模型化研究兩細(xì)胞間基因、蛋白耦合振蕩中的噪聲效應(yīng)
2024-01-18 07:53:12李九智王書恒劉彼得趙新軍
原子與分子物理學(xué)報 2024年4期
關(guān)鍵詞:信號
蘇 瑞,李九智,李 循,王書恒,劉彼得,趙新軍
(1.伊犁師范大學(xué) 物理科學(xué)與技術(shù)學(xué)院 新疆凝聚態(tài)相變與微結(jié)構(gòu)實驗室,伊寧 835000;2.伊犁師范大學(xué) 微納電傳感器技術(shù)與仿生器械實驗室,伊寧 835000; 3 新疆維吾爾自治區(qū)人民醫(yī)院 泌尿外科,烏魯木齊 830000)
1 引 言
噪聲可以導(dǎo)致基因調(diào)控網(wǎng)絡(luò)中的細(xì)胞間變異[1],根據(jù)噪聲來源,可分為內(nèi)在和外在噪聲[2,3].基因表達(dá)包括兩個主要步驟:轉(zhuǎn)錄和翻譯,與許多其他生化機(jī)制一樣,由于基因產(chǎn)物拷貝數(shù)的隨機(jī)波動,發(fā)生蛋白質(zhì)合成的基因表達(dá)本質(zhì)上是隨機(jī)的[4],產(chǎn)生的蛋白質(zhì)量會因群體中的細(xì)胞而異,并且會隨著時間的推移在單個細(xì)胞中發(fā)生變化,這樣即使所有細(xì)胞都處于完全相同的狀態(tài),仍然會導(dǎo)致基因轉(zhuǎn)錄和翻譯的反應(yīng)事件在不同的時間、以不同的順序在不同的細(xì)胞中發(fā)生,這種隨機(jī)效應(yīng)是由基因序列及其編碼的蛋白質(zhì)的特性在局部設(shè)置的,被稱為“內(nèi)在”噪聲.此外,由于細(xì)胞中的其他分子種類,例如 RNA 聚合酶 (RNAP),它們本身就是基因產(chǎn)物,會隨著時間和細(xì)胞的不同而變化.這種變化會導(dǎo)致感興趣基因表達(dá)的額外的和相應(yīng)的波動,這種波動被稱為“外在”噪聲.因此,外在噪聲獨立于基因而產(chǎn)生,但會作用于基因[3-6].
體節(jié)的模式(PSM)是通過涉及胚胎尾端振蕩基因表達(dá)的機(jī)制來追蹤的,在一些脊椎動物(例如,斑馬魚)體節(jié)細(xì)胞中,兩個連鎖的振蕩基因 her1 和 her7 編碼抑制基因調(diào)控蛋白,其中的 Notch 信號是相鄰區(qū)域同步所必需的.Notch 是信號途徑中的一種跨膜蛋白受體,其信號途徑在多樣化的發(fā)育過程中起作用[7].與許多其他生化機(jī)制一樣,由于her1 和 her7 等基因產(chǎn)物拷貝數(shù)的隨機(jī)漲落,成為了 Notch 信號系統(tǒng)主要的噪聲來源,并會導(dǎo)致基因調(diào)控網(wǎng)絡(luò)中細(xì)胞間的變異可能性增加[8-10].Keskin 等人[11]發(fā)現(xiàn),外在噪聲支配時鐘基因 her1 和 her7 轉(zhuǎn)錄變異性,而基因內(nèi)在噪聲在低轉(zhuǎn)錄水平時升高,并與平均表達(dá)水平呈負(fù)相關(guān).雖然噪聲通常被認(rèn)為是一種必須過濾掉的擾動,但它也可以在系統(tǒng)中誘發(fā)新的有組織的行為,而這些行為在確定性條件下是不存在的,噪聲導(dǎo)致生物和物理系統(tǒng)中出現(xiàn)自發(fā)秩序(包括隨機(jī)共振、噪聲引起的相變和噪聲引起的雙穩(wěn)態(tài))[12,13].另外,生命系統(tǒng)本質(zhì)上是有噪聲的,并且經(jīng)過優(yōu)化以在存在隨機(jī)波動的情況下持續(xù)運行[14].Linden等人[15]的研究結(jié)果也表明,噪聲有助于持續(xù)的樹突非線性,它可以被認(rèn)為是樹突輸入集成策略的主要組成部分.Steuer 等人[16]證明了在細(xì)胞周期過程隨機(jī)漲落(噪聲)會導(dǎo)致動力學(xué)發(fā)生質(zhì)的變化,引起的信號通路系統(tǒng)振蕩的出現(xiàn).實驗結(jié)果進(jìn)一步表明[17-20],Notch 通路突變體中的總噪聲由于不同步振蕩和振幅降低而升高,噪聲是野生型胚胎轉(zhuǎn)錄變異的主要組成部分,噪聲可能會調(diào)節(jié) Notch 通路系統(tǒng)中調(diào)節(jié)因子的轉(zhuǎn)錄水平,破壞her1 和 her7 等基因的振蕩表達(dá),導(dǎo)致染色體基因座的表觀遺傳狀態(tài)的細(xì)胞間變異.這樣,非常有趣的問題出現(xiàn)了,那就是在 Notch 信號通路系統(tǒng)中,噪聲到底是優(yōu)化促進(jìn)信號傳導(dǎo)的穩(wěn)定性,還是擾亂穩(wěn)定的信號傳傳輸,內(nèi)在噪聲和外在噪聲各自是如何具體調(diào)控通路中信號傳導(dǎo),它們的作用是相同還是相反?既然生化反應(yīng)是隨機(jī)的,隨機(jī)的反應(yīng)伴隨著隨機(jī)的漲落即噪聲,隨機(jī)的反應(yīng)以及隨機(jī)的噪聲必然會使得生物系統(tǒng)的生化反應(yīng)無規(guī)律,不能產(chǎn)生節(jié)律性,然而生物系統(tǒng)往往能夠維持穩(wěn)健的節(jié)律性.這樣,隨機(jī)噪聲是如何將無規(guī)的隨機(jī)反應(yīng)變成有規(guī)律的節(jié)律性,必然有內(nèi)外噪聲的調(diào)控效應(yīng).那么,內(nèi)外噪聲是如何調(diào)控,使得生物系統(tǒng)維持穩(wěn)健的節(jié)律性?之前的研究[21-24]在噪聲影響基因調(diào)控動力學(xué)方面已經(jīng)取得了重要進(jìn)展,發(fā)現(xiàn)了一些重要的影響特性,但對于內(nèi)在噪聲和外在噪聲是如何通過各自調(diào)節(jié)機(jī)制調(diào)控通路同信號傳導(dǎo),進(jìn)而維持生物系統(tǒng)穩(wěn)健的周期節(jié)律性,還沒有給出明確的結(jié)果.最近的建模研究強(qiáng)調(diào)了分子噪聲的重要性,及其對產(chǎn)生單細(xì)胞節(jié)律性的貢獻(xiàn)[25-27].重要的是,細(xì)胞間耦合和分子噪聲的組合以前沒有被證明在沒有組件振蕩元素的情況下會產(chǎn)生振蕩.因此始終沒有確定細(xì)胞間 Notch 信號系統(tǒng)中內(nèi)在和外在噪聲信號對細(xì)胞分裂的調(diào)控作用,很難從基因表達(dá)調(diào)控中的隨機(jī)變異(噪聲)中得出的正確的結(jié)論.鑒于目前對開發(fā)基于細(xì)胞的療法的興趣,Notch 信號可能成為操縱細(xì)胞命運決定結(jié)果以在體外產(chǎn)生特定細(xì)胞類型的有吸引力的目標(biāo).在本文中,將建立動力學(xué)理論模型,研究兩細(xì)胞間 Notch 信號通路系統(tǒng)振蕩過程中基因、蛋白耦合振蕩中的內(nèi)在、外在噪聲效應(yīng),明確內(nèi)在噪聲和外在噪聲是如何通過各自調(diào)節(jié)機(jī)制調(diào)控通路中信號傳導(dǎo)機(jī)理,為設(shè)計阻止Notch 功能異常導(dǎo)致的多種疾病和癌癥的通路治療方案提供參考.
2 理論模型
實驗結(jié)果表明[20,21],在 Notch 信號系統(tǒng)中,時鐘基因 her1 和 her7 等的轉(zhuǎn)錄調(diào)節(jié)deltaC 蛋白(delta)合成,同時存在細(xì)胞間耦合和分子噪聲調(diào)節(jié)her1 和 her7 等,細(xì)胞間耦合可以誘導(dǎo)周期振蕩器群體的節(jié)律性,以及細(xì)胞間耦合可以同步并提高細(xì)胞內(nèi)、細(xì)胞間的振蕩.因此,細(xì)胞間 Notch 信號系統(tǒng)的物理模型如圖1 所示為:
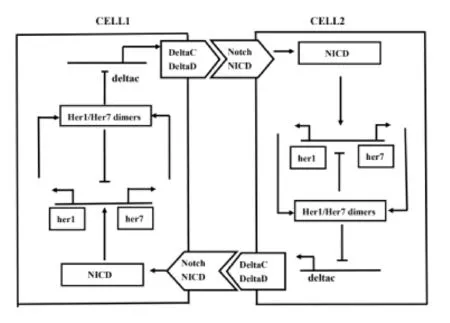
圖1 細(xì)胞間 Notch 信號通路的具體模型示意圖.
基于 Hill 動力學(xué)與 Michaelis-Menten 方程[28,29],兩細(xì)胞間 her1 和 her7、delta 蛋白及其相應(yīng)的 mRNA:mher1、mher7、mdelta 耦合過程中,Notch 信號通路系統(tǒng)各組分濃度隨時間演化的動力學(xué)方程組為
(1)
(2)
(3)
(4)
(5)
(6)
以上方程組中:
(7)
(8)
(9)
其中φdelta=[delta(t)]/[delta0],φher1=[her1(t)]/[her10],φher7=[her7(t)]/[her70],[delta0]=40 nM,[her10]=[her70]=1000 nM為臨界值.方程式 (1)-(6)中,ξ(t) 為高斯白噪聲,其性質(zhì)有:<ξ(t)>=0,<ξ(t)ξ(t′)>=δ(t-t′),Dγ(γ=h,d,mh,md)為噪聲幅值.通過兩細(xì)胞間 NICD 與 delta 耦合激活,兩細(xì)胞間形成了基因、蛋白耦合激活動力學(xué)特性,方程 (1)-(3)中由基因編碼的 her1 和 her7、delta 蛋白合成中的產(chǎn)生的噪聲為內(nèi)噪聲.方程 (4)-(6)中相應(yīng)的 mRAN 激活過程中,基因表達(dá)產(chǎn)生的隨機(jī)波動為外在噪聲.Ozbudak 與 Lewis[30]已經(jīng)通過擬合實驗數(shù)據(jù),確定了量化體系的模型中的部分參數(shù),我們也可以通過考察模型對參數(shù)取值的計算結(jié)果,與實驗結(jié)果對比的檢測參數(shù)設(shè)置的合理性.模型中的參數(shù)取值見表1.

Table 1 模型中的參數(shù)取值
3 結(jié)果與討論
兩細(xì)胞間基因、蛋白耦合振蕩具有周期振蕩特性,通過 Notch 信號通路,her1、her7、delta 等基因、蛋白調(diào)控細(xì)胞間供體、受體活性,完成一次次細(xì)胞間的信號傳導(dǎo),實現(xiàn)細(xì)胞的多次分裂增殖[21,30].
圖2呈現(xiàn)了Dh=Dd=Dmh=Dmd=0 時[her1]、[her7]、[deltaC]、[mher1]、[mher7]與[mdeltaC]隨時間演化的動力學(xué)關(guān)系.從圖2a-2f 可以看出,無噪聲影響時,[her1]、[her7]、[deltaC]、[mher1]、[mher7]與[mdelta]隨時間的演化均呈現(xiàn)了周期性振蕩特性,這意味著基因的轉(zhuǎn)錄在兩細(xì)胞中基因、蛋白幾乎同步激發(fā).細(xì)胞間基因、蛋白耦合振蕩的周期性表明,如果時鐘和波前機(jī)制要引導(dǎo)規(guī)則的體節(jié)分割,相鄰的細(xì)胞必須同步振蕩,這是通過在細(xì)胞間信號傳導(dǎo)過程中運行的 Notch 信號機(jī)制來實現(xiàn)同步[21,31].因此,delta 表達(dá)與 her1 和 her7 的表達(dá)同時受到 her1/her7 蛋白直接作用于 delta 啟動子的調(diào)節(jié),并且 delta 蛋白質(zhì)壽命很短,因為遞送到細(xì)胞表面的 delta 蛋白會被迅速內(nèi)吞,降解的 delta 振蕩將根據(jù)其相鄰細(xì)胞中的 delta 水平引起每個細(xì)胞中 Notch 激活水平的振蕩,激活 Notch 刺激 her1 和 her7 的表達(dá)[32].這樣,her1 和 her7 的轉(zhuǎn)錄也受兩個振蕩影響的產(chǎn)物控制:激活的 Notch 的正調(diào)節(jié)和 her1/her7 的抑制,可在任何轉(zhuǎn)錄控制系統(tǒng)中,her1/her7 在 PSM 中的振蕩稱為體節(jié)時鐘,PSM與確定的體節(jié)組織之間的運動界面稱為體節(jié)發(fā)生波前.因此,體節(jié)圖案依賴于時鐘和波前機(jī)制,一個完整的省略雛形由在一個完整的時鐘周期過程中從 PSM 中出現(xiàn)的一組細(xì)胞組成[33-35].
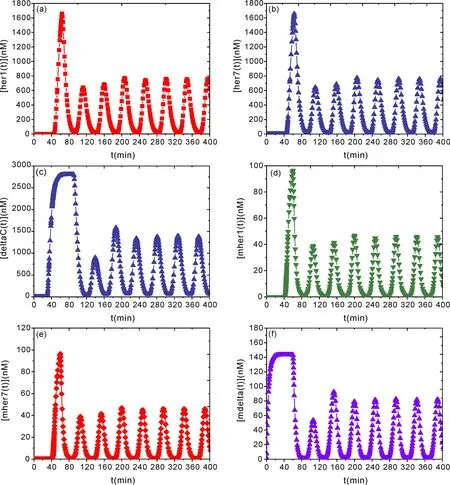
圖2 Dh=Dd=Dmh=Dmd=0 時[her1]、[her7]、[delta]、[mher1]、[mher7]與[mdelta]隨時間演化的動力學(xué)關(guān)系.
her1、her7 和 delta 等基因的轉(zhuǎn)錄調(diào)節(jié)合成等隨機(jī)漲落不可避免,基因表達(dá)是一個隨機(jī)的或“嘈雜的噪聲”過程,這種噪聲有兩種方式,轉(zhuǎn)錄和翻譯等生化過程的固有隨機(jī)性會產(chǎn)生“內(nèi)在”噪聲,以及其他細(xì)胞成分的數(shù)量或狀態(tài)的波動會間接導(dǎo)致特定基因表達(dá)的變化,從而代表“外在”噪聲.mRNA 分子中的莖環(huán)等結(jié)構(gòu)元件可通過與其非翻譯區(qū)結(jié)合而產(chǎn)生內(nèi)噪聲,mRNA 的非翻譯區(qū)通常包含的這些莖環(huán),可以可逆地改變構(gòu)型,使單個 mRNA 具有翻譯活性/非活性[3,4].
圖3 呈現(xiàn)了在 Dh=0.00005、Dd=Dmd=Dmh=0、τher1=τher7=2.8 min,τmher1=τmher7=6.5 min、τmdelta=16.5 min 時,τdelta=30.0 min 與τdelta=50.5 min條件下,[her1]、[her7]、[delta]、[mher1]、[mher7]與[mdelta]隨時間演化的動力學(xué)關(guān)系.從圖3可以看出,由于內(nèi)噪聲影響,使得兩細(xì)胞間基因、蛋白隨著時間演化周期振幅逐漸降低.由此表明了,Notch 信號通過內(nèi)噪聲信號迅速的干擾,將會出現(xiàn)周期節(jié)律動力學(xué)特性的改變,經(jīng)過長時間的干擾,好可能出現(xiàn)信號傳遞的“休克”.但這種周期節(jié)律的改變可以通過時滯τdelta調(diào)節(jié)恢復(fù),通過時滯調(diào)控作用,在低幅或“休克”開始后約250分鐘內(nèi)驅(qū)動靶基因的上調(diào),上調(diào)了各蛋白表達(dá)水平.這樣到產(chǎn)生 NICD 所需的時間延遲.在后期(3 到 5 個體節(jié))階段熱休克的胚胎批次中,出現(xiàn)前的延遲再次增長[36],延遲會隨著“休克”階段的不同而變化,導(dǎo)致 NICD 的誘導(dǎo)水平存在差異.因此,時滯性有利于細(xì)胞間 Notch 信號通路中各基因、蛋白,以及耦合的基因、蛋白表達(dá)水平在“休克”后再次提升,以及周期節(jié)律的恢復(fù).此外,Notch 信號通路系統(tǒng)還可以通過轉(zhuǎn)錄因子調(diào)節(jié) RNA聚合酶的結(jié)合率來影響基因表達(dá)[37].
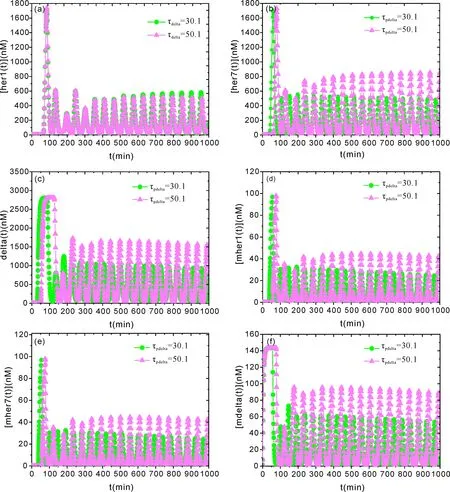
圖3 不同 τdelta 條件下,[her1]、[her7]、[delta]、[mher1]、[mher7]與[mdelta]隨時間演化的動力學(xué)關(guān)系.
除了時滯性的調(diào)控作用,細(xì)胞中的其他分子種類,例如 RNA 聚合酶和上游轉(zhuǎn)錄調(diào)節(jié)因子,是基因產(chǎn)物,因此也會隨著時間和細(xì)胞的不同而變化,這些變化導(dǎo)致時鐘基因表達(dá)的相關(guān)波動,并被確定為外在噪聲,因此,噪聲也會產(chǎn)生相應(yīng)的調(diào)節(jié)作用.
圖4 呈現(xiàn)了在 Dh=0.00004、Dmh=0.000001、Dmd=0.00001、τher1=τher7=2.8、τdelta=55.1、τmher1=τmher7=6.4、τmdelta=16.1時,Dd=0 與Dd=0.00005 條件下,[her1]、[her7]、[delta]、[mher1]、[mher7]與[mdelta]隨時間演化的動力學(xué)關(guān)系.從圖4 可以看出,隨著時間演化,內(nèi)外噪聲使得兩細(xì)胞間基因、蛋白耦合振福減弱,由此恢復(fù)正常的周期節(jié)律動力學(xué).由此表明了,當(dāng) NICD 適度表達(dá)的中等水平出現(xiàn)的隨機(jī)漲落,使得兩細(xì)胞間基因、蛋白耦合的同步性得以恢復(fù),伴隨著 her1 和 her7 表達(dá)峰值水平的適當(dāng)降低,依次誘導(dǎo)了下游基因蛋白的表達(dá)降低.其次,當(dāng) NICD 非常強(qiáng)烈地過度表達(dá)時,振蕩變得逐漸增強(qiáng),系統(tǒng)趨向于 her1 和 her7 的過高表達(dá)的狀態(tài).這些基因的適度表達(dá)值的早期消失可能導(dǎo)致體節(jié)邊界缺陷的相對早期發(fā)生.當(dāng) Notch 信號中斷時,噪聲會促使基因、蛋白恢復(fù)正常的周期節(jié)律振蕩,會對轉(zhuǎn)錄的總變異性抑制產(chǎn)生最大貢獻(xiàn)[21,38].并且,在任何轉(zhuǎn)錄控制系統(tǒng)中,基因表達(dá)調(diào)控中的隨機(jī)變異或噪聲都會有助于產(chǎn)生波動[2],在這里,我們發(fā)現(xiàn)適度的噪聲能夠恢復(fù) Notch 信號通路中的周期節(jié)律性.在 Notch 通路信號傳導(dǎo)過程中,內(nèi)外噪聲隨機(jī)性伴隨著信號傳導(dǎo)會同時作用.Keskin 等人[21]的結(jié)果表明,Notch 信號通路中,基因表達(dá)的噪聲控制有利于持續(xù)振蕩,為了進(jìn)一步明確內(nèi)外噪聲的調(diào)控效應(yīng),可以考察體系當(dāng)中內(nèi)外噪聲同時作用時,兩細(xì)胞間基因、蛋白耦合周期振蕩的動力學(xué)特性.
首先我們考察內(nèi)外噪聲對基因表達(dá)的自抑制振蕩的影響,在內(nèi)外噪聲同時作用時,假定噪聲在影響自抑制基因表達(dá)過程中產(chǎn)生相反效應(yīng),基于Ozbudak 與 Lewis[30]的研究,自抑制動力學(xué)方程可表為:
(10)
圖5 呈現(xiàn)了在k=4.50 時b=1.0、Di=0.0045、De=0.00098 與b=1.5、Di=0.0045、De=0.0025 條件下,[her1]與[deltaC]隨時間演化的動力學(xué)關(guān)系.圖5a 呈現(xiàn)了基因表達(dá)的直接自抑制可以產(chǎn)生振蕩,其中 Her1 或 Her7 蛋白直接與其自身基因的調(diào)控 DNA 結(jié)合以抑制轉(zhuǎn)錄實現(xiàn)周期振蕩.在內(nèi)、外噪聲共同作用條件下,圖5b 呈現(xiàn)了與無噪聲環(huán)境中幾乎完全一致的周期振蕩動力學(xué)特性.由此表明了內(nèi)、外噪聲共同作用可使得基因表達(dá)自抑制呈現(xiàn)出持續(xù)振蕩模式.

圖5 [her1]與[deltaC]隨時間演化的動力學(xué)關(guān)系.
對于給定的自調(diào)節(jié)基因,合成相應(yīng)的蛋白,在內(nèi)外噪聲同時作用時,假定噪聲在影響蛋白合成、基因激活表達(dá)過程中也產(chǎn)生相反效應(yīng),基于 Ozbudak 與 Lewis[30]的研究基礎(chǔ),在噪聲影響條件下,基因合成相應(yīng)的蛋白過程中動力學(xué)方程可表為:
(11)
(12)
圖6 呈現(xiàn)了在不同條件下,[her1]與[her7]隨時間演化的動力學(xué)關(guān)系.圖6 呈現(xiàn)了在內(nèi)、外噪聲共同作用條件下,基因、蛋白也呈現(xiàn)了與無噪聲環(huán)境中幾乎完全一致的周期振蕩動力學(xué)特性.由此表明,內(nèi)、外噪聲共同作用可使得基因合成相應(yīng)的蛋白過程中,系統(tǒng)恢復(fù)基因表達(dá)自抑制呈現(xiàn)出持續(xù)振蕩模式.內(nèi)外噪聲共同調(diào)節(jié)使得體系完全恢復(fù)到無噪聲條件下的動力學(xué)狀態(tài),由于噪聲的隨機(jī)性,內(nèi)外噪聲很可能會產(chǎn)生相反的效應(yīng).生物系統(tǒng)通過時間延遲,增大了反應(yīng)成分的量積累,從而增加了隨機(jī)的內(nèi)噪聲,使得內(nèi)在噪聲根據(jù)外在噪聲自激活,在一定程度上抵消了外在噪聲的影響,使得 Notch 信號通路系統(tǒng)能在內(nèi)、外噪聲的作用下優(yōu)化運行[2,16].

圖6 τm=2.8 min時[her1]與[deltaC]隨時間演化的動力學(xué)關(guān)系.
圖6 結(jié)果進(jìn)一步證明了,在內(nèi)、外噪聲同時作用下,內(nèi)外噪聲的擾動效應(yīng)相互消除,使得 Notch 信號通路能夠在內(nèi)、外噪聲環(huán)境下依然能夠較好地保持周期節(jié)律的動力學(xué)特性.復(fù)雜系統(tǒng)調(diào)控特性需要內(nèi)外噪聲以及時間延遲共同調(diào)節(jié)恢復(fù)原有的周期節(jié)律性振蕩特性.這是由于,兩細(xì)胞間基因、蛋白耦合周期振蕩動力學(xué)涉及多組分、多步驟特性,這種多組分、多步驟必然伴隨著時間延遲調(diào)控,隨著時間延遲的增加,合成反應(yīng)時間增加,體系不可避免地出現(xiàn)隨機(jī)漲落,即內(nèi)、外噪聲,大量隨機(jī)的內(nèi)、外噪聲在長時間的統(tǒng)計效應(yīng)相互抵消,使得體系在內(nèi)外噪聲同時作用時,在一定程度上回復(fù)原有的周期節(jié)律性振蕩特性,保證了兩細(xì)胞間基因、蛋白最終合成的周期節(jié)律性.因此,生物系統(tǒng)調(diào)節(jié)體系的噪聲是通過時間延遲調(diào)控,使得體系出現(xiàn)更多的隨機(jī)性,大量的隨機(jī)內(nèi)、外噪聲互相影響,產(chǎn)生的效應(yīng)可以互相抵消,這樣,基因表達(dá)的噪聲反而有利于控制持續(xù)振蕩[16,39].基因通常在細(xì)胞發(fā)育過程中的表達(dá)高度可變,這種高度可變性受到時間延遲控制,進(jìn)而調(diào)節(jié)內(nèi)、外噪聲,使其在存在隨機(jī)波動的情況下產(chǎn)生相反效應(yīng),互相抵消以保證優(yōu)化系統(tǒng)正常的動力學(xué)過程.
4 結(jié) 論
本文建立動力學(xué)理論模型,研究兩細(xì)胞間基因、蛋白耦合振蕩中的噪聲效應(yīng).研究發(fā)現(xiàn),兩細(xì)胞間基因、蛋白耦合振蕩具有周期振蕩特性,表明了相鄰的 PSM 細(xì)胞同步振蕩特性.Notch 信號通過內(nèi)、外噪聲信號迅速的干擾,將會出現(xiàn)周期節(jié)律動力學(xué)特性的改變,這種周期節(jié)律的改變可以通過時滯調(diào)節(jié)恢復(fù);在另一方面,內(nèi)外噪聲也具有恢復(fù)兩細(xì)胞間基因、蛋白耦合振蕩正常的周期節(jié)律動力學(xué)特性,適度的噪聲能夠恢復(fù) Notch 信號通路中的周期節(jié)律性.通過考察內(nèi)外噪聲對基因表達(dá)的自抑制振蕩的影響,我們還發(fā)現(xiàn),內(nèi)、外噪聲共同作用可使得基因表達(dá)自抑制呈現(xiàn)出持續(xù)振蕩模式.通過分析對于給定的自調(diào)節(jié)基因、合成相應(yīng)的蛋白過程中內(nèi)外噪聲同時作用,我們進(jìn)一步發(fā)現(xiàn),內(nèi)、外噪聲共同作用可使得基因合成相應(yīng)的蛋白過程中,系統(tǒng)恢復(fù)基因表達(dá)自抑制呈現(xiàn)出持續(xù)振蕩模式,進(jìn)一步明確了在 Notch 通路信號傳導(dǎo)過程中,內(nèi)外噪聲隨機(jī)性伴隨著信號傳導(dǎo)會有利于持續(xù)振蕩.
生物系統(tǒng)的噪聲使得體系出現(xiàn)更多的隨機(jī)性,大量的隨機(jī)內(nèi)、外噪聲互相影響,產(chǎn)生的效應(yīng)可以互相抵消,這樣,基因表達(dá)的噪聲有時反而有利于控制持續(xù)振蕩[2,16,39].發(fā)生蛋白質(zhì)合成的基因表達(dá)本質(zhì)上是隨機(jī)的.蛋白質(zhì)從生化反應(yīng)的角度來看,基因表達(dá)包括兩個主要步驟:轉(zhuǎn)錄和翻譯.雖然 RNA 聚合酶在前者中產(chǎn)生 mRNA 分子,但在后者中通過核糖體進(jìn)行蛋白質(zhì)合成,每個反應(yīng)對應(yīng)于相關(guān)物種的產(chǎn)生和衰變.具體而言,由于尋找潛在問題的解決方案所涉及的微妙之處,根據(jù)概率分布量化物種數(shù)量已成為一項有趣且具有挑戰(zhàn)性的工作[40].本文理論結(jié)果揭示了時滯、內(nèi)外與噪聲對 Notch 信號通路動力學(xué)的一種調(diào)控機(jī)制,確定了內(nèi)外噪聲各自的調(diào)控效應(yīng),澄清了內(nèi)外噪聲共同作用調(diào)控體系持續(xù)周期振蕩的物理機(jī)制,理論結(jié)果符合實驗[21,39],可為設(shè)計阻止 Notch 體系基因、蛋白變異導(dǎo)致的多種疾病和癌癥的通路治療方案提供理論依據(jù).
猜你喜歡
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
媽媽寶寶(2019年10期)2019-10-26 02:45:34
中國生殖健康(2019年3期)2019-02-01 06:12:26
鐵道通信信號(2018年11期)2019-01-19 01:15:08
電子制作(2018年11期)2018-08-04 03:25:42
鐵道通信信號(2018年2期)2018-04-18 12:18:10
鐵道通信信號(2016年11期)2016-06-01 12:11:32
鑿巖機(jī)械氣動工具(2016年3期)2016-03-01 04:00:25
中國病理生理雜志(2015年8期)2015-12-21 12:38:06
