且Prdx6蛋白表達(dá)的降低可能與鐵死亡有關(guān)。

圖2 OA患者軟骨組織中Prdx6和GPX4的表達(dá)情況(n=5)
2.3 Cur對(duì)Erastin誘導(dǎo)的C28/I2軟骨細(xì)胞系鐵死亡細(xì)胞活力和細(xì)胞毒性的影響Erastin和不同濃度Cur處理C28/I2軟骨細(xì)胞系8 h,通過MTT法檢測細(xì)胞活力,結(jié)果顯示,與Erastin組比較,Cur能濃度依賴性抑制Erastin誘導(dǎo)的關(guān)節(jié)軟骨細(xì)胞活力的降低,提高細(xì)胞活力,且在Cur濃度為20 μmol/L時(shí)保護(hù)效果顯著(t=9.00,P<0.01)(圖3A)。LDH法檢測細(xì)胞毒性,結(jié)果顯示(圖3B)Erastin組關(guān)節(jié)軟骨細(xì)胞中LDH釋放增加(t=6.78,P<0.01),表明Erastin增加了關(guān)節(jié)軟骨細(xì)胞的毒性,而用20 μmol/L Cur處理后可以明顯降低軟骨細(xì)胞LDH的釋放(t=6.55,P<0.01),表明Cur可以降低Erastin所致的軟骨細(xì)胞毒性。

圖3 Cur對(duì)Erastin誘導(dǎo)的軟骨細(xì)胞鐵死亡細(xì)胞活力(n=6)和細(xì)胞毒性(n=4)的影響
2.4 Cur對(duì)Erastin誘導(dǎo)的C28/I2軟骨細(xì)胞系鐵死亡脂質(zhì)ROS產(chǎn)生和GSH含量的影響流式細(xì)胞術(shù)量化比較FITC-A+脂質(zhì)過氧化細(xì)胞最大百分比,結(jié)果顯示(圖4A、B)Erastin處理后軟骨細(xì)胞脂質(zhì)ROS明顯增加(t=9.64,P<0.01),而Cur處理后能明顯降低脂質(zhì)ROS的水平(t=4.76,P<0.01)。GSH結(jié)果顯示(圖4C)Erastin處理后軟骨細(xì)胞內(nèi)GSH含量明顯降低(t=115.20,P<0.01),而Cur處理后軟骨細(xì)胞內(nèi)GSH含量明顯提高(t=98.66,P<0.01)。

圖4 Cur對(duì)Erastin誘導(dǎo)的軟骨細(xì)胞脂質(zhì)ROS水平和GSH含量的影響(n=3)
2.5 Cur對(duì)Erastin誘導(dǎo)的C28/I2軟骨細(xì)胞系Prdx6和鐵死亡相關(guān)蛋白表達(dá)的影響Western blot檢測結(jié)果顯示在Erastin誘導(dǎo)的軟骨細(xì)胞鐵死亡過程中,Cur處理可以降低軟骨細(xì)胞ACSL4(t=11.87,P<0.01)蛋白表達(dá),上調(diào)SLC7A11(t=5.19,P<0.01)、Prdx6(t=3.77,P<0.05)、FTH(t=4.11,P<0.05)、GPX4(t=8.06,P<0.01)蛋白表達(dá)。說明Cur可能通過上調(diào)Prdx6蛋白表達(dá)抑制C28/I2軟骨細(xì)胞系鐵死亡。見圖5。

圖5 Cur對(duì)Erastin誘導(dǎo)的軟骨細(xì)胞鐵死亡Prdx6和鐵死亡蛋白表達(dá)的影響(n=3)
2.6 Cur分子與Prdx6蛋白的分子對(duì)接情況通過分子對(duì)接技術(shù)探究Cur分子與Prdx6蛋白的結(jié)合情況,結(jié)果顯示Cur分子可以與Prdx6蛋白結(jié)合,二者之間存在相互作用力:范德華力;Cur分子與Prdx6蛋白163位VAL纈氨酸殘基有Pi-Sigma鍵結(jié)合,與LYS144、LEU167、ALA171氨基酸殘基有Pi-Alkyl鍵結(jié)合。見圖6。

圖6 Cur分子與Prdx6蛋白分子對(duì)接圖
3 討論
OA因其病因和發(fā)病機(jī)制的復(fù)雜性,目前臨床上針對(duì)OA的預(yù)防和根治尚無有效的治療藥物和治療方法,因此OA發(fā)生發(fā)展的分子機(jī)制亟待深入探究。軟骨細(xì)胞作為關(guān)節(jié)軟骨中唯一的細(xì)胞類型,參與軟骨基質(zhì)的合成[8]。生理情況下,軟骨細(xì)胞的分化與死亡及軟骨基質(zhì)的合成和分解維持動(dòng)態(tài)平衡。而在OA的發(fā)病進(jìn)程中,這種平衡被打破,關(guān)節(jié)組織部位大量的炎癥、氧化應(yīng)激等因素誘發(fā)軟骨細(xì)胞過度死亡,進(jìn)而引起關(guān)節(jié)功能的喪失。OA發(fā)病進(jìn)程中存在凋亡、自噬、壞死等類型的細(xì)胞死亡方式,然而靶向抑制這些類型的細(xì)胞死亡并不能有效抑制軟骨損傷,提示OA病程中可能存在其他類型的細(xì)胞死亡方式。因此,對(duì)軟骨細(xì)胞死亡方式及其調(diào)控機(jī)制的研究將有助于OA的防治。
本研究表明OA患者和OA大鼠發(fā)病進(jìn)程存在鐵死亡,Prdx6蛋白表達(dá)的降低可能與鐵死亡有關(guān)。研究[9]報(bào)道Cur通過抑制NF - κB信號(hào)通路抑制軟骨細(xì)胞凋亡從而發(fā)揮軟骨保護(hù)作用,且Cur在一些癌細(xì)胞鐵死亡中也發(fā)揮作用,但其在軟骨細(xì)胞鐵死亡中的作用未見報(bào)道。臨床研究[10]表明OA患者血清中脂質(zhì)ROS較正常人明顯增加,并伴有GSH水平的降低。本研究表明Cur處理減少了脂質(zhì)ROS的產(chǎn)生,提高了軟骨細(xì)胞內(nèi)GSH含量。鐵死亡的發(fā)生依賴于脂質(zhì)ROS的累積及其介導(dǎo)的脂質(zhì)過氧化反應(yīng)[11],因此通過調(diào)控細(xì)胞內(nèi)ROS的水平及其下游信號(hào)級(jí)聯(lián)效應(yīng),能夠干預(yù)細(xì)胞鐵死亡的發(fā)生。
此外,本研究結(jié)果顯示Cur處理后ACSL4蛋白表達(dá)降低,Prdx6、SLC7A11、FTH、GPX4蛋白表達(dá)增加。GPX4能夠利用GSH將過氧化的脂質(zhì)還原為無毒性的脂醇,抑制ACSL4可減少多不飽和脂肪酸整合到磷脂上,且SLC7A11作為組成System xc-的兩個(gè)亞基之一,可以促進(jìn)胱氨酸合成GSH[12]進(jìn)而抑制軟骨細(xì)胞鐵死亡。Prdx6作為一種過氧化還原蛋白,它不僅結(jié)構(gòu)獨(dú)特,而且具有谷胱甘肽過氧化物酶活性和鈣非依賴性磷脂酶活性,其活性調(diào)控受到亞細(xì)胞定位、底物結(jié)合及翻譯后修飾等因素影響。通過不同活性功能參與多種疾病的發(fā)生與發(fā)展,在腫瘤、炎性疾病及缺血性腦卒中等疾病中具有重要作用。研究[13]結(jié)果顯示在腫瘤細(xì)胞鐵死亡過程中,Prdx6能夠發(fā)揮清除脂質(zhì)ROS的功能,是一個(gè)新的腫瘤細(xì)胞鐵死亡的負(fù)性調(diào)控因子。本研究結(jié)果顯示Cur和Prdx6之間存在較強(qiáng)的相互作用,結(jié)合Cur對(duì)軟骨細(xì)胞鐵死亡的抑制作用,表明Cur可能通過上調(diào)鐵死亡過程中Prdx6蛋白表達(dá)抑制軟骨細(xì)胞鐵死亡。
綜上所述,本研究表明Cur可能通過上調(diào)鐵死亡過程中Prdx6蛋白的表達(dá),促進(jìn)Prdx6清除脂質(zhì)ROS,進(jìn)而抑制Erastin誘導(dǎo)的軟骨細(xì)胞鐵死亡。后續(xù)還需以Prdx6為靶點(diǎn)探討其在Cur抑制軟骨細(xì)胞鐵死亡發(fā)生發(fā)展中的作用及其可能的分子機(jī)制,為OA的防治提供新思路。
