低聚半乳糖植物乳桿菌發酵乳的代謝組學分析及其腸道菌群調節活性研究
2024-01-16 08:51:28戶行宇賴田甜劉同吉薛瑞陳雅璐楊貞耐
中國乳品工業 2023年12期
戶行宇,賴田甜,劉同吉,薛瑞,陳雅璐,楊貞耐
(北京工商大學北京老年營養與健康教育部重點實驗室北京食品營養與人類健康高精尖創新中心北京市食品添加劑工程技術研究中心,北京 100048)
0 引 言
低聚半乳糖(Galacto-oligosaccharide,GOS)是一類通過β-半乳糖苷酶的水解和轉半乳糖基活性,將水解乳糖得到的多個半乳糖單元以β-(1-4)、β-(1-6)或β-(1-3)等糖苷鍵連接在半乳糖或葡萄糖單元,得到聚合度(DP)在2~8 之間的寡糖混合物[1-2]。GOS 通常被稱為“雙歧桿菌生長因子”,其不但可以促進腸道內有益菌的增殖,還可以抑制大腸桿菌、致病性梭狀芽胞桿菌等有害菌的繁殖,起到平衡腸道菌群的作用[3-4]。研究發現,食用GOS 會在結腸中產生短鏈脂肪酸(SCFA),包括丙酸、乙酸和丁酸等酸性物質。這些代謝物,尤其是丁酸,在結直腸癌的治療中發揮著重要的保護作用[5]。
傳統發酵乳一般由嗜熱鏈球菌(Thermophilus)、德氏乳桿菌保加利亞亞種(Lactobacillus)或雙歧桿菌(Bifidobacterium)發酵生成,相比于鮮牛乳因其含有大量的益生菌,可以改善腸道健康,備受消費者青睞[6],植物乳桿菌能夠產生乳酸菌素、過氧化氫、雙乙酰等多種天然活性物質,具有維持腸道內菌群平衡的作用[7]。但是,目前關于GOS 對植物乳桿菌發酵牛乳的代謝產物的影響以及GOS 發酵乳調節腸道菌群活性研究較少[8]。因此,本文利用課題組前期制備的GOS 以及分離自內蒙古傳統奶酪的植物乳桿菌NMGL2 制備GOS 發酵乳[9-10],采用代謝組學方法分析該發酵乳的差異代謝物,并采用體外人體糞便共培養模型,探究GOS 植物乳桿菌發酵乳的腸道菌群調節活性,為進一步了解GOS 對發酵乳功能性的影響提供參考。
1 材料與方法
1.1 實驗材料與試劑
鮮牛乳,北京三元股份有限公司;植物乳桿菌(Lactobacillus plantarum)NMGL2(保藏編號為CGMCC No. 18495),-80 ℃甘油管保存;低聚半乳糖(GOS,90.17%),實驗室制備;MRS 培養基,北京奧博星生物技術有限公司;甲醇、L-2-氯苯丙氨酸、乙腈、異丙醇、甲酸、丙醇,賽默飛公司;氯化鈉、酵母提取物、蛋白胨、膽鹽、磷酸緩沖液(PBS),索萊寶生物科技有限公司;血紅素、維生素K1、牛膽鹽、半胱氨酸,國藥集團化學試劑有限公司。
1.2 儀器與設備
UHPLC -Q Exactive HF-X 超高效液相色譜串聯傅里葉變換質譜,賽默飛公司;HSS T3 色譜柱,Waters 公司;JXDC-20 氮氣吹掃儀,上海凈信實業發展有限公司;LNG-T88 臺式快速離心濃縮干燥器,太倉市華美生化儀器廠;Wonbio-96c 高通量組織破碎儀,上海萬柏生物科技有限公司;SBL-10TD 超聲波清洗機,寧波新芝生物科技股份有限公司;厭氧培養盒,三菱化學公司;HZQ-Q 數顯pH 計,哈爾濱市東聯電子有限公司。
1.3 方法
1.3.1 GOS 植物乳桿菌發酵乳的制備
將-80 ℃甘油管保存的植物乳桿菌NMGL2 以3%(V/V)的接菌量接種至冷卻后的MRS 液體培養基中,37 ℃條件下靜置培養48 h;活化2 代后,獲得菌懸液(菌懸液的菌濃度為1010CFU/mL)。向鮮牛乳中分別添加0、2%、4%、6%(m/V)的GOS,90 ℃下巴氏消毒殺菌10 min,然后冷卻至37 ℃,將活化好的菌懸液接種至巴氏殺菌奶中,37 ℃靜置發酵24 h。發酵后的樣品于-80 ℃冷凍保存。
1.3.2 GOS 植物乳桿菌發酵乳非靶向代謝組學分析
將-80 ℃保存的4 組不同GOS 添加量的樣品取出,室溫解凍,分別命名為GOS_0、GOS_2、GOS_4、GOS_6。取100 μL 發酵乳樣品加入400 μL 提取液(V甲醇∶V水=4∶1)含0.02 g/L 的內標(L-2-氯苯丙氨酸)進行代謝產物提取。冷凍組織研磨儀研磨,然后低溫超聲提取。將樣品靜置于-20 ℃,離心30 min 后取上清液,采用超高效液相色譜串聯傅里葉變換質譜UHPL-Q Exactive HF-X 系統進行LC-MS 上機分析[11]。
色譜條件:HSS T3 色譜柱(100 mm×2.1 mm,1.8 μm),柱溫設為40 ℃。流動相A 組成:95%水+5%乙腈(含0.1%甲酸),流動相B 組成:47.5%乙腈+47.5%異丙醇+5%水(含0.1%甲酸)。分離梯度:流動相B 從0 逐步升至100%,流速從0.4 mL/min 升至0.6 mL/min;再逐步將流動相B 從100% 降至0,流速調整為0.4 mL/min。質譜條件:選用正負離子的掃描模式,質量掃描范圍m/z:70~1 050。正離子電壓選用3 500 V,負離子電壓選用2 800 V,MS1 分辨率70 000,MS2 分辨率17 500[12]。
質控樣本(Quality control,QC)是通過將所有樣品的等體積混合制成混合樣品。在整個分析過程中,間隔插入,以提供一組可評估重復性的數據[11]。完成上機之后,對LC-MS 原始數據進行處理。將MS 和MSMS的質譜信息與代謝公共數據庫HMDB 和Metlin 數據庫進行匹配整理,從而得到代謝物的相關信息[12]。將整理后的數據上傳至美吉生物云平臺(https://cloud.majorbio.com)進行相關的數據分析。采用人類代謝組數據庫(HMDB)進行代謝物分類,通過KEGG 數據庫進行代謝通路的注釋,從而篩選得到差異代謝物所參與的通路[14]。
1.3.3 GOS 植物乳桿菌發酵乳對腸道菌群的影響
(1)發酵乳體外模型消化處理。參照郝曉娜等[8]的方法稍作修改。將10 mL 發酵乳與10 mL 的NaCl(35 mmol/L)水溶液混合制成發酵乳溶液,模擬胃液環境水解1 h,然后模擬腸道環境消化1 h,最終得到體外模擬消化后的樣品。
(2)BCM 培養基的配制。參照郝曉娜等[8]的方法稍作修改。依次稱取酵母提取物、蛋白胨、膽鹽等培養基組成成分,用蒸餾水定容至2 L,用蒸餾水定容至2 L,用1 mol/L 乙酸調pH 至7.0,滅菌(121 ℃,15 min)后取出備用。
糞便共培養。實驗當天需要收集健康成人(2 男1女,年齡20~28 歲,BMI 值均在20.0~23.5 之間)的糞便,志愿者無飲酒和吃藥的情況,近期內未服用過抗生素等藥類,且未患有慢性腸道類疾病。將中段糞便與PBS 緩沖液震蕩混勻,在厭氧培養箱中,利用PBS緩沖溶液將糞懸液濃度配制成20%(m/V),過濾收集濾液,2 h 內完成糞懸液的處理。將體外模擬消化后的樣品與糞懸液以1∶2 的比例混合,加入BCM 培養基,將其置于厭氧工作站中培養,培養4、12 h 取樣,最終的樣品分組如表1 所示。將培養液于10 000g、4 ℃下離心20 min,從而去除上清液,獲得沉淀的菌體,利用16S rRNA 測定菌群組成的多樣性。將測序篩選獲得的有效序列進行OUT 的劃分聚類,對糞便樣本中菌群組成的多樣性及樣本之間的差異進行分析。
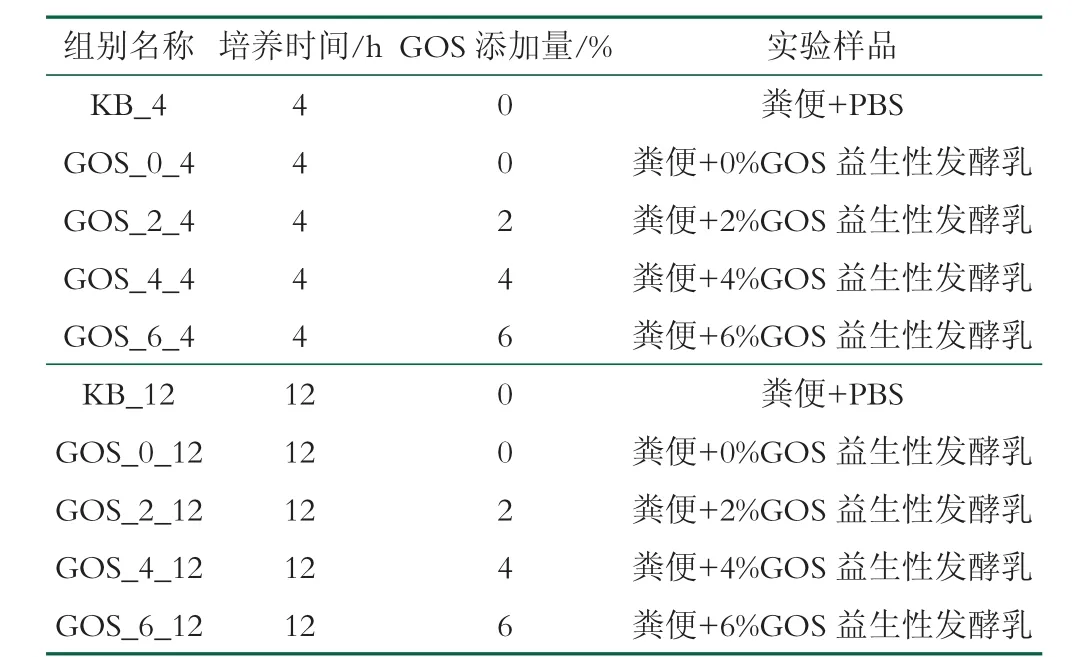
表1 實驗分組
1.3.4 數據分析
每個樣品平行測試3 次,數據以平均值±標準誤表示,采用Excel、IBM SPSS Statistics 25 和origin 8.5 軟件進行處理。P<0.05 表示具有統計學差異。
2 結果與分析
2.1 代謝組學結果分析
2.1.1 代謝產物的主成分分析
PCA 分析(Principal Component Analysis),即主成分分析,是一種對大量實驗數據進行降維簡化分析的技術,得分圖中各點相距越近說明樣本之間越相似,2樣本相距越遠說明樣本之間差異性越高[13]。本研究利用LC-MS 對GOS 不同添加量的益生性發酵乳中的代謝產物組成進行分析。由圖1 可知,在正負離子掃描模式下全部QC 樣品聚集成一簇,且與受試樣品的區分明顯,說明該數據采集過程中儀器穩定性非常好,所得數據準確可靠。在陽離子、陰離子模式下,第1、2 主成分的累計解釋量分別為78.5%、83.8%。且不同樣品組間區分較明顯,距離較大,說明隨著GOS 添加量的增加,發酵乳體系內代謝物組成具有明顯的差異性[14]。

圖1 發酵乳代謝產物PCA 分析
2.1.2 代謝產物的OPLS-DA 分析
OPLS-DA 是一種在PCA 的基礎上再進一步計算分析的方法,該方法能夠盡可能地去除未控制變量對數據的影響,更好對組間差異進行區分[15]。圖2、圖3、圖4 分別為GOS_0 與GOS_2、GOS_0 與GOS_4、GOS_0 與GOS_6 代謝產物的OPLS-DA 分析圖。由圖可知,添加GOS(2%、4%、6%)的益生性發酵乳與未添加GOS 的發酵乳在OPLS-DA 圖上的距離較大,都能夠實現組間的完全區分,同時在陽離子、陰離子模式下第一、二主成分的解釋度較高,說明添加GOS的益生性發酵乳與未添加GOS 的發酵的代謝產物存在著明顯的差異。添加GOS 的益生性發酵乳與未添加GOS 的發酵乳OPLS-DA 判別分析的參數結果如表2 所示,R2X、R2Y和Q2分別表示模型的解釋率和預測率,數值越是接近于1,表示該模型越為理想,對樣本的解釋率和預測率越好[16]。由表2 可知GOS_0 與GOS_2、GOS_0 與GOS_4、GOS_0 與GOS_6 這3 個模型的解釋率和預測率均在0.8 以上,進一步證實3種實驗模型的區分程度較好。
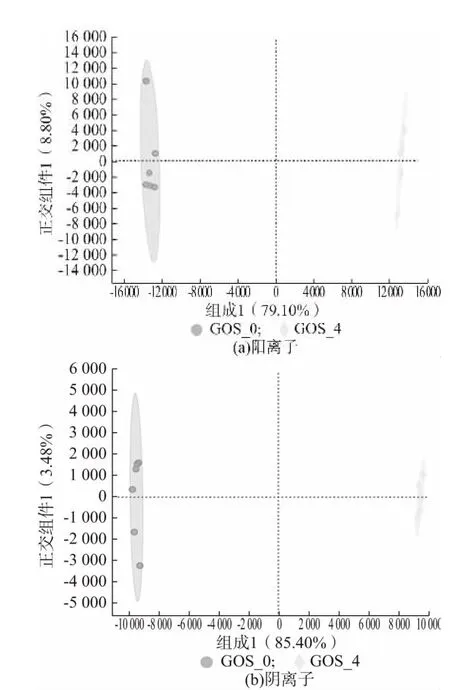
圖3 GOS_0 與GOS_4 代謝產物的OPLS-DA 分析
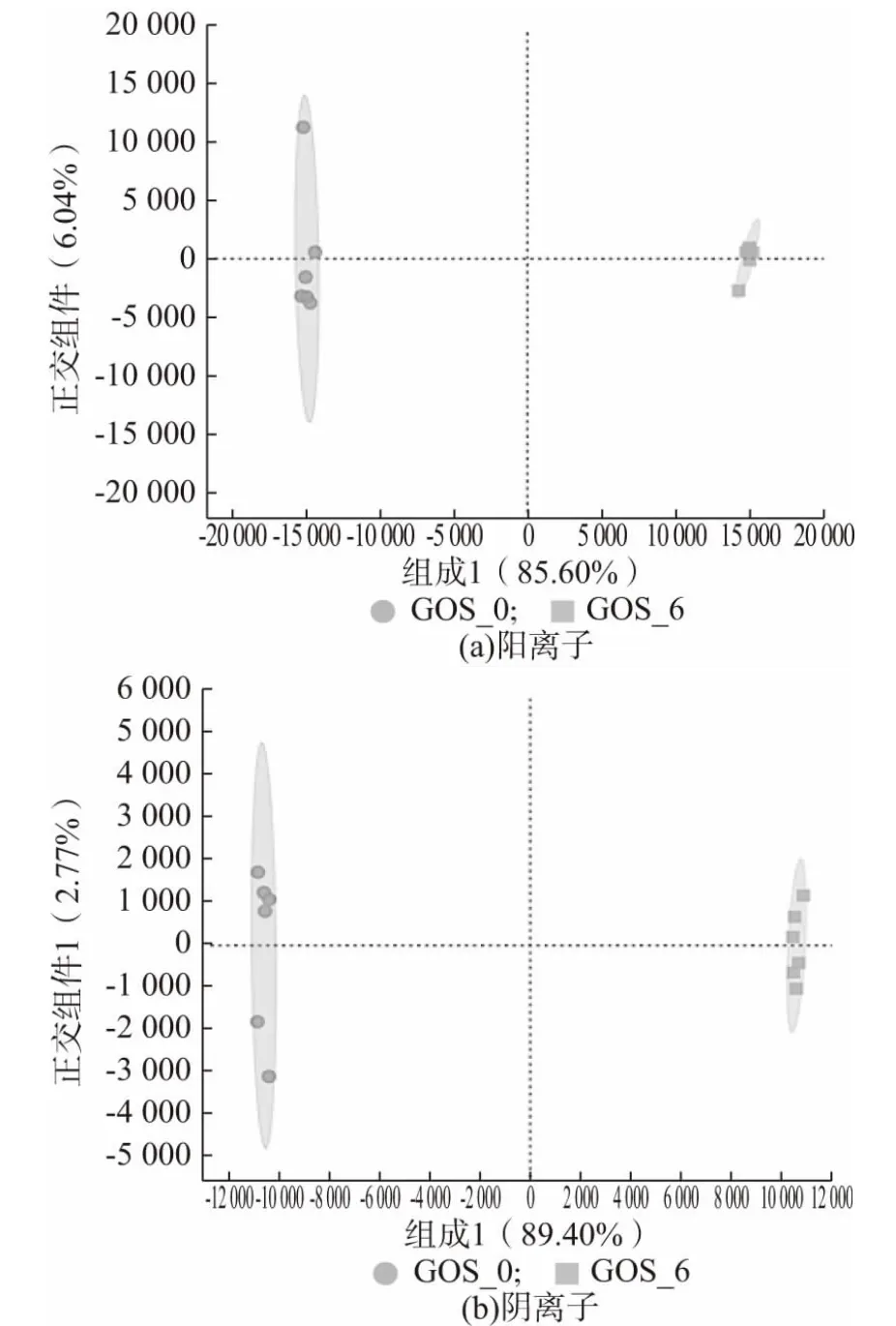
圖4 GOS_0 與GOS_6 代謝產物的OPLS-DA 分析
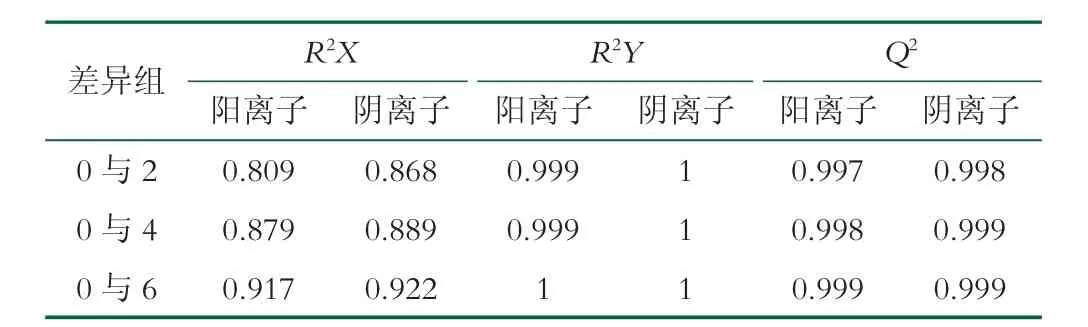
表2 OPLS-DA 統計結果
2.1.3 發酵乳的差異代謝物篩選
代謝組學是對生物樣本中存在的小分子的綜合研究,是一種更好地理解宿主-微生物相互作用的新方法,也是對樣本潛在益生性的一種探究[17]。為了明確不同GOS 添加量的益生性發酵乳之間的代謝物差異,以P<0.05、VIP值>1 為篩選標準,將GOS 添加量為2%、4%、6%的益生性發酵乳分別與未添加GOS(0%)的發酵乳進行兩兩分組比較,并分析各差異組之間代謝物數目的異同。圖5 反映了GOS_0 與GOS_2、GOS_0 與GOS_4、GOS_0 與GOS_6 差異代謝產物的數目。從圖5 中可以看到,GOS_2 與GOS_0有139 種差異代謝物,GOS_4 與GOS_0 有147 種差異代謝物,GOS_6 與GOS_0 有149 種差異代謝物。

圖5 差異代謝物數目
結合KEGG 數據庫對這些差異物質中的功能性代謝產物進行進一步篩選,結果如表3 所示,篩選出24種功能性代謝產物影響代謝過程。圖6 為KEGG 化合物的分類,由圖6 可知,篩選出來的功能性差異代謝產物主要包括氨基酸、有機酸、核苷酸及碳水化合物。其中,氨基酸是生命體的物質基礎,是生命體正常生長代謝至關重要的營養物質,也能增加機體抵抗力[18],若攝入其不足會導致低血糖[19]。氨基酸不僅可以縮短發酵時間,其作為風味化合物的前體物質,能夠進一步轉化成醛類、酯類和醇類等物質,從而影響發酵產品的風味[13]。有機酸除了調味以外,還具備殺菌和抑菌等作用,可以調節宿主免疫功能[19]。核苷酸幾乎參與了生物體內的所有反應過程,可以為生物體核酸合成提供材料,并且在能量代謝上具有重要功能,缺乏核苷酸會損害腸道、肝臟和免疫系統[20]。碳水化合物是生命體的重要供能物質,可以起到調節生命體日常活動的作用。其中異檸檬酸鹽、烏頭酸、肌醇、2-脫氧-D-核糖、β-羥基丁酸等含量顯著增加,1-磷酸葡萄糖、尿酸含量降低。
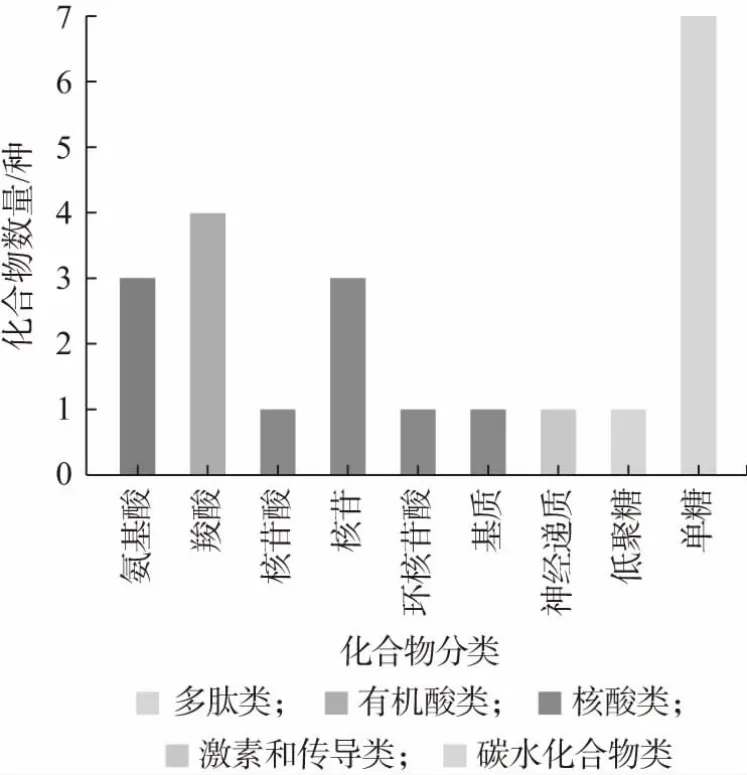
圖6 KEGG 化合物分類
2.1.4 代謝通路分析
GOS 植物乳桿菌發酵乳的差異代謝物的代謝途徑分析結果如圖7 所示。由圖7 可知,通路分析結果顯示共得到26 個途徑,以P<0.05 為標準對關鍵代謝途徑進行篩選,其中有4 條存在顯著性差異,分別為氨基糖和核苷酸糖代謝途徑、半乳糖代謝途徑、TCA 循環、嘧啶代謝途徑。胞嘧啶核苷、異檸檬酸鹽、烏頭酸、肌醇、β-羥基丁酸等顯著增加,1-磷酸葡萄糖、尿酸含量降低。胞嘧啶核苷參與嘧啶代謝途徑,是抗病毒與抗腫瘤類藥物的生產過程中的重要中間體[21]。代謝途徑與功能性差異代謝產物聯合分析如圖8 所示,異檸檬酸鹽與烏頭酸作為有機酸,參與三羧酸(TCA)循環,TCA 循環是生物體內營養物質氧化供能的共同途徑,異檸檬酸、烏頭酸含量增加,說明能夠提高生物體內營養物質的“逐步釋能”[22]。β-羥基丁酸在機體內含量較低,但其以信號分子、能量底物等多種物質形式廣泛參與到機體健康成長過程中,β-羥基丁酸經脫氫酶作用被氧化形成乙酰乙酸,乙酰乙酸經酮體氧化,而后通過硫解酶催化得到乙酰CoA[23],通過乙酰CoA 連接糖酵解途徑與TCA 循環,其作為具有生物活性的小分子物質,具有保護神經、心血管等組織器官的作用,對緩解骨質減少有保護功能。尿酸是人體嘌呤代謝的終產物,磷酸戊糖途徑屬于嘌呤代謝的一種[19],嘌呤代謝異常會引發多種疾病,尿酸主要由腎臟排泄,尿酸積聚可能會導致痛風。肌醇參與半乳糖代謝途徑,其常作為營養強化劑被應用于功能性食品中,可以保護肝臟,對骨骼肌代謝和骨骼生長具有積極作用,并且可以加快脂肪的消耗,有效預防脂肪堵塞心血管癥狀[24]。
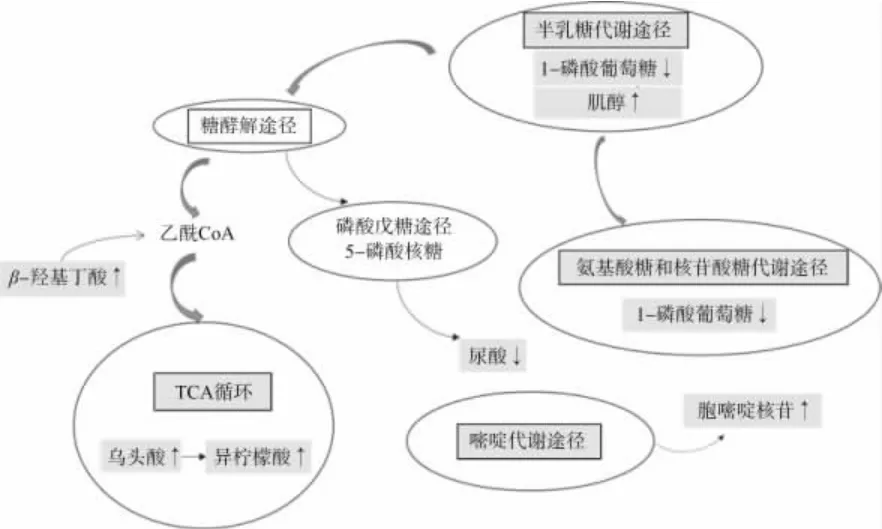
圖8 代謝途徑與功能性差異代謝產物關聯分析
因此,GOS 的添加能夠促進發酵乳次級代謝的合成與積累,使GOS 植物乳桿菌發酵乳具有保護心血管、增強骨骼密度、供給人體能量等潛在功效,是一種能潛在降低痛風風險的發酵乳產品。
2.2 GOS 植物乳桿菌發酵乳對腸道菌群的調節作用
2.2.1 物種α-多樣性分析
α-多樣性常用來指示某個特定的區域或者生態系統內的物種或種群的多樣性,其中ace 指數常用來反映群落的豐富度,ace 指數越高,群落的豐富度越大,如圖9(a)所示,ace 指數無顯著變化(P>0.05),說明群落中含有的物種總數無顯著變化;simpson 指數常用來反映群落物種多樣性,群落多樣性是對群落豐富度與均勻度的綜合分析,simpson 指數越大,物種多樣性越小,由圖9(b)可知,GOS 植物乳桿菌發酵乳降低了群落多樣性,可能是部分益生菌成為了優勢菌群,因此使得群落的均勻度降低。Arnold 等用含益生元的食物喂食動物2 周后,動物微生物組成的α-多樣性顯著降低,有益微生物的相對豐度增加,包括Bifidobacterium和Bacteroides等[25]。此外,有研究發現GOS 降低了整體菌群多樣性,增加了擬桿菌屬與乳酸桿菌屬的豐度。
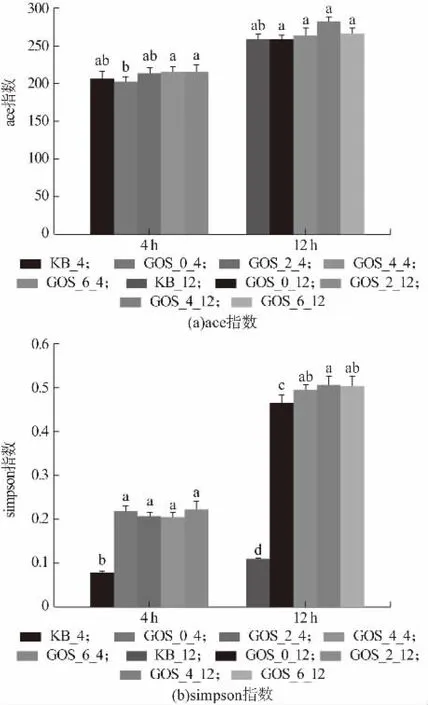
圖9 物種多樣性分析
2.2.2 GOS 益生性發酵乳對腸道菌群群落的組成分析
益生菌通過多種途徑影響人體健康,微生物群落組成失衡(失調)會導致冠心病、脂肪肝、類風濕性關節炎、炎癥性腸病等多種疾病[1]。圖10 反映的是門水平上腸道菌群的物種組成。由圖10 可知,腸道菌群主要由放線菌門(Actinobacteria)、厚壁菌門(Firmicutes)、擬桿菌門(Bacteroidetes)、變形菌門(Proteobacteria)組成,添加GOS 的益生性發酵乳和糞便共培養4、12 h 后,在門水平上,Actinobacteria 和Bacteroidetes 的相對豐度有所提高,而Firmicutes 的相對豐度降低;圖11 反映的是屬水平上腸道菌群的物種組成。由圖11 可知,在屬水平上,GOS 益生性發酵乳增加了群落中Bifidobacterium, Bacteroides,Koalas等有益菌的豐度,抑制了Shigella等有害菌的繁殖。在人體腸道環境中,Bifidobacterium屬于Actinobacteria 的主要菌屬,Bifidobacterium的增殖可代謝產生乳酸、乙酸、丁酸等這類酸性物質,這些酸性物質可降低機體腸道環境的pH,對有害菌的生長起到抑制作用[26]。Koalas可以產生短鏈脂肪酸,其屬于Firmicutes,Koalas相對豐度增加,而Firmicutes 整體豐度下降,說明GOS益生性發酵乳對Firmicutes 起到一定的調節作用[8]。因此,GOS 益生性發酵乳雖然降低了菌群多樣性,但是提升了有益菌的相對豐度,降低了有害菌的相對豐度,有益于調節人體健康。
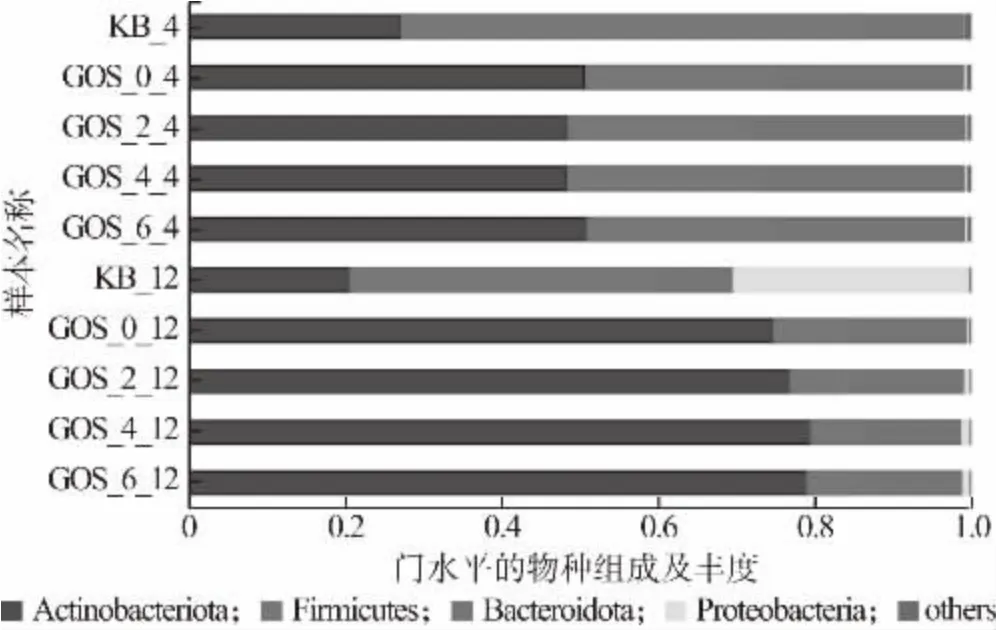
圖10 腸道菌群在門水平的物種組成及其豐度
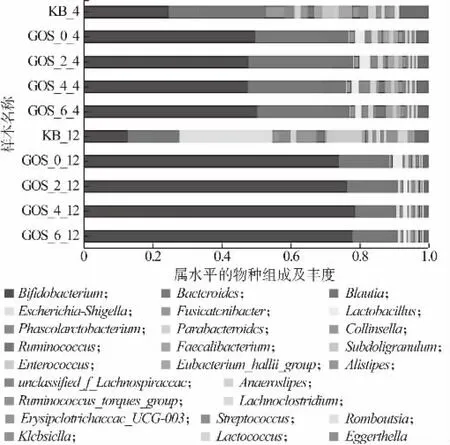
圖11 腸道菌群在屬水平的物種組成及其豐度
2.2.3 物種β-多樣性分析
采用β-多樣性分析探究不同樣本間微生物群落物種組成的相似度及差異性。PCoA 分析(Principal co-ordinates analysis),即主坐標分析,其與PCA 分析相類似。結果如圖12(a)所示,同種顏色的點代表來自同一分組的樣本,培養4 h 與12 h 后,添加GOS 的益生性發酵乳與空白對照組空間距離較遠,說明菌群組成相似性低,表明添加GOS 的益生性發酵乳對腸道菌群組成的影響較大。
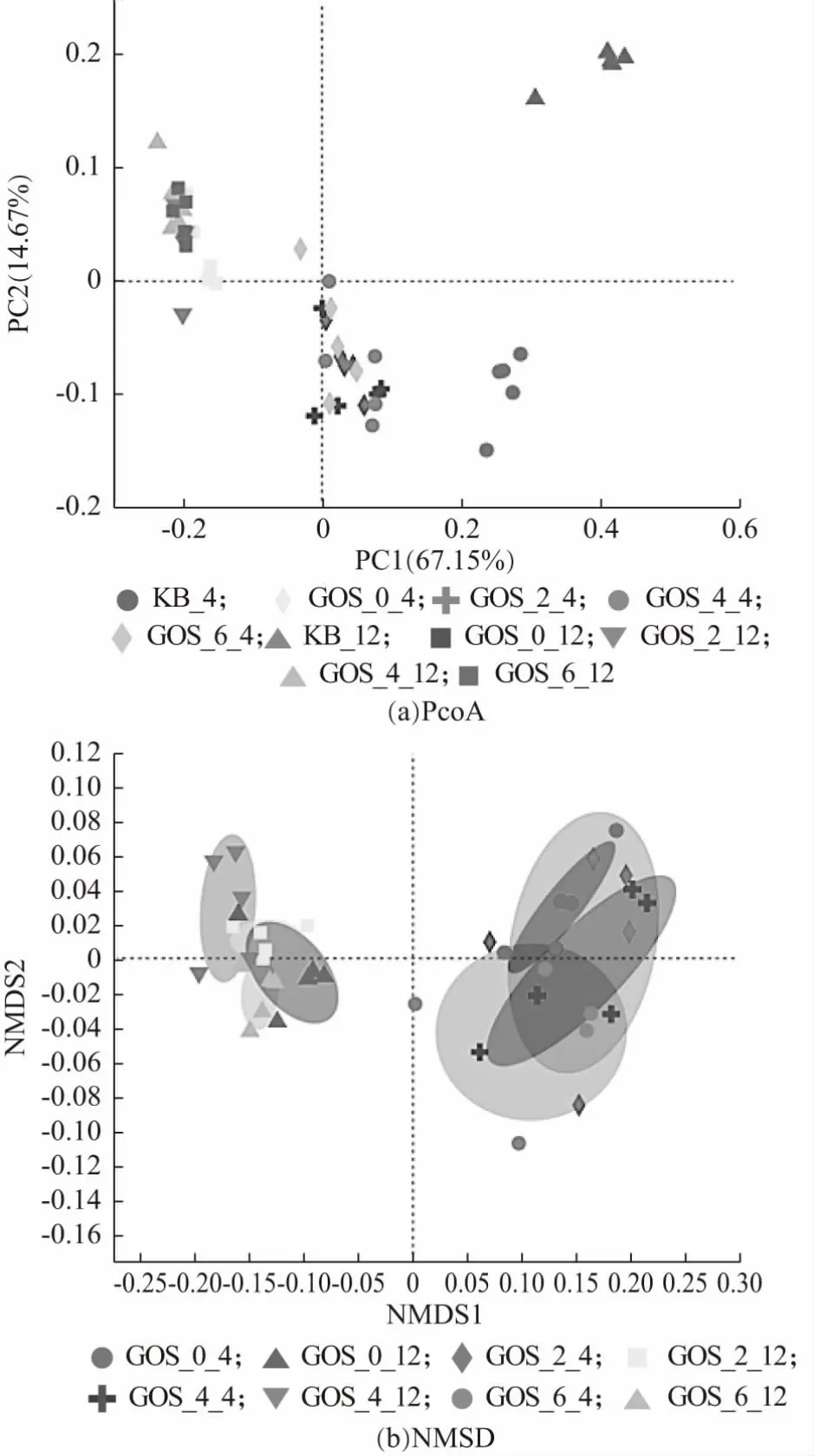
圖12 腸道菌群β-多樣性分析
非度量多維尺度分析(NMDS 分析)是可以在保留原始樣本的關系基礎之上,降維分析比較不同樣本組之間的差異。stress 值為0.042,低于0.05,表明NMDS結果具有很好的代表性,反映添加GOS 的益生性發酵乳所導致的群落結構差異。如圖12(b)所示,圖中每一個樣本都用一個點來表示,同組樣本的顏色一樣,兩點之間相距越緊密表明兩者的菌群結構組成越相近。由圖12(b)可見,培養4 h 后,在屬水平上,GOS 添加量為4%與6%的益生性發酵乳與未添加GOS 的發酵乳對腸道菌群的影響有較大差異,而GOS 添加量為2%的益生性發酵乳對腸道菌群的影響差異較小。培養12 h 后,在屬水平上,GOS 添加量為4%的發酵乳對腸道菌群的影響具有顯著差異。說明GOS 添加量為4%時,該GOS 益生性發酵乳對腸道菌群物種組成影響最大。
3 結 論
代謝組學研究表明,植物乳桿菌發酵乳中添加GOS 后,其中氨基酸、有機酸、核苷酸及碳水化合物等代謝物顯著增加(P<0.05),同時發現了4 條具有顯著性差異的代謝產物代謝途徑,分別是氨基糖和核苷酸糖代謝途徑、半乳糖代謝途徑、TCA 循環、嘧啶代謝途徑,正好同顯著增加的代謝產物相對應。說明GOS的添加可以加快植物乳桿菌代謝蛋白質、核酸和乳糖,其原理推測是因為GOS 的添加首先激活植物乳桿菌體內半乳糖代謝途徑,提高植物乳桿菌對GOS的利用,加快植物乳桿菌的生長,進一步催化體內的氨基酸和核苷酸糖代謝途徑、TCA 循環和嘧啶代謝途徑,促進植物乳桿菌對蛋白質、核酸和乳糖的代謝。人體腸道菌群在GOS 益生性發酵乳中培養后,增加了雙歧桿菌、擬桿菌屬、考拉菌屬等有益菌的豐度,抑制了大腸埃希氏菌等有害菌的繁殖。DA SILVA 等[27]在對低聚果糖(FOS)的體外糞菌發酵實驗中發現FOS 可以顯著促進發酵液中擬桿菌和雙歧桿菌的增殖,并促進了腸道細菌有益代謝產物短鏈脂肪酸的產生,ZHANG Y A 等[28]在對比果聚糖和FOS 對豬腸道菌群的影響中發現兩者對腸道微生物組成和代謝存在不同動態影響,果聚糖比FOS 具有更強的促進回腸中微生物氨基酸發酵和糞便中碳水化合物發酵的能力。由此說明腸道菌群的種類與氨基酸、碳水化合物、有機酸等代謝物含量息息相關,GOS 通過促進植物乳桿菌代謝生成大量的氨基酸、碳水化合物和有機酸等代謝物從而改善腸道菌群中有益菌和有害菌的比例。
猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
中學生數理化·七年級數學人教版(2019年10期)2019-11-25 07:33:58
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
電子制作(2018年18期)2018-11-14 01:48:24
中學生數理化·高一版(2018年9期)2018-10-09 06:46:50
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
湖南教育·C版(2018年3期)2018-06-05 16:54:36
山東工業技術(2016年15期)2016-12-01 05:31:22
鄭州大學學報(醫學版)(2015年2期)2015-02-27 14:50:46
